转Cry1Ac+Cry2Ab棉对棉铃虫的控制作用及对天敌捕食烟粉虱功能反应的影响
2015-10-13雒珺瑜张帅吕丽敏王春义朱香镇崔金杰
雒珺瑜,张帅,吕丽敏,王春义,朱香镇,崔金杰
中国农业科学院棉花研究所,棉花生物学国家重点实验室,安阳 455000
转Cry1Ac+Cry2Ab棉对棉铃虫的控制作用及对天敌捕食烟粉虱功能反应的影响
雒珺瑜,张帅,吕丽敏,王春义,朱香镇,崔金杰
中国农业科学院棉花研究所,棉花生物学国家重点实验室,安阳 455000
文章以转Cry1Ac基因棉(中棉所41)和常规棉(中棉所49)为对照,研究了转Cry1Ac+Cry2Ab基因棉(639020)在棉花生长的关键时期——蕾期(二代棉铃虫发生期)、花期(三代棉铃虫发生期)和花铃期(四代棉铃虫发生期)对棉铃虫的控制作用,同时研究了639020棉田主要捕食性天敌(中华草蛉幼虫、龟纹瓢虫、小花蝽和草间小黑蛛)对烟粉虱的捕食功能,明确了639020棉花在生长的关键时期对棉铃虫的控制效果及对棉田主要捕食性天敌捕食功能反应的影响。结果表明,639020棉花对二代和三代棉铃虫具有良好的控制作用,抗虫性分别比中棉所41提高了52.85%和16.22%,其中前者差异达显著水平,后者差异不显著。在棉花蕾期、花期和花铃期,639020棉田棉铃虫落卵量都比中棉所41棉田和中棉所49棉田低(除二代棉铃虫发生期);棉铃虫幼虫数量都极显著低于常规棉,且都低于防治指标,但与中棉所41棉田无显著差异。639020棉田中华草蛉、龟纹瓢虫、小花蝽和草间小黑蛛对烟粉虱的捕食功能与中棉所41棉田和常规棉田相比无显著变化。研究结果以期为新型转基因棉花环境安全性研究及其外源基因的抗虫遗传效应和生产应用前景进行安全性评价。
转Cry1Ac+Cry2Ab棉;棉铃虫;捕食功能
全世界已有10多个主产棉国推广种植Bt棉,总面积为1500多万公顷[1]。转Bt基因棉随着种植面积的扩大和种植时间的延长,靶标害虫棉铃虫(Helicoverpa armigera)对Bt毒素的抗性虽然没有质的飞越,但其耐受性呈现逐年上升的趋势[2,3],持续大面积种植转Bt基因棉的潜在抗性风险将成为影响其使用寿命的限制性因子[4~6]。为避免将来单一种植Bt基因棉对棉铃虫抗性问题带来毁灭性的灾难,近年中国农业科学院棉花研究所成功将Cry2Ab基因转入我国主栽棉花品种,获得了转复合基因Cry1Ac+ Cry2Ab基因棉花新材料。复合基因棉花新材料的出现和种植,可以从抗性位点结合方面延缓棉铃虫抗性的产生。目前对转Cry1Ac+Cry2Ab基因棉的研究开展了其对棉蚜(Aphis gossypii)[7]、节肢动物群落[8]及棉田天敌生长发育[9]的影响,但对主要的目标性状未进行研究,很难对其目标基因遗传效应、生产应用价值和目标性状的效果有一个很好的定论。
