57NG208 与南涧果蔗正反交后代的遗传变异分析
2015-07-13朱建荣边芯郎荣斌俞华仙冯蔚桃联安董立华安汝东经艳芬
朱建荣,边芯,郎荣斌,俞华仙,冯蔚,桃联安, 董立华,安汝东,经艳芬*
(1.云南省农业科学院甘蔗研究所瑞丽育种站,云南 瑞丽678600;2.云南甘蔗遗传育种重点实验室,云南 开远661600)
现代甘蔗品种作为高度杂合异源多倍体作物,至少含3 ~5个种的血缘[1],基本上是POJ 和Co 系列品种的近亲杂交后代[2],品种间遗传基础相近。中国自育甘蔗品种的细胞质源非常有限,仅来自斑扎马新黑谭(Bandjarmasin Hitan)、黑车里本(Black Cheribon)、拔地拉(Badila)、克林斯他琳娜(Crystalina)和卡路打不廷(Kaludai Boothan),且以前2个种为主[3]。用于拓宽甘蔗遗传基础的野生种有斑茅[4]、滇蔗茅[5–7]、蔗茅[8]、细茎野生种[9]、大茎野生种[10]等。大茎野生种的长势旺盛,茎硬抗风,抗虫,宿根性好,低糖,高纤维。国内外甘蔗育种机构利用大茎野生种育成的优良品种(系)有美国夏威夷H37–1933,中国台湾PT43–52、F146、F152、ROC22、昆士兰32MQ,美国运河点育种场CP36–138,中国广州粤糖79–177[11]等。关于细胞质对作物性状[12]、抗病[13–17]、抗虫[18]、抗倒性[19]及抗旱性[20]等方面的研究已有报道。细胞质遗传与细胞核遗传不同,主要表现为母系遗传,其抗性不分离,遗传连续性和稳定性比较高[21],因此,将甘蔗野生种作为细胞质来源可以增强甘蔗的抗性和适应性。
关于大茎野生种创新种质主要性状遗传变异的研究较少。本研究中利用大茎野生种57NG208和中国种南涧果蔗进行杂交,获得正交、反交F1后代群体,通过SSR 分子标记对后代进行真实性鉴定,选取132个真实性后代进行聚类分析,并对其重要农艺性状的遗传变异进行研究,探讨2个原种分别做母本对后代的影响及其后代相关性状的遗传变异情况,以期为中国种、大茎野生种的细胞质利用提供参考依据。
1 材料与方法
1.1 材料
以云南省农业科学院甘蔗研究所瑞丽育种站保育的大茎野生种57NG208(以下简称P1)和中国种南涧果蔗(以下简称P2)为亲本进行杂交(P1×P2为正交,P2×P1为反交)。于2012年杂交季节获得2个群体的后代材料,于2012年3月10日进行杂交花穗种子播种,4月20日假植,2012年5月29日定植于大田。大田随机区组排列。每个组合随机选取9株,3次重复。每重复种植1 行,行长6.5 m,行距1.1 m,其管理与大田一致,2013年1月26日调查农艺性状。
1.2 叶片基因组DNA 的提取与SSR 分析
