基于脯氨酸合成积累的甘蔗分蘖期耐旱生理效应分析
2015-09-21杨杰文叶昌辉莫俊杰周鸿凯
杨 善,杨杰文,叶昌辉,莫俊杰,周鸿凯
(广东海洋大学农学院,广东 湛江 524088)
基于脯氨酸合成积累的甘蔗分蘖期耐旱生理效应分析
杨善,杨杰文,叶昌辉,莫俊杰,周鸿凯
(广东海洋大学农学院,广东 湛江 524088)
为探究干旱胁迫下分蘖期甘蔗植株内游离脯氨酸积累及其合成关键酶的响应机制,以甘蔗品种ROC22为材料进行桶栽干旱胁迫实验,测定生长于7种不同土壤水分下的甘蔗叶片中丙二醛含量、电导率、游离脯氨酸含量以及δ-OAT、P5CS、SOD的活性。结果表明,随着干旱胁迫强度增大,植株体内不同程度地激活了P5CS、δ-OAT、SOD的活性,合成积累了大量的游离脯氨酸,以抵抗丙二醛等氧化物的积累所造成对细胞膜透性的破坏。但在沙土的土壤水分为2.64%(相对持水量21.2%)时,达到了ROC22的萎蔫系数。相关统计分析结果,6个甘蔗生理指标间的简单正相关均达到了显著水平。偏相关分析结果,细胞膜透性分别与P5CS活性、脯氨酸含量为负的偏相关且达到了显著水平,丙二醛、δ-OAT、SOD分别与脯氨酸含量的偏相关系数均到达了极显著水平。此外,P5CS与SOD存在共线性,多元相关性分析表明P5CS对游离脯氨酸积累的作用显著地大于δ-OAT,在干旱胁迫下,甘蔗脯氨酸的两个合成途径表现为以Glu→Pro途径主,Orn→Pro途径为辅。
甘蔗;脯氨酸;P5CS;δ-OAT;干旱胁迫。
农业生产常受到逆境条件的影响,其中干旱和盐渍引起的渗透胁迫对农作物的生长和产量影响最大。作物通过长期的自然进化和人工选育,已经形成了许多抵抗外界逆境条件的机制[1]。在水分胁迫下植物首先受害的是细胞膜系统,过氧化作用导致细胞衰老,其主要氧化产物丙二醛(MDA)含量明显增加,质膜透性(PMP)增大,离子外渗,同时大量积累游离脯氨酸(Pro)[2];而脯氨酸的增加有助于保持细胞或组织的持水能力,提高农作物在干旱和盐渍胁迫条件下的适应性,是获得高产和充分利用自然资源的重要途径[3]。
甘蔗(Saccharum officinarum)是中国重要的糖料和能源作物,目前主要蔗区分布在桂中南、滇西南、粤西和海南等南方黄、红壤地区,而且90%以上的种植面积为缺乏灌溉的旱坡地,季节性干旱是限制这些地区甘蔗生产的首要环境因素[4]。因此,甘蔗抗旱耐旱生理亦为研究热点之一,陈如凯等[5]研究表明,干旱胁迫下甘蔗体内脂质过氧化作用加剧,质膜透性增加,耐旱性弱的品种变化尤为明显。在反复干旱胁迫处理下,不同甘蔗品种的生长势、叶片水势及叶绿素含量均有不同程度降低,叶片脯氨酸和丙二醛含量升高,叶片细胞质膜透性增大[6]。黄诚梅等[7]用3个甘蔗品种于伸长期以PEG胁迫1-6d模拟干旱胁迫的研究表明,胁迫条件下甘蔗叶片的脯氨酸含量明显上升的趋势、△1-吡咯啉-5-羧酸合成酶(P5CS)活性显著提高,并表现为品种间的差异显著,但δ-鸟氨酸转氨酶(δ-OAT)活性的变化不明显。刘洋等[8]认为,干旱胁迫下甘蔗及其近缘植物的抗氧化保护酶系统如过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、超氧化物歧化酶(SOD)活性及可溶性蛋白含量等都会发生不同的变化。