烟粉虱(Bemisia tabaci)是一种世界性的害虫,在全世界几十个国家和地区均有分布。烟粉虱具有复杂和广泛的寄主植物,给世界各国的农业生产造成严重威胁和巨大的经济损失。近年来,我国大棚蔬菜及设施园艺作物迅猛发展,冬天大棚和设施农业中有适宜烟粉虱生存和繁殖的气候条件,为烟粉虱的常年转主危害奠定了条件。以前,棉田在防治棉铃虫时可以兼治的烟粉虱,现在由于种植了转基因抗虫棉花,用药量减少或不进行防治,使得棉田烟粉虱得不到兼治,从而取代棉铃虫地位上升为主要害虫[10~18]。目前对烟粉虱的防治主要采用化学防治,由于连续长期用药,使烟粉虱对大部分化学农药产生了很强的抗性和交互抗性[19],给烟粉虱的防治带来了极大的困难,因此开展烟粉虱生物防治,对减少化学农药的使用、延缓抗性和保护生态环境,具有重要的实践价值。
本文为进一步明确新型转Cry1Ac+Cry2Ab基因棉花对抗靶标害虫抗性的遗传效应和在育种方面的应用前景,系统测定了新型转基因棉花对靶标害虫的抗虫效果及其对主要天敌的影响。本研究拟结合室内生物测定和田间小区调查两种方法,明确转Cry1Ac+Cry2Ab棉花新材料对靶标害虫的控制作用和田间种群动态的影响,研究了转Cry1Ac+Cry2Ab棉田的主要捕食性天敌对烟粉虱的捕食功能,为转新型基因棉花的环境安全评价积累科学数据,为我国转基因棉花的环境安全管理提供科学依据,同时为转基因抗虫棉田主要害虫烟粉虱的综合防治技术提供理论指导。
1 材料和方法
1.1 材料
1.1.1 棉花
转Cry1Ac+Cry2Ab基因棉639020由中国农业科学院棉花研究所生物技术研究室提供;中棉所41为转Cry1Ac基因棉,中棉49为常规非转基因棉花,均由中国农业科学院棉花研究所遗传育种研究室提供。
1.1.2 试虫
烟粉虱采自非转基因棉田;棉铃虫于2013年9月采自田间,当年在室内人工饲养的第2代棉铃虫幼虫,由中国农业科学院棉花研究所养虫室提供;中华草蛉幼虫、龟纹瓢虫、小花蝽和草间小黑蛛成虫分别采自639020棉田、中棉所41棉田和中棉49棉田。
1.2 方法
1.2.1 棉花种植
试验于2014年5月上旬至9月下旬期间,将棉花种植于中国农业科学院棉花研究所试验农场,每材料种植3小区,小区面积122 m2,小区间随机排列,棉花种植株距为0.27 m,行距为0.80 m。棉花全生育期不进行化学防治,其他农事操作按常规棉田操作和管理。
1.2.2 田间种群调查方法
试验采用对角线五点取样方法[20],分别在二代、三代和四代棉铃虫发生高峰期,调查不同棉田棉铃虫落卵量和棉铃虫幼虫数量,每个棉铃虫发生世代每隔1 d调查1次,共计调查3次;每次每小区调查5个样点,每个样点连续调查两行20株棉花,共100株;棉田棉铃虫落卵量和棉铃虫幼虫数量分别以“百株落卵量”或“百株虫量”表示。
1.2.3 棉铃虫室内生物测定
在棉花生长的蕾期(二代棉铃虫发生期)、花期(三代棉铃虫发生期)分别采集639020、中棉所41和中棉49顶部第二片展开叶,每20片叶为1次重复,每个品种3次重复。将采集的叶片装入生测盒(直径9 cm、高15 cm),每个生测盒装入1片棉叶,每片棉叶上接入5头棉铃虫孵化2 d的幼虫,盖上盖子,放入光照培养箱中(温度27±0.5℃,相对湿度70%~ 80%,光照周期L∶D=14 h∶10 h),5 d后调查棉铃虫幼虫死亡数和活虫数,以常规棉为对照,计算转基因抗虫棉花对棉铃虫抗性的幼虫校正死亡率。计算公式: ()