以+1 叶片为样本,叶片基因组DNA 提取参照文献[22]中的方法。SSR 引物选用文献[23]中多态性好 的 MSSCIR21 、 MSSCIR26 、 MSSCIR36 和SMC336BS 等共4 对引物。反应体系、PCR 扩增条件的确定参照文献[24]中的方法。扩增产物经95℃变性后,在5%的变性聚丙烯酰胺凝胶上进行电泳分离,凝胶染色采用文献[25]中的快速银染法。
1.3 数据分析
分子数据采用人工读带的方式,每对SSR 引物检测1个位点,在相同迁移位置上,有带记为1,无带记为0,每条带相当1个等位基因,建立0–1 矩阵。采用软件NTSYSpc 2.1 计算出Jaccard 遗传相似性系数(GS)[26],并根据相似性系数进行UPGMA 聚类分析,构建聚类图。相关计算公式如下:
变异系数(CV)=标准差/后代平均值;
遗传传递力(Ta)=后代平均值/亲中值;
优势率(H)=(后代平均值-亲中值)/亲中值[27]。
2 结果与分析
2.1 群体的相似性系数
根据SSR 分子标记鉴定结果,从每个组合随机筛选真杂种66个,利用Jaccard 相似性系数分析2个群体后代之间的亲缘关系,结果表明,正交组合66个后代间的相似性系数为0.227~0.955,平均为0.618;反交组合66个后代间的相似性系数为0.206~0.941,平均为0.593。可见,正交、反交F1群体间的相似性系数无论是其范围还是平均值均存在一定的差异。
根据Jaccard 遗传相似性系数对正交组合后代及其亲本作UPGMA 聚类图(图1)。由图1 可见,在相似性系数为0.578 处,P1和P2明显被分开;在相似性系数0.655 处,P1所在的组又分为5个小组,包括只有5个后代的A1 和有7个后代的A2 和A5,及后代最多的A4 和后代较多的A3(A4 和A3 分别含有19 和17个后代);P2所在的组分为B1 和B2共2个小组,分别包含有7个和6个后代。结果表明,66个后代中有54个跟母本聚类在一起,表现出母性效应,即细胞质效应明显。
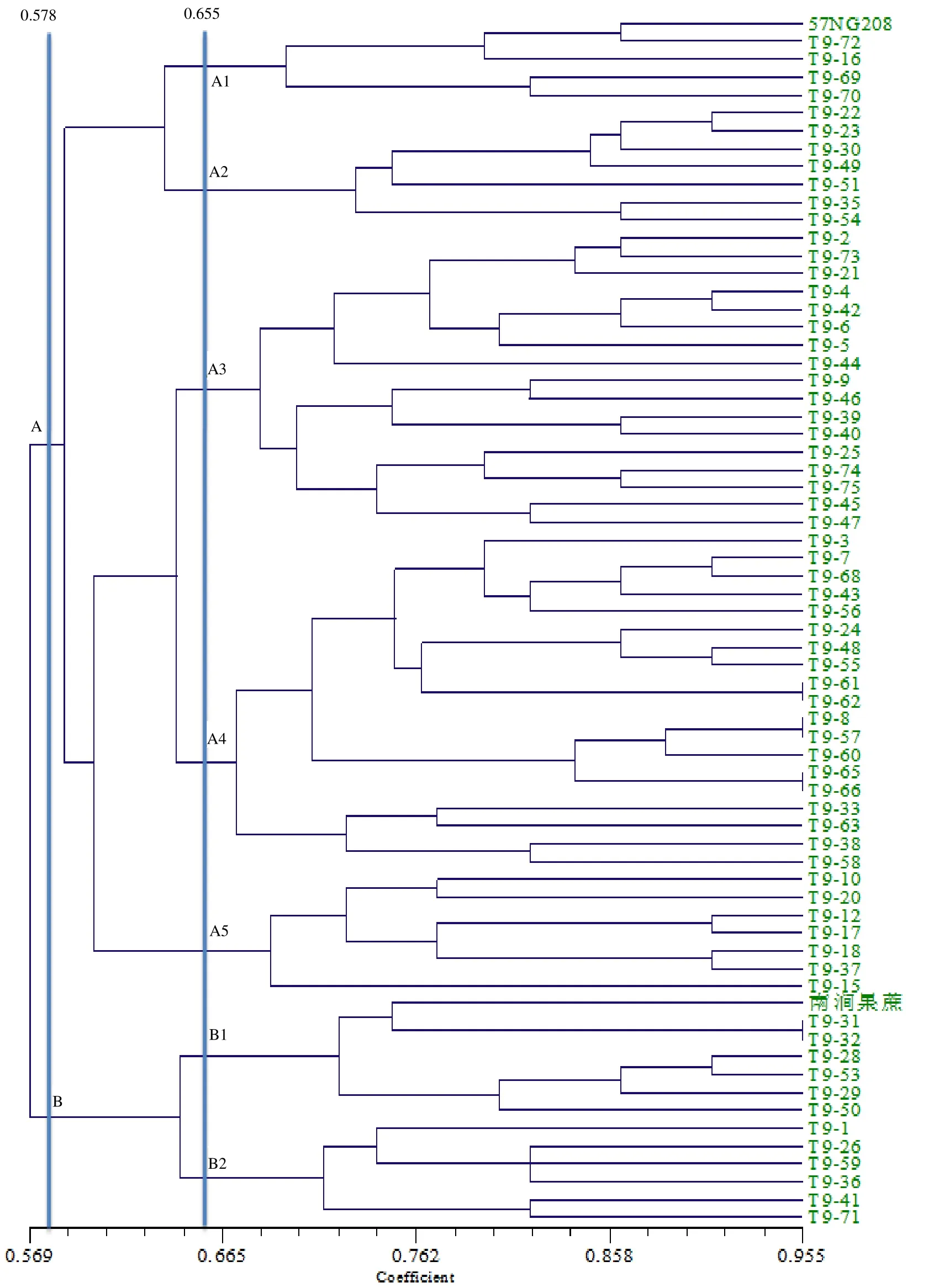
图1 根据遗传相似性系数构建的正交组合UPGMA 聚类图 Fig. 1 UPGMA dendrogram of orthogonal combination according to theirgenetic coefficient
由图2 可见,在相似性系数0.497 处,双亲及66个后代被分为2组,P2为独立一组,其余聚在一类;在相似性系数0.599 处,亲本P1所在的组可分为6 小组,其中包含有28个后代的B3(B3 含有的后代最多);在相似系数0.941 处,编号为2、4、15、37 后代的表现非常相似,彼此聚类在一起。B1 和B4组均含有后代14个;B5、B2 和B6 含有的后代数分别为6、3、2个。