然而,针对干旱胁迫下甘蔗体内基于游离脯氨酸积累及其合成关键酶的抗旱生理效应方面的研究未见报道。本研究以甘蔗品种ROC22为材料,测定干旱胁迫下甘蔗叶片的游离脯氨酸、丙二醛含量、电导率 (Cond),及P5CS、δ-OAT、SOD活性等6个生理生化指标,旨在探索甘蔗分蘖期植株内游离脯氨酸积累及其合成关键酶的响应机制,为甘蔗抗旱耐旱机制的深入研究,以及旱坡地高产高糖栽培技术的创新提供参考。
1 材料与方法
1.1实验材料与实验设计
参试材料为粤西蔗区主推品种ROC22,采用桶栽实验方法,实验土壤为砾质沙土(田间最大持水量为12.5%)。选用高50 cm、口径40 cm的塑胶桶(距桶底10 cm处对称打4个小孔),每桶装干砾质沙土8.0 kg,共6桶。于2013年9月11日,将18段甘蔗双芽苗按每桶3段分别种入6个桶内,种苗入土深度为10 cm,浇水保持土壤为湿润状态,12 d后萌芽,每桶浇淋500 mL的0.2 g/mL荷兰复合肥营养液,往后每隔2 d浇淋自来水500 mL。于2013年10月26日下午,将桶栽甘蔗搬入玻璃温室内进行干旱处理,不再浇水。2013年10月27日,甘蔗出苗后34 d,处于分蘖期,开始第1次取样(即干旱处理0 d),以后每2天(即48 h)取样1次,连续取样7次,即7种水分水平。每次每桶取一个样,均在9:30—10:00时段内剪取样本(-1叶与+1叶)进行实验测试分析,同时取5~10 cm土层的土样测定土壤含水量。
1.2测定方法
取-1叶进行丙二醛含量、植物组织细胞通透性测定、SOD活性测定;取+1叶进行脯氨酸含量、P5CS活性、δ-OAT活性测定。丙二醛的含量测定按高俊凤[9]的方法;植物组织细胞通透性的测定采用电导法[9]进行;土壤含水量的测定采用烘干法[10]进行测定; P5CS的抽提按照Kavi等[11]的方法,其活性测定参照黄诚梅[12]的方法进行,一个酶活性单位(U)定义为每分钟生成1 μmol γ-谷氨酰胺(湿样)所需要的酶量,单位为U·g-1·min-1;δ-OAT的抽提按照Delauney等[13]的方法进行,其活性测定按照Kim等[14]的方法进行,一个酶活性单位(1 U)定义为1 h生成1 mmol P5C所需要的酶量(湿样),单位为U·g-1·h-1;脯氨酸含量的测定按照Bates等[15]的方法;SOD活性测定依据南京建成生物工程研究所生产的试剂盒测定,试剂配制按照说明书操作。
1.3统计分析方法
应用统计分析软件SPSS 19.0进行数据的统计分析,用Duncan法测定处理间各生理指标的差异显著性(P<0.05),并在图中进行标注。
2 结果与分析
2.1土壤含水量
由图1可见,随着干旱处理天数的增加,土壤含水量按一定的梯度下降,并且在每个梯度之间均有显著性的差异,其相应的土壤含水量梯度为11.72%、9.96%、8.86%、6.52%、4.96%、3.65%、2.64%。表明本研究的干旱胁迫梯度设计具有代表性和可靠性,该水分的梯度即可作为显著性差异的干旱胁迫水平而进行相应指标的测定与分析。