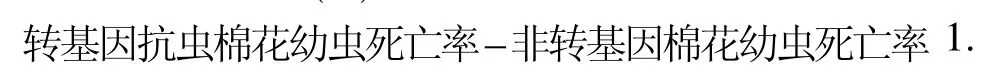
2.4 天敌捕食功能反应
天敌捕食功能反应分别以采自639020棉田、中棉所41棉田和中棉49棉田的中华草蛉幼虫、龟纹瓢虫、小花蝽和草间小黑蛛成虫为天敌,以采自非转基因棉田的烟粉虱为猎物,并在室内统一饥饿12 h后进行试验。用不同密度(每管 15、30、45、60、75头)的猎物和1头天敌放置在直径为9 cm、高15 cm的大玻璃试管内,然后置于光照培养箱中。每种天敌每个猎物密度6个试管为一次重复,试验均重复3次。24 h后调查剩余烟粉虱数量,建立捕食功能反应模型——HollingⅡ型反应模型,根据模型计算天敌日最大捕食量。功能反应圆盘方程为:
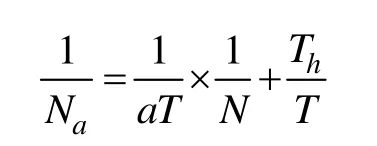
式中:Na为被捕食的猎物数量;N为猎物密度;a为瞬时攻击率;T为实验持续时间(1 d);Th为处置1头猎物时间。
1.3 统计分析
运用SPSS 13.0软件对所有数据进行统计分析,采用单因素方差分析(One-way analysis,ANOVA)和Duncan's差异显著性分析,检验同一时期不同小区内棉田棉铃虫落卵量和幼虫数量的差异。
2 结果与分析
2.1 转Cry1Ac+Cry2Ab棉在二代棉铃虫和三代棉
铃虫发生期对棉铃虫室内控制效果
转Cry1Ac+Cry2Ab棉639020和转Cry1Ac基因中棉所41棉叶片在二代棉铃虫和三代棉铃虫发生期对棉铃虫室内控制效果见表1。在二代棉铃虫和三代棉铃虫发生期,639020和中棉所41棉叶片室内生物测定棉铃虫幼虫校正死亡率分别为81.39%、53.25%和57.97%、49.88%;两个棉铃虫发生时期639020棉对棉铃虫的抗虫性比中棉所41分别高52.85%和16.22%,前者差异达极显著水平,后者差异不显著;两种转基因棉花对棉铃虫幼虫的控制作用均随着棉花的生长呈现下降的趋势,但639020棉由于前期效果较好,后期显示出下降比例较大。
2.2 转Cry1Ac+Cry2Ab棉在二代、三代和四代发生
期对田间棉铃虫的影响
2.2.1 棉田落卵量
在棉花生长的关键时期——蕾期(二代棉铃虫发生期)、花期(三代棉铃虫发生期)和花铃期(四代棉铃虫发生期)3种棉田百株棉铃虫落卵量如图1所示。