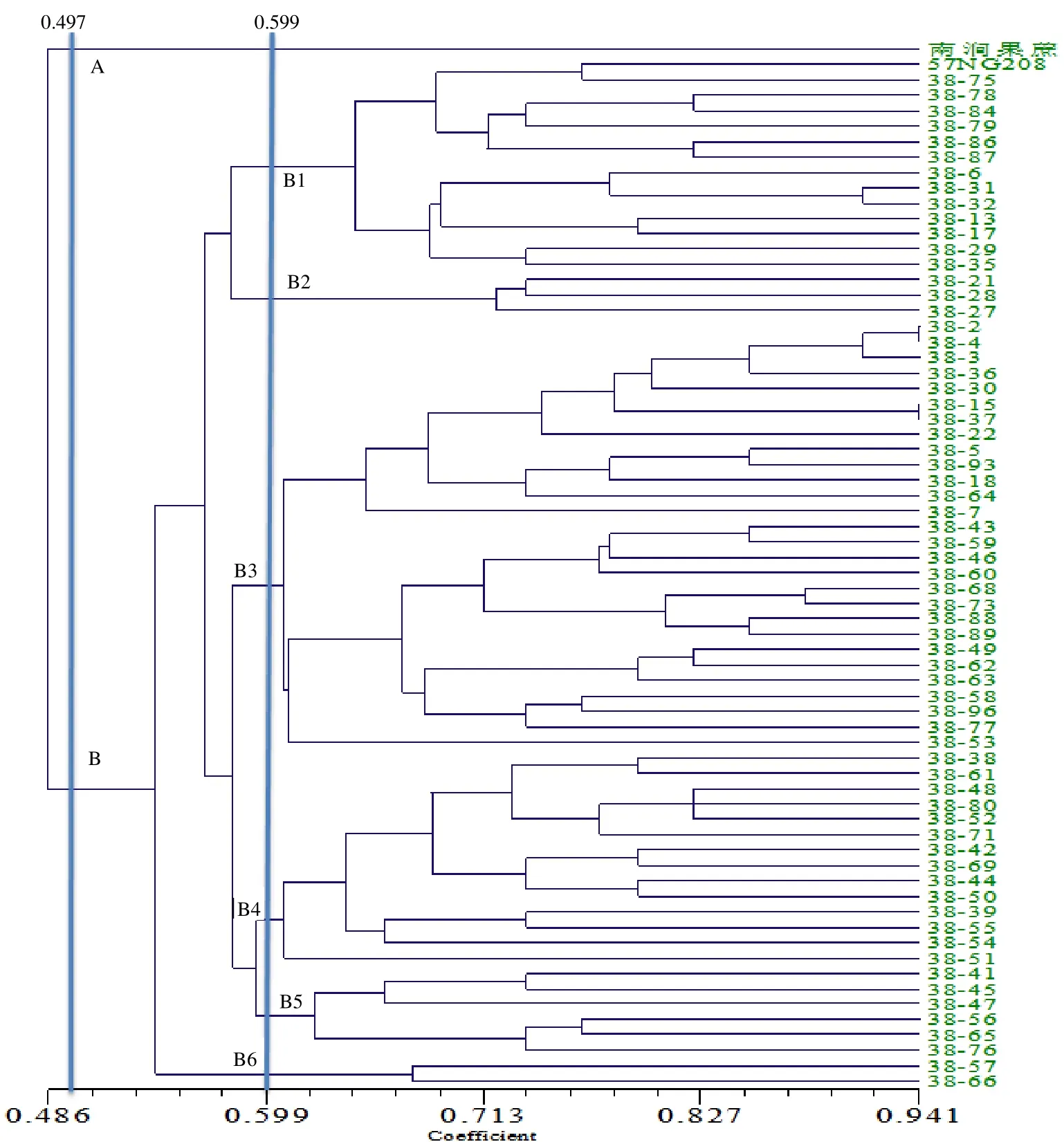
图2 根据遗传相似性系数构建的反交组合UPGMA 聚类图 Fig.2 UPGMA dendrogram of reciprocal combination according to theirgenetic coefficient
2.2 正反交后代株高的遗传变异
甘蔗有性世代的生长期一般是5—6月开始,12月至竖年1月结束,生长时间为半年,因此,有性世代的株高只能反映其生长趋势。株高与倒伏具有一定的正相关性,在其他条件一定的情况下,植株越高,甘蔗倒伏的可能性越大。本试验结果表明,株高为受多基因控制的数量性状遗传(表1,图3、图4)。2个群体后代的遗传传递力分别为82.45%和81.30%,平均株高都低于亲中值(233 cm),也低于低亲植(198 cm)。正交组合后代低于低亲的比率达62.12%,介于双亲间的比率为37.88%,反交组合后代相应值分别为67.67%和32.33%,2组合没有出现超高亲后代,平均优势率为-18.13%,这是利用大茎野生种改良甘蔗品种抗倒伏性状的遗传基础。

表1 正反交后代株高、茎径、丛有效茎数、锤度和单茎重及其变异系数 Table 1 Hereditary variation of plant height, stem diameter, millable stalks per pool , brix and single stem weight on the reciprocal crossesgeneration

图3 57NG208×南涧果蔗后代株高的频率分布 Fig.3 Frequency distribution of height on progenies of 57NG208 × fruit sugarcane of Nanjian
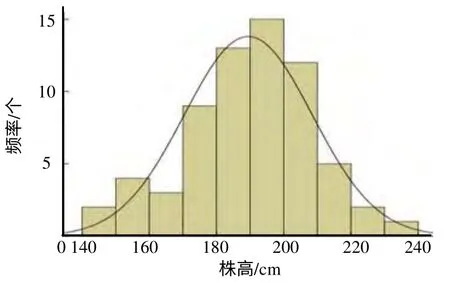
图4 南涧果蔗×57NG208 后代株高的频率分布 Fig.4 Frequency distribution of height on progenies of fruit sugarcane of Nanjian× 57NG208
2.3 正反交后代农艺性状均为数量遗传
试验结果表明,正、反交组合后代株高为受多基因控制的数量性状遗传(图3 和图4)。正、反交组合株高分离极值分别为126~233 cm 和140~234 cm,正、反交后代株高变异系数分别为9.79%和10.07%,说明株高的离散程度不大。茎径、丛有效茎、单茎重和锤度的分布频率与株高分布频率相似,均为正态分布,表现为数量性状遗传(茎径、丛有效茎、单茎重和锤度的频率分布图略)。
2.4 正反交后代茎径的遗传变异