图1 干旱胁迫下土壤含水量的变化Fig.1 Changes in water content of soil
2.2甘蔗干旱胁迫生理效应分析
随着干旱胁迫强度的增加,甘蔗叶片内游离脯氨酸的含量、P5CS活性、δ-OAT活性、SOD活性等而增加,其中脯氨酸含量在干旱处理第4天开始有显著性地增加,第8天急剧增加,第10天达到游离脯氨酸累积量的高峰值,为第一次取样时(土壤含水量为11.72%)的6.5倍,此后即开始显著性减少(图2_a)。
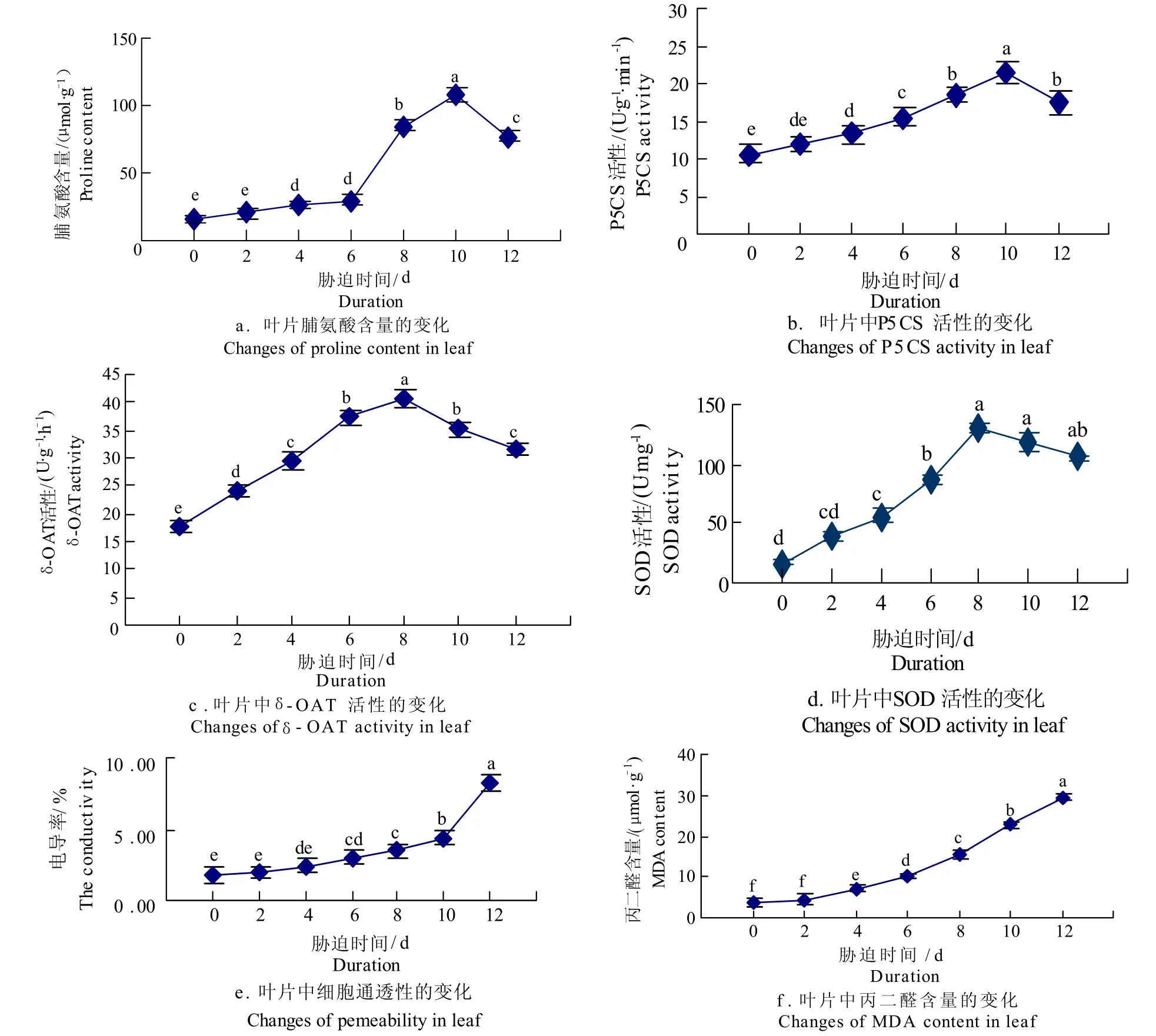
图2 干旱胁迫下各项生理指标Fig.2 Various physiological indicators under drought stress
P5CS活性和SOD活性在干旱胁迫下有相似的增长趋势,分别在处理后第4、6天开始有显著性地增加,在第10天和第8天达到峰值,为第一次取样时的2.05和7.2倍,随后P5CS活性开始显著下降,而SOD活性则缓慢下降(图2_b,2_d)。δ-OAT活性在干旱处理开始即显著增加,到处理后的第8天到达峰值,之后开始显著下降(图2_c)。细胞膜透性和丙二醛含量在干旱处理开始时呈现缓慢递增,在处理后的第8天电导率开始显著增加,在第12天时急剧增加;处理后第4天开始,丙二醛含量持续显著增加,并随着处理时间而增量扩大 (图2_e,2_f)。说明在干旱的胁迫下,甘蔗植株内不同程度地激活了P5CS、δ-OAT的活性,合成积累了大量的游离脯氨酸,同时也激活了SOD活性,以抵抗干旱胁迫下甘蔗植株内丙二醛等氧化物的积累而所造成对细胞膜透性的破坏。
2.3甘蔗干旱胁迫生理指标间的相关性分析
由表1可知,甘蔗干旱胁迫的6个生理指标间的简单相关系数均到达了显著水平,表明了这6个生理指标间均有密切的相关关系。同时可见,脯氨酸与P5CS、MDA与Cond、P5CS与SOD的相关系数达到0.91以上,表明这些因素之间相当密切,而在多元回归分析时可能存在共线性现象。而偏相关分析表明,MDA、Cond、δ-OAT、SOD等与脯氨酸的偏相关系数均到达了极显著水平,其中SOD为最大,Cond和δ-OAT为负向偏相关性。另外,MDA与Cond、P5CS,Cond与P5CS,SOD与P5CS、δ-OAT的偏相关性达到显著水平。
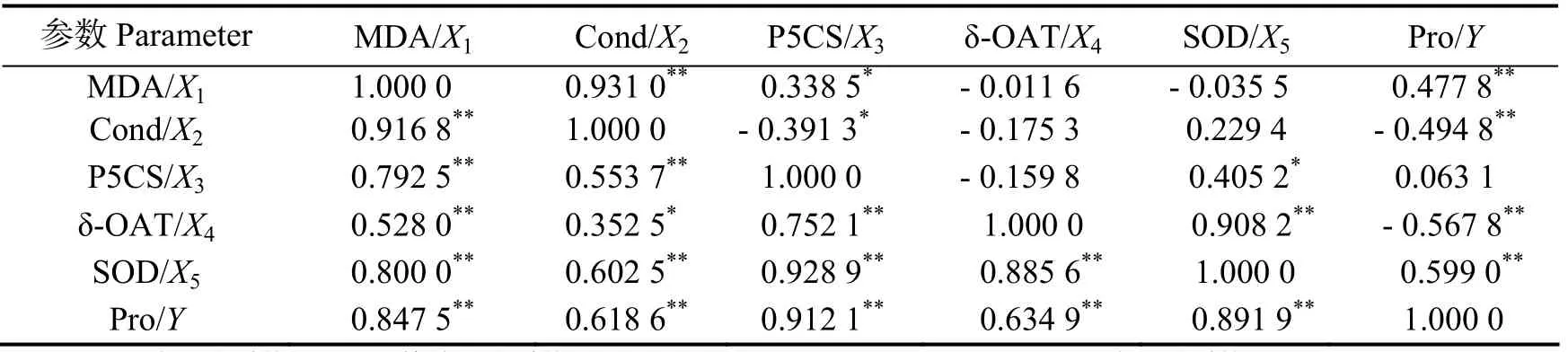
表1 各个生理指标与脯氨酸含量的简单、偏相关性分析Table 1 Simple correlation analysis and partial correlation analysis of physiological indexes of sugarcane under drought stress
从表2可知,脯氨酸与其他5个生理指标多元回归分析的共线性诊断分析,No.4的特征值为0.019 8,由此可知这5个生理指标之间存在共线性现象。结合简单相关分析的结果,即进行脯氨酸与P5CS、SOD的多元回归的共线性诊断分析及其通径分析如表3和表4。