由图1可见,在二代棉铃虫发生期,转Cry1Ac+ Cry2Ab棉639020、转Cry1Ac基因棉中棉所41和常规棉中棉所49棉田百株棉铃虫落卵量平均分别为297.4粒、331.6粒和206.6粒,639020棉田比中棉所41棉田少10.3%,比中棉所49棉田多44.0%,差异均不显著;在三代棉铃虫发生期,639020、中棉所41和中棉所49棉田百株棉铃虫落卵量平均分别为47.1粒、107.6粒和73.3粒,639020棉田比中棉所41和中棉所49棉田分别少56.2%和35.7%,前者差异显著,后者差异不显著;在四代棉铃虫发生期,639020、中棉所41和中棉所49棉田百株棉铃虫落卵量平均分别为22.7粒、32.2粒和23.7粒,639020棉田比中棉所41和中棉所49棉田分别少29.5%和4.2%,差异均不显著。

表1 两种转基因抗虫棉对棉铃虫校正死亡率(%)
2.2.2 棉田棉铃虫幼虫数量
在棉花生长的关键时期——蕾期(二代棉铃虫发生期)、花期(三代棉铃虫发生期)和花铃期(四代棉铃虫发生期)3种棉田百株棉铃虫幼虫数量如图2所示。由图2可见,在二代棉铃虫发生期,转Cry1Ac+ Cry2Ab棉639020、转Cry1Ac基因棉中棉所41和常规棉中棉所49棉田百株棉铃虫幼虫数量平均分别为7.1头、7.1头和59.8头,639020棉田和中棉所41棉田棉铃虫幼虫数量均比常规棉田少88.1%,差异达极显著水平;在三代棉铃虫发生期,639020棉、中棉所41和中棉所49棉田百株棉铃虫幼虫数量平均分别为4.9头、4.4头和36.3头,639020和中棉所41棉田百株棉铃虫幼虫数量分别比常规棉田少86.5%和87.9%,差异均达极显著水平;在四代棉铃虫发生期,639020棉、中棉所41和中棉所49棉田百株棉铃虫幼虫数量平均分别为0.9头、1.4头和15.1头,639020和中棉所41棉田百株棉铃虫幼虫数量分别比常规棉田少94.0%和90.7%,差异均达极显著水平;3个时期转基因棉田棉铃虫幼虫数量均在防治指标以下,可见田间控制效果良好。
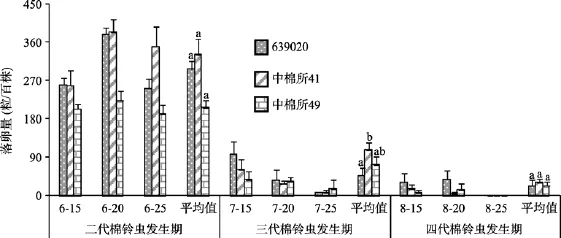
图1 棉铃虫发生期3种棉田棉铃虫落卵量
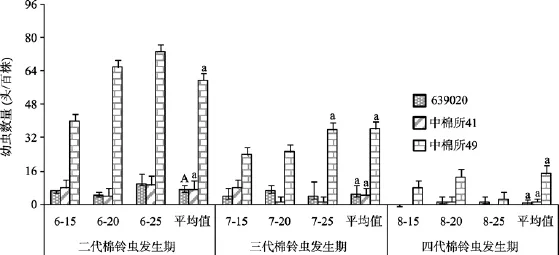
图2 棉铃虫发生期3种棉田棉铃虫幼虫数量
2.3 3种棉田主要捕食性天敌对烟粉虱捕食功能反应
2.3.1 3种棉田主要捕食性天敌对烟粉虱捕食功能反应
图3结果表明,在相同的环境条件下,棉田中华草蛉幼虫、龟纹瓢虫、小花蝽和草间小黑蛛成虫对烟粉虱成虫的捕食量均随烟粉虱密度的增加而上升,当烟粉虱的密度增加到75头/管后,4种天敌捕食量增加的速度变慢,呈现出负增长的趋势和曲线;在相同猎物密度条件下,3种棉田中每种天敌的捕食量差异不显著。
2.3.2 主要捕食参数
4种天敌对烟粉虱的捕食量与时间绘成散点图结果均符合HollingⅡ型反应如(表2)。由表2可见,功能反应方程相关系数均大于0.9544,表明在室温条件下中华草蛉幼虫、龟纹瓢虫、小花蝽和草间小黑蛛成虫的捕食量与烟粉虱的密度呈显著相关关系。由表2可见,中华草蛉幼虫对烟粉虱成虫具有较强的捕食作用。4种天敌中龟纹瓢虫对烟粉虱的捕食作用最强,日最大捕食量均在240头以上,其次是中华草蛉幼虫和小花蝽成虫,均在100头以上,而草间小黑蛛最低,平均为70头左右。