正、反交后代群体茎径的平均值(1.82 cm)均小于亲中值(2.35 cm),变异系数不大,分别为14.12%和15.49%,遗传传递力分别为80.30%和74.21%。正交组合后代介于双亲间的比率(48.48%)与低于低亲的比率(51.52%)相近,而反交组合的相应值分别为33.33%和66.67%,两者的比值接近1∶2。2组合的超高亲值均为0;正交、反交组合后代分离极值分别为1.33~2.43 cm 和1.00~2.42 cm,极值差分别为1.10 和1.42,组合间茎径相差不大。
2.5 正反交后代锤度的遗传变异
正交、反交组合的平均锤度(18.3%)均高于亲中值(15.2%),正交组合的平均锤度甚至高于高亲值。正交、反交组合的遗传传递力都超过了100%,分别为130.09%和110.50%,优势率分别为30.09%和10.50%,这说明锤度的遗传主要是受基因间的加性效应控制,受非加性效应的影响较小。正交、反交组合的变异系数不大,分别为7.2%和10.76%,说明锤度性状的离散度小,在分离群体中表现出相对的稳定性,受环境的影响较小。
正交后代锤度的超高亲比率(86.36%)较高,介于双亲间的比率为13.64%,而反交组合相应的值分别为24.24%和75.76%,2组合低于低亲的比率均为零,说明正交组合宜将高锤度遗传给后代。正交组合锤度的分离极值为16.3%~23.0%,反交组合的为13.3%~21.5%,极值差分别为6.7 和8.2,正交组合中出现高锤度(达23%)的单株有2 株。
2.6 正反交后代丛有效茎数的遗传变异
正交、反交后代在丛有效茎数上有超高的遗传传递力,分别为140.69%和144.16%,优势率分别为40.69%和44.16%,说明丛有效茎数受基因加性效应影响明显。正交、反交组合的变异系数较大,均大于50%,分别为51.41%和51.85%,这说明2个群体后代的分离度比较大,选择后代丛有效茎数突出个体的机会也较大。2个组合后代的表现介于双亲间的较多,均超过了1/2,正交组合的略高;正交组合的超高亲比率为34.84%,反交组合的超高亲比率为37.88%,高于正交组合;正、反交组合低于低亲的比率分别为4.55%和6.06%。
2.7 正反交后代单茎重的遗传变异
由表1 中可见,正交、反交群体的平均单茎重为0.427 kg,均小于亲中值0.657 kg,但高于低亲值0.329 kg;正交、反交群体的遗传传递力分别为69.43%和60.60%,均超过60%;变异系数分别为33.08%和38.63%,反交群体的变异系数比正交群体的大。正交群体的分离极值为0.185~0.915 cm,反交群体的为0.145~0.905 cm,极值差分别为0.730和0.760。单茎重超双亲的均为零,主要是介于双亲间的较多,正交介于双亲间的比率(81.82%)与低于低亲比率(18.18%)的比值接近4∶1,而反交群体两者的比值接近2∶1(65.15%∶34.85%),说明正交组合倾向于偏重遗传,反交组合倾向于偏轻遗传。