表2 Y与5个X因素多元回归分析的共线性诊断Table 2 Y and five X-factors in multiple regression analysis of collinearity diagnostics
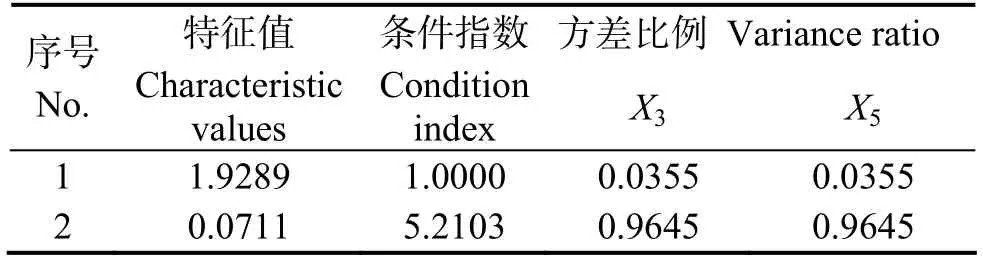
表3 Y与X3、X5的多元回归的共线性诊断Table 3 Y and X3, X5in multiple regression analysis of collinearity diagnostics
由表3可知,P5CS与SOD之间确实存在共线性现象,表明了表1中出现的P5CS对脯氨酸偏相关系数较低的状况,是由于P5CS与SOD之间存在的共线性所造成。
由表4的分析结果表明,P5CS和SOD因素对脯氨酸均有极显著的偏相关关系,而且P5CS对脯氨酸的偏相关关系比SOD因素的还大。P5CS对脯氨酸的直接通径系数为0.609 3,通过SOD的系数为0.302 8,而SOD对脯氨酸的直接通径系数为0.326 0,通过P5CS的系数为0.566。

表4 Y与X3、X5的多元回归及通径分析Table 4 Complex correlation analysis of Y and X3, X5
表5可见,各个指标对脯氨酸的直接通径系数中,SOD为1.093 8,其对脯氨酸的正向直接作用最大;而Cond与δ-OAT对脯氨酸有显著的负向作用。由于P5CS与SOD存在共线性现象,虽然P5CS对脯氨酸的直接通径系数较小,但通过SOD和MDA对脯氨酸的间接通径系数均较大,分别为0.970 5、0.571 5;剩余通径系数仅为0.063。
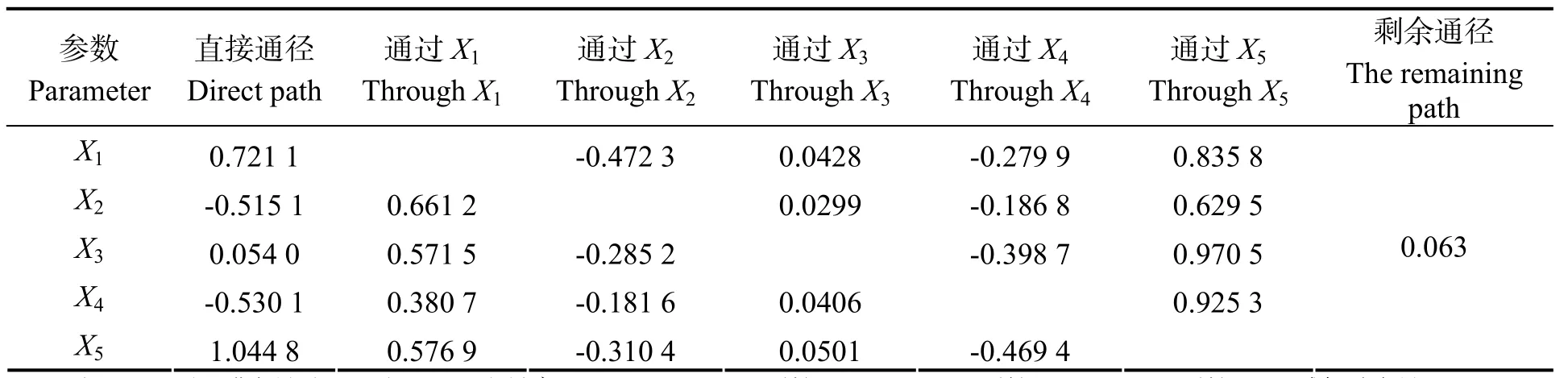
表5 各个生理指标对脯氨酸的通径分析Table 5 Path analysis of physiological indexes to proline
2.4植株游离脯氨酸与P5CS、δ-OAT相关性分析
从表6、表7可知, P5CS和δ-OAT对脯氨酸的偏相关系数分别为0.854 2和-0.197 2,P5CS对脯氨酸的偏相关系数达极显著水平;P5CS对脯氨酸的直接通径系数为1.004 7,通过δ-OAT的系数为-0.0927,而δ-OAT对脯氨酸的直接通径系数为-0.123 0,通过P5CS的系数为0.757 0。这一结果表明,在干旱胁迫下,甘蔗植株脯氨酸的合成积累过程,P5CS对脯氨酸含量的作用显著地大于δ-OAT的作用。同时,剩余通径系数为0.161 6,说明还有随机因素对植株脯氨酸含量有较大的影响。

表6 P5CS、δ-OAT与脯氨酸含量的简单、偏相关性分析Table 6 P5CS, δ-OAT and proline of simple correlation analysis and partial correlation analysis

表7 P5CS、δ-OAT与脯氨酸的通径分析Table 7 Path analysis of P5CS, δ-OAT and psroline
3 讨 论
3.1干旱胁迫下甘蔗分蘖期的脯氨酸积累效应