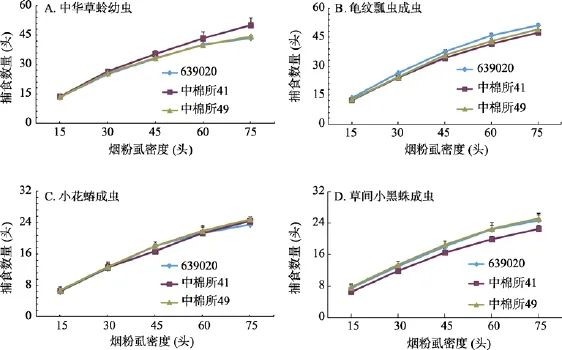
图3 3种棉田4种天敌对烟粉虱成虫捕食功能反应
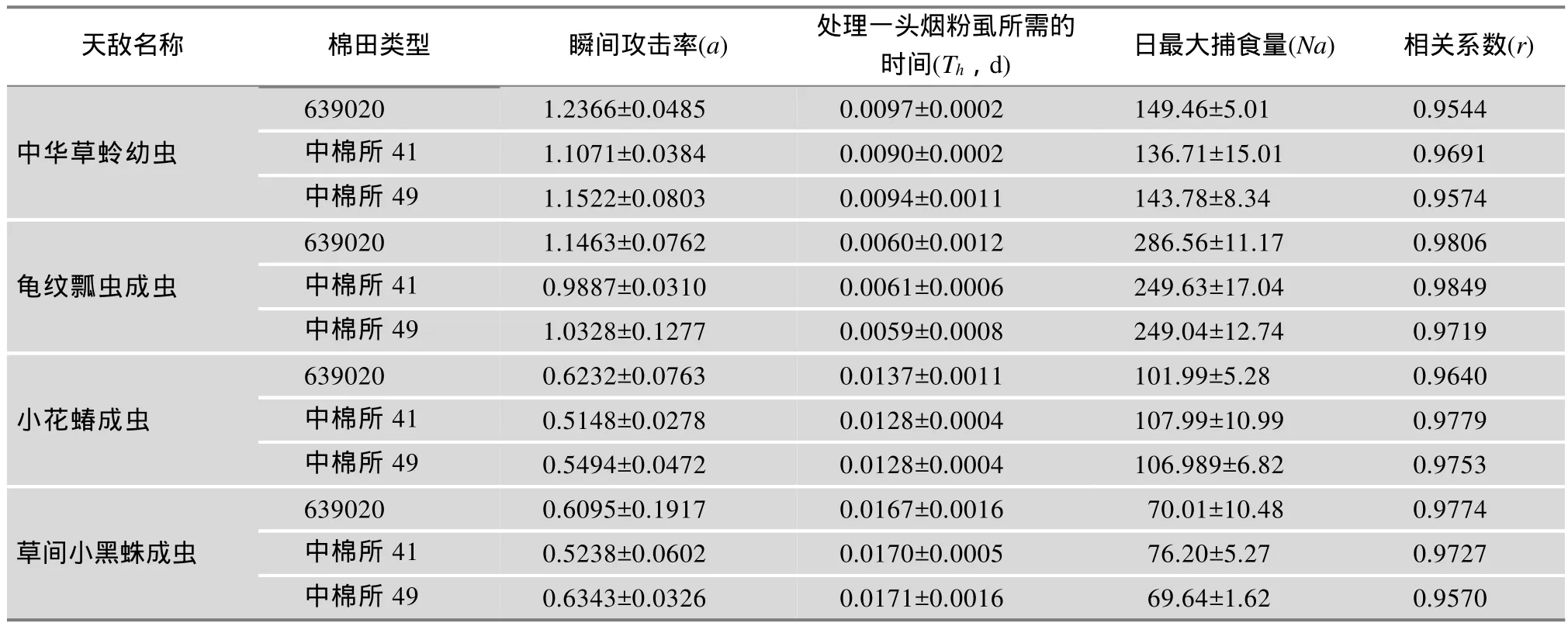
表2 3种棉田天敌对烟粉虱捕食功能参数
639020和中棉所41棉田天敌的捕食功能系数、瞬间攻击率、日最大捕食量较常规棉中棉所49之间无显著差异,也没有明显的规律,表明转639020棉和Bt棉在本文实试验条件下没有引起天敌捕食功能的变化。
3 讨 论
本研究结果表明,转Cry1Ac+Cry2Ab棉田在3个棉铃虫发生高峰期,棉铃虫幼虫数量均在防治指标以下,室内对靶标害虫也具有较好的控制作用;表明转Cry1Ac+Cry2Ab棉不仅对棉田靶标害虫具有良好的控制效果,同时在本实验条件和实验期内对天敌没有产生显著影响,与本课题组前期研究结果[21,22]在转高产基因和转Bt棉田的结论一致;长期生存在转Cry1Ac+Cry2Ab棉田的中华草蛉、龟纹瓢虫、小花蝽和草间小黑蛛对烟粉虱的捕食功能没有明显的变化,与张安盛等[23,24]、崔金杰等[25]和荆英等[26]的研究结果基本一致。
转基因抗虫棉大面积种植后,其抗虫基因对靶标害虫的控制作用明显,但靶标害虫为植食性害虫,其也是棉田天敌的食物。因此,转基因棉田靶标害虫数量的变化,则会引起食物链上天敌等害虫的变化,进而引起昆虫群落内部节肢动物群落种群动态或种类变化,甚至可能会影响到棉田节肢动物优势种的演替。可见转新型基因棉田节肢动物群落物种优势种和优势度的演替规律需进一步系统研究。
外来生物在自然状态或人类作用下在异地可以生长和繁殖的现象就是外来生物的入侵。外来生物入侵必然引起本土生物的变化,引起其食物和空间上的重新分布,因此外来生物入侵在生物多样性丧失或变化的过程中扮演着重要的角色,对一个生态区或多个生态区乃至整个生态系统生物多样性的完整性会造成极大的威胁[27]。近年来烟粉虱已入侵我国各大棉区并在我国大面积长期定居,且烟粉虱的植物寄主非常繁多和复杂。目前随着转基因棉花种植时间的延长和种植面积的扩大,抗虫棉田棉铃虫用药防治次数减少,烟粉虱得不到及时兼治或防治,已经成为我国重要的棉田害虫[12,15]。由于烟粉虱的寄主种类多,生长繁衍速度快,如不尽快找到防治棉田烟粉虱的有效方,一旦种群扩大且治理无方,不紧棉花生产造成巨大损失,烟粉虱将会转主至其他寄主植物如麻、瓜、豆角等,造成其他农业生产的巨大损失。因此棉田充分发挥天敌资源对烟粉虱的自然控制作用,是烟粉虱科学防治的重要环节,在建立烟粉虱防控体系中发挥重要的作用。本文在室内研究结果表明,中华草蛉、龟纹瓢虫对烟粉虱具有很强的捕食和控制潜力,但由于棉田自然环境因素复杂,天敌生存条件的多元化,烟粉虱活动范围及活动能力等均会在天敌捕食害虫过程和效果过程中起到一定的干扰作用,需进一步在棉田环境条件下研究自然天敌对烟粉虱的捕食能力。
[1]James,ISAAA,2007,http://www.isaaa.org.