3 结论与讨论
以57NG208 与南涧果蔗正反交群体中的132个真实性后代为研究材料,正交、反交F1群体间相似性系数无论是范围还是平均值均存在一定差异。2个群体的UPGMA 聚类图显示,2个亲本均被分在不同组,正交组合中,与母本57NG208 聚在一起的后代有54个;反交组合中,母本南涧果蔗单独聚为一类,66个后代均与父本57NG208 聚在一起。无论是遗传距离,还是与母本聚在一起的后代数量,均存在明显差异,同时遗传相似系数和田间农艺性状亦表现出明显不同,这说明不同母本作为细胞质源对后代的确存在一定的影响。
本试验中农艺性状株高、茎径、锤度、丛有效茎数和单茎重均呈连续分布,属于数量性状遗传,易受环境影响。正交组合有2 株单株表现高糖,反交组合一个后代有16 条丛有效茎。2个组合变异系数排在前3 的依次是丛有效茎数、单茎重和茎径,株高和锤度的变异系数最低,正交组合株高和锤度的变异系数均低于10%,而反交组合这2个性状的变异系数略高于10%。这与前人研究结果一致[28–29]。除了丛有效茎数外,正交组合单茎重、茎径、株高和锤度的平均值均高于反交组合的平均值。
目前生产上所用甘蔗的亲缘关系相近,遗传基础狭窄。本试验中创制新的细胞质源均是在真实性鉴定基础上进行分析研究的,农艺性状调查仅是新植表现,调查过程中并未发现倒伏现象,而且后期观察叶片的持绿程度明显好于同期其他甘蔗材料。宿根的相关性状有待继续跟踪调查。
[1] Arceneau G.Cultivated sugarcane of the world and their botanical derivation[J].Proc Int Soc Sugar Cane Techno,1965,12:844–854.
[2] Sreenivasan T V,Ahhowwalia B S,Heinz D J.Sugarcane Improvement Through Breeding[M].New York:Elsevier Press,1987:211–254.
[3] 刘新龙,马丽,陈学宽,等.利用系谱图和 SSR 标记分析云南自育优良甘蔗品种的遗传关系[J].湖南农业大学学报:自然科学版,2010,36(6):613–619.
[4] 邓海华,符成,李奇伟,等.斑茅F1杂交可育亲本选择及其回交后代鉴定和主要经济性状[J].热带作物学报,2004,25(4):97–101.
[5] 陆鑫,毛钧,刘洪博,等.甘蔗野生种滇蔗茅种质创新利用研究Ⅰ.甘蔗与滇蔗茅远缘杂交F1群体构建与SSR 分子标记鉴定[J].植物遗传资源学报,2012,13(3):321–324.