甘蔗长期生长在热带、亚热带的环境中,往往可通过形态、生理生化等多种途径的变化来维持干旱胁迫下体内的水分平衡,其中生理抗旱一直是研究的热点。周鸿凯等[16]的研究认为,甘蔗抗旱耐旱能力有两种类型:第一种为具有强大的吸收和转运功能系统,其缺水临界值较低,从而表现好的抗旱性;第二种为靠迅速积累游离的脯氨酸来抵抗不良的条件,而表现抗旱力好。本研究结果表明,当沙土的土壤含水量低于9.96%时(相对持水量为79.6%),甘蔗植株内P5CS、δ-OAT的活性即被显著地激活了,合成积累了大量的游离脯氨酸,同时也激活了SOD活性,以抵抗干旱胁迫下甘蔗植株内丙二醛等氧化物的积累而所造成对细胞膜透性的破坏。从表面上看,似乎ROC22属于第二种类型,靠迅速积累大量脯氨酸来抵抗不良环境。若要准确判定ROC22属于哪一种抗旱类型,还有待与其它甘蔗品种进行深入的对比探究。有研究表明,MDA含量高低和细胞质膜透性变化是反映细胞膜脂过氧化作用强弱和质膜破坏程度的重要指标,也是反映水分胁迫对甘蔗造成伤害的重要参数[17],高三基等[18]研究也认为MDA、SOD等可作为甘蔗抗旱性鉴定指标。本研究在沙土的土壤水分低于2.64%(相对持水量为21.2%)时,甘蔗叶片的细胞膜透性和丙二醛含量急剧地增大,同时,P5CS、δ-OAT、SOD等的活性急剧地下降,游离脯氨酸的积累量亦开始显著地减少,表明此时植株已经无法抵抗该干旱强度的胁迫。由此可见,在砾质沙土中土壤水分为2.64%(相对持水量为21.2%)时,已到达了ROC22的萎蔫系数。
3.2干旱胁迫下甘蔗分蘖期生理指标间的相关性
本研究结果表明,干旱胁迫下6个甘蔗生理指标间的简单正相关均达到了显著水平,说明了甘蔗分蘖期植株体内的这些生理生化过程间关系密切,甘蔗受到干旱胁迫时,细胞内的渗透压增加,质膜产生过氧化作用,MDA含量提高,使得质膜受损透性增加,而促使游离脯氨酸的合成积累,以降低细胞内的渗透压,同时SOD活性也增大,用于清除O2-、·OH等活性氧(ROS)的积累,保持植株的生长发育。这与陈如凯等[4]、梁丽琼等[19]以甘蔗为材料的研究结论类似。进一步的偏相关分析表明,细胞膜透性分别与P5CS活性、脯氨酸含量为负的偏相关且达到了显著水平。毛才良等[20]的研究认为,大麦叶片细胞电导率上升是由于细胞电解质外渗,而细胞电解质的外渗主要是离子胁迫所致。同理,本研究甘蔗由于水分胁迫,导致细胞电解质外渗,电导率增大,细胞膜透性增大,P5CS活性增强,生物体内合成更多的脯氨酸,从而细胞液溶质含量变大,减少细胞内电解质外渗,降低了电导率。同时,丙二醛、δ-OAT、SOD分别与脯氨酸含量的偏相关系数均到达了极显著水平,其中δ-OAT与脯氨酸为负向偏相关,说明干旱胁迫下,甘蔗脯氨酸含量多了,会使得δ-OAT活性降低,有反馈调节的作用。
3.3干旱胁迫下甘蔗分蘖期体内游离脯氨酸的合成积累途径