[2]梁革梅,谭维嘉,郭予元.棉铃虫对Bt的抗性筛选及交互抗性研究.中国农业科学,2000,33(4):46–53.
[3]Shen JL,Lu P,He DJ,Zhou WJ,Chen J,Meng FX,Zhou XM,Su JY,Gao CF,Cen HM,Xu L,Pu QH.Monitoring of Bt resistance in CBW in China.Proceedings of the 15ThInternational Plant Protection Congress,2004:285.
[4]许新军.棉铃虫对Bt毒素—Cry1Ac抗性的生化及分子机理研究[学位论文].南京:南京农业大学,2005.
[5]Meng FX,Shen JL,Zhou WJ,Cen HM.Long-term selection for resistance to transgenic cotton expressing Bacillus thuringiensis toxin in Helicoverpa armigera (Hübner)(Lepidoptera:Noctuidae).Pest Manag Sci,2004, 60(2):167–172.
[6]Akhurst RJ,James W,Bird LJ,Beard C.Resistance to the Cry1Ac delta-endotoxin of Bacillus thuringiensis in the cotton bollworm,Helicoverpa armigera(Lepidoptera: Noctuidae).J Econ Entomol,2003,96(4):1290–1299.
[7]雒珺瑜,崔金杰,张帅,王春义,辛惠江.转Cry1Ac+ Cry2Ab基因棉对棉蚜生命表参数及种群动态的影响.应用昆虫学报,2012,49(4):906–910.
[8]雒珺瑜,崔金杰,张帅,陆雪君.转cry1Ac+cry2Ab基因棉对棉田节肢动物群落的影响.植物保护,2011,37(6): 90–92,111.
[9]雒珺瑜,崔金杰,辛惠江.转Cry1Ac+Cry2Ab基因棉花对天敌生长发育的影响.中国棉花,2011,38(12):11–12.
[10]Li HB,Wu KM,Xu Y,Yang XR,Yao J,Wang F. Population dynamics of pest mirids in cotton filed in Southern Xinjiang.Chinese Bull Entomol,2007,44(2): 219–222.
[11]Guo JY,Zhou HX,Wan FH,Liu XJ,Han ZJ.Population dynamics and damage of Lygus lucorum Mayr in Bt cotton fields under two control measures.Chinese Bull Entomol, 2005,42(4):424–428.
[12]Lu YH,Wu KM,Jiang YY,Guo YY,Desneux N. Widespread adoption of Bt cotton and insecticide decrease promotesbiocontrolservices.Nature,2012,487(7407): 362–365.
[13]Lu YH,Qiu F,Feng HQ,Li HB,Yang ZC,Wyckhuys KAG,Wu KM.Species composition and seasonal abundance of pestiferous plant bugs(Hemiptera:Miridae) on BtCotton in China.Crop Prot,2008,27(3–5): 465–472.
[14]Lu YH,Wu KM.Mirid bugs in China:pest status and management strategies.Outlooks Pest Manag,2011,22(6): 248–252.
[15]Lu YH,Wu KM,Jiang YY,Xia B,Li P,Feng HQ, Wyckhuys KAG. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in china.Science,2010,328(5982):1151–1154.
[16]Wu KM,Li W,Feng HQ,Guo YY.Seasonal abundance of the mirids,Lybux lucorum and Adelphocoris spp.(Hemiptera: Miridae)on Bt cotton in northern China.Crop Prot,2002, 21(10):997–1002.
[17]Wu KM,Guo YY.Theevolution ofcotton pest management practices in China.Annu Rev Entomol,2005,50: 31–52.