[6] 陆鑫,刘新龙,毛钧,等.甘蔗野生种滇蔗茅种质创新利用研究Ⅲ.滇蔗茅杂种F1群体的表型变异与遗传多样性分析[J].植物遗传资源学报,2013,14(4):749–753.
[7] 陆鑫,苏火生,林秀琴,等.甘蔗野生种滇蔗茅种质创新利用研究Ⅱ.滇蔗茅杂种F1群体重要农艺性状的遗传分析[J].湖南农业大学学报:自然科学版,2012,38(2):121–124.
[8] 娄红波,王先宏,何丽莲,等.8 份蔗茅与甘蔗杂种F1材料的ISSR 多态性分析[J].西南农业学报,2010,23(5):1409–1412.
[9] 桃联安,经艳芬,董立花,等.云南甘蔗细茎野生种82–114 测交后代主要性状遗传分析[J].植物遗传资源
学报,2011,12(3):419–424.
[10] 陆鑫,蔡青,王丽萍,等.大茎野生种57NG208 杂种后代综合评价[J].中国糖料, 2008(3):15–17.
[11] 李杨瑞.现代甘蔗学[M].北京:中国农业出版社,2012:106–116.
[12] 王文明,周开达,文宏灿,等.胞质效应在杂交水稻主要数量性状上的多样性[J].中国水稻科学,1997,11(2):65–69.
[13] 蔡玉红,刑少辰.籼型杂交稻不同细胞质对稻瘟病抗性差异的研究[J].邯郸农业高等专科学校学报,1995 (3):24–26.
[14] 姬广海,许志刚.水稻品种对细菌性条斑病的抗性研究[J].西南农业大学学报,2001,23(2):164–166.
[15] 侯宁,刘春光,刘根齐,等.异源细胞质小麦的赤霉病抗性研究[J].麦类作物学报,2003,23(1):7–11.
[16] 易自力,徐乃瑜.不同山羊草细胞质效应的比较研究[J].草业学报,2000,9(1):73–78.
[17] 徐乃瑜,易自力,何瑞锋.小麦异源细胞质对主要病害抗性影响的初步研究[J].作物学报,1991,17(5):326–339.
[18] 刘光杰,占小登,沈君辉,等.水稻同核异质不育系材料对白背飞虱抗性的研究初报[J].中国水稻科学,2003,17(1):89–90.
[19] 吴郁文,张翠兰,刘春光,等.异源细胞质小麦育种技术[J].中国科学:C 辑,1998,28(1):57–64.
[20] 邹剑秋,王艳秋,张飞,等.A1、A3 型细胞质甜高粱品种抗倒性能研究[J].中国农业大学学报,2012,17(6):92–97.
[21] 易自力,徐乃瑜.小麦异源细胞质遗传效应的比较研究[J].应用与环境生物学报,2000,6(4):307–312.
[22] 刘新龙,蔡青,毕艳,等.中国滇蔗茅种质资源遗传多样性的AFLP 分析[J].作物学报,2009,35(2):262–269.
[23] 刘洪博,应雄美,毛钧,等.11 份割手密遗传多样性的SSR 分析[J].植物遗传资源学报,2013,14(3):189–193.
[24] Aitken K S,Jackon P A,McIntyre C L.A combination of AFLP and SSR markers provides extensive map coverage and identification of homo(eo) logous linkagegroups in a sugarcane cultivar[J].Theor Appl Genet,2005,110:789–801.
[25] 刘新龙,蔡青,毕艳,等.甘蔗AFLP 标记和SSR 标记的PAGE 胶快速银染检测方法[J].江苏农业学报,2009,25(2):433–435.
[26] 刘新龙,蔡青,吴才文,等.甘蔗品种资源的表型遗传多样性研究[J].生物多样性,2010,18(1):37–43.
[27] 崔艳波,陈慧,乐文全,等.‘京白梨’与‘鸭梨’正反交后代果实性状遗传倾向研究[J].园艺学报,2011,38(2):215–224.
[28] 黄文华.甘蔗杂交后代遗传变异分析与标记检测群体构建[D].福建农林大学,2005:23–28.
[29] 阙友雄,黄文华,许莉萍,等.甘蔗杂交后代遗传变异评价及高糖和低糖池构建[J].热带作物学报,2009,30(6):811–816.