植物脯氨酸合成、累积及代谢是一个受非生物胁迫和细胞内脯氨酸浓度高度调控的生理生化过程[21]。植物体内脯氨酸的合成分为两条生物合成途径[22],其中△1-毗咯琳-5-羧酸合成酶(P5CS)和δ-鸟氨酸转氨酶(δ-OAT)分别是脯氨酸合成途径中谷氨酸途径[23]和鸟氨酸途径[23]的关键酶。在盐、干旱、重金属等逆境胁迫下,哪一种途径对脯氨酸积累的影响占据主导地位,则与植物的种类、生理状态和植物体内的氮素水平有关[24]。Roosens等[25]研究不同生育期植物脯氨酸积累途径发现,幼小植株以Orn途径为主,成年植株以Glu途径为主,而在渗透胁迫条件下两条途径发挥同样重要的作用。本研究中,以游离脯氨酸为因变量、其他5个生理指标为自变量的通径分析结果表明,各个指标对游离脯氨酸的直接通径系数中,SOD对游离脯氨酸的正向直接作用最大,而细胞膜透性与δ-OAT对游离脯氨酸表现为负向的直接作用。由于P5CS与SOD存在共线性现象,虽然P5CS对Y的直接通径系数较小,但通过SOD对游离脯氨酸具有较大的间接通径系数,说明P5CS对植株游离脯氨酸合成积累具有较大的作用。同时,干旱胁迫下甘蔗分蘖期叶片内的P5CS、δ-OAT活性均被显著地激活,植株游离脯氨酸与P5CS、δ-OAT的多元相关性分析表现为P5CS对游离脯氨酸积累的作用显著地大于δ-OAT。表明甘蔗在干旱胁迫下,P5CS、δ-OAT活性均被显著地激活,启动各自的途径合成积累游离脯氨酸,以抵制干旱胁迫所造成的破坏,表现为以Glu→Pro途径主,Orn→Pro途径为辅。这与黄诚梅等[6]用3个甘蔗品种于伸长期以PEG胁迫模拟干旱胁迫,认为甘蔗叶片中脯氨酸生物合成的谷氨酸合成途径比鸟氨酸合成途径更占优势地位的结论相似。而赵福庚等[26]的研究则认为,盐胁迫下可明显激活大麦幼苗体内脯氨酸合成的鸟氨酸途径,使得Orn →Pro途径成为脯氨酸合成积累的主要途径。
植物游离脯氨酸合成积累是一个十分复杂的生理生化过程,由许多生理生化反应及多个基因调控。本研究结果表明,尚有一些因子对游离脯氨酸的合成积累有影响。其他研究表明,除脯氨酸的合成酶受激活外,还有其它一些酶如脯氨酸运输酶[27]、谷氨酰胺合成酶[28]等也参与了脯氨酸的累积过程。因此,高等植物体内游离脯氨酸合成积累的机制,还有待于从植物细胞信号传递、脯氨酸运输、相关酶的作用机制等方面进一步研究证明。
[1]INES Slama,TAHAR Ghnaya,DORSAF Messedi,et al. Effect of sodium chloride on the response of thehalophyte species Sesuvium portulacastrum grown in mannitol-induced water stress[J]. J Plant Res,2007,120(2):291–299.
[2]OUZOUNIDOU G,ILIAS I F,GIANNAKOULA A, et al. Effect of water stress and NaCl triggered changes on yield,physiology,biochemistry of broad bean(Vicia faba)plants and on quality of harvested pods[J]. Biologia,2014,69(8):1010- 1017.