[18]WuKM,LuYH,FengHQ,JiangYY,ZhaoJZ. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton.Science,2008, 321(5896):1676–1678.
[19]张芝利.关于烟粉虱大发生的思考.北京农业科学, 2000,18(增刊):1–3.
[20]中华人民共和国农业部.2007.农业部 953号公告-12.2-2007(转基因植物及其产品环境安全检测 抗虫棉花 第2部分:生存竞争能力)http://www.moa.gov.cn/ zwllm/tzgg/gg/200801/t20080110_952066htm.
[21]雒珺瑜,刘传亮,张帅,王春义,吕丽敏,李春花,李付广,崔金杰.转RRM2基因棉生长势和产量及对棉田节肢动物群落的影响.植物生态学报,2014,38(7):785–794.
[22]雒珺瑜,王春义,辛惠江,崔金杰.转基因棉田节肢动物群落调查时间间隔的探讨.棉花学报,2013,25(5):440–445.
[23]张安盛,李丽莉,于毅,门兴元.中华草蛉幼虫对西花蓟马若虫的捕食功能反应与搜寻效应.植物保护学报, 2007,34(3):247–251.
[24]张安盛,于毅,李丽莉,张思聪.东亚小花蝽(Orius sauteri)成虫对入侵害虫西花蓟马(Frankliniella occidentalis)成虫的捕食作用.生态学报,2007,27(5):1903–1909.
[25]崔金杰,雒珺瑜,王春义,李树红,李春花.转双价基因(Bt+CpTI)棉对棉田主要捕食性天敌捕食功能反应的影响.南京农业大学学报,2005,28(1):48–51.
[26]荆英,黄建,韩巨才,马瑞燕.刀角瓢虫对烟粉虱的捕食作用.植物保护学报,2004,31(3):225–229.
[27]黄红娟,叶万辉.外来种入侵与物种多样性.生态学杂志,2004,23(2):121–126.
(责任编委:黄原)
The effects of transgenic Cry1Ac+Cry2Ab cotton on cotton bollworm control and functional response of predators on whitefly
Junyu Luo,Shuai Zhang,Limin Lü,Chunyi Wang,Xiangzhen Zhu,Jinjie Cui
State Key Laboratory of Cotton Biology,Cotton Research Institute,Chinese Academy of Agricultural Sciences,Anyang 455000, China
In this study,we detected and clarified the roles of transgenic Cry1Ac+Cry2Ab cotton“639020”in controlling cotton bollworm(Helicoverpa armigera)during critical periods of bud stage(second generation of bollworm),flowering stage(third generation of bollworm)and bolling stage(fourth generation of bollworm)as well as the influences of 639020 cotton on functional response of the main predators(Chrysopa sinica larvae, Propylaea japonica,Orius and Erigonidium graminicola)on whitefly using transgenic Cry1Ac cotton“CCRI41”and conventional cotton“CCRI49”as the control.Our results showed that the 639020 cotton well controlled the second and third generation of bollworm,and the level of insect resistance increased by 52.85%and 16.22% separately compared with that of CCRI41,with a significant effect on the second generation of bollworm.Moreover, the number of bollworm eggs in 639020 cotton field was lower than that in CCRI41 and CCRI49 cotton fields(except the second generation of bollworm)during the cotton bud,flowering and bolling stages.Although the number of bollworm larvae in 639020 cotton field was significantly lower than that in CCRI49 field,and both under the controlling index,it has no significant difference compared with that in CCRI41 cotton field.There were also no obvious changes in predator functions of Chrysopa sinica,Propylaea japonica,Orius and Erigonidium graminicola on bemisia tabaci between 639020,CCRI41 and CCRI49 cotton filed.This study evaluated the safety of new transgenic cotton on environment, anti-insect activity of exogenous gene and the safety of production and application prospect.
transgenic Cry1Ac+Cry2Ab cotton;Helicoverpa armigera;predator function
2014-11-30;
2015-01-29
转基因生物新品种培育重大专项(编号:2014ZX08011-002)资助
雒珺瑜,学士,副研究员,研究方向:转基因生物安全。E-mail:luojunyu1818@126.com
崔金杰,博士,研究员,研究方向:转基因生物安全。E-mail:cuijinjie@126.com
10.16288/j.yczz.14-418
时间:2015-3-11 9:20:02
URL:http://www.cnki.net/kcms/detail/11.1913.R.20150311.0920.004.html