[3]RUTHERFORD R S. The assessment of proline accumulation as a mechanism of drought resistance in sugarcane[J]. Prec S Afric Sug Tech Asseci,1989,63:136-141.
[4]李富生,何丽莲. 植物对非生物胁迫的生理响应及甘蔗抗旱抗寒性研究进展[J]. 甘蔗,2004,11(1):31-37.
[5]陈如凯,张木清,陆裔波. 干旱胁迫对甘蔗生理影响的研究[J]. 甘蔗,1995,2(1):1-6.
[6]杨建波,诸葛少军,黎海涛,等. 干旱胁迫对甘蔗生长生理的影响及品种抗旱性评价[J]. 南方农业学报,2012,43(8):1114-1120.
[7]黄诚梅,毕黎明,杨丽涛,等. 聚乙二醇胁迫对甘蔗伸长期间叶中脯氨酸积累及其代谢关键酶活性的影响[J].植物生理学通讯,2007,43(1):77-80.
[8]刘洋,姚艳丽,林希昊,张海林,苏俊波. 干旱胁迫对甘蔗近缘材料抗氧化系统酶活性的影响[J]. 西南农业学报,2012,25(3):852-855.
[9]高俊凤. 植物生理学实验技术[M]. 西安:世界图书出版公司,2000.
[10]周钟瑜. 土壤水分测定方法[M]. 北京:水利水电出版社,1986:11-12.
[11]KAVI KISHOR P B,HONG Z L,MIAO G H, et al. Overexpression of △1-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants [J]. Plant Physiol,1995,108(4):1387-1394.
[12]黄诚梅. 甘蔗脯氨酸积累与△1-吡咯啉-5-羧酸合成酶(ScP5CS)基因克隆及转化研究[D]. 广西大学,2007:17-18.
[13]HU C-AA,DELAUNEY A J,VERMA D P S. A bifunctional enzyme(△1-pyrroline-5-carboxylate synthetase)catalyzes the first two steps in proline biosynthesis in plants[J]. Proc Natl Acad Sci USA 1992,89(19):9354-9358.
[14]Kim H R,Rho H M W,Park J W,et al. Assay of ornithine aminotransferase with ninhydrin[J]. Anal Biochem,1994,223(2):205-207.
[15]BATES L S,WALDREN R P,TEARE I D. Rapid determination of free proline for water-stress studies[J]. Plant and Soil,1973,39(1):205-207.
[16]周鸿凯,叶振邦. 干旱对甘蔗叶片的细胞透性及脯氨酸积累效应的研究初报[J]. 甘蔗糖业,1988(4):39-41.
[17]陈少裕. 膜脂过氧化与植物逆境胁迫[J]. 植物学通报,1989,6(4):211-217.
[18]高三基,罗俊,张华,等. 甘蔗抗旱性生理生化鉴定指标[J]. 应用生态学报,2006,17(6):1051-1054.
[19]毛才良,刘有良. 盐胁迫大麦苗体内的Na+、K+分配与叶片耐盐量[J].南京农业大学学报,1990,13(3):32-36.
[20]罗俊,张木清,林彦铨,等.甘蔗叶绿体荧光参数、MDA含量及膜透性与耐旱性的关系[J].福建农业大学学报,1999,28(3):257-262.
[21]KIYOSUE T,YOSHIBA Y,YAMAGUCHI SK. A nuclear gene encoding mitochondrial proline dehy-drogenase, an enzyme involved in proline metabolism,is upregulated by proline but downregulated by dehydration in Arabidopsis thaliana[J]. Plant Cell,1996,8(8):1323-1335.
[22]LEHMANN S,FUNK D,Szabados L. Proline metabolism and transport in plant development[J]. Amino Acids,2010,39(4):949-962.
[23]DELAUNEY A J,HU C-AA,KISHOR P B K,et al . Cloning of ornithine -aminotransferase cDNA from Vigna aconitifolia by trans-comple-mentation in Escherichia coliand regulation of proline biosynthesis[J]. J Biol Chem 1993,268(25):18673-18678.
[24]HERVIEU F,LE D L,HUAUL T C. Contribution of ornithine aminotransferase to proline accumulation in NaCl treated radish cotyledons[J]. Plant Cell Environ,1995,18(2):205-210.
[25]ROOSENS N H C J,THU T T, ISKANDAR H M. Isolation of ornithine- aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis[J]. Plant Physiol,1998,117(1):263-271.
[26]赵福庚,孙诚,刘友良. 盐胁迫激活大麦幼苗脯氨酸合成的鸟氨酸途径[J]. 植物学报,2001,43(1):36-40.
[27]IGARASHI Y,YOSHIBA Y,TAKESHITA T. Molecular cloning and characterization of a cDNA encoding proline transporter in rice[J]. Plant Cell Physiol,2000,41(6):750-756.
[28]BRUGIEREA N,DUBOISB F,LIMAMIA A M. Glutamine synthetase in the phloem plays a major role in controlling proline production[J]. Plant Cell,1999,11(10):1995-2012.
(责任编辑:陈庄)
Analysis of Drought toLerance Physiological Effect Based on Proline Biosynthesis and Accumulation at the Tillering Stage in Sugarcane
YANG Shan, YANG Jie-wen, YE Chang-hui, MO Jun-jie, ZHOU Hong-kai
(College of Agriculture, Guangdong Ocean University, Zhanjiang 524088,China)
To explore the responses of free proline accumulation and its synthesis key enzymes at the tillering stage in sugarcane under drought stress,sugarcane variety ROC22 as materials was cultivated in bucket for drought stress test. The malondialdehyde content, conductivity, free proline content and the activity of δ-OAT, P5CS and SOD were measured under the seven soil moisture levels in sugarcane leaves.The results showed that when drought stress strength increased, the activity of P5CS, δ-OAT and SOD were activated in different degree, synthesized and accumulated a largeamount of free proline, to resist the damage caused by accumulation of MDA and other oxides for the cell membrane permeability. But when soil moisture was 2.64% (relative water capacity was 21.2%) in sandy soil, the wilting coefficient reached up to ROC22. Statistical analysis of the results showed that thesimple correlation between 6 physiological indexes of sugarcane had reached a significant level. Further partial correlation analysis showed that the cell membrane permeability and P5CS, proline content were negative partial correlation and reached a significant level. MDA, δ-OAT, SOD and proline content of partial correlation coefficients had reached a significant level. Meanwhile, SOD and P5CS were collinear and the multiple correlation analysis showed that the effects excertedon proline accumulation by P5CS was significantly greater than that by δ-OAT, and it appeared that the glutamic acid pathway (Glu→Pro)was the main pathway, and ornithine pathway(Orn→Pro)supplemented under drought stress in sugarcane.
sugarcane;proline;P5CS;δ-OAT;drought stress
S566.1
A
1673-9159(2015)06-0087-07
10.3969/j.issn.1673-9159.2015.06.016
2015-06-24
国家自然科学基金(41073059);广东省科技计划项目(2013B020301005,2013B020301008);广东省良种培育和引进专项(201201148)
杨善(1987—),男,硕士研究生,研究方向为热带作物栽培生理。
周鸿凯(1962—),男,研究员,研究方向为作物学。E-mail: 897961801@qq.com
