中国近海入侵贝类及其影响
2015-07-01宋积文董艳红李海涛
宋积文, 董艳红, 李海涛, 赵 研
1中国海洋石油总公司,北京 100094; 2国家海洋局南海环境监测中心,广东 广州 510300;3中国科学院生态环境研究中心,北京 100085
中国近海入侵贝类及其影响
宋积文1, 董艳红2, 李海涛2, 赵 研3*
1中国海洋石油总公司,北京 100094;2国家海洋局南海环境监测中心,广东 广州 510300;3中国科学院生态环境研究中心,北京 100085
海洋贝类种类繁多,是近海底栖生态系统的优势种,在生态系统物质循环和能量流动中起着极其重要的作用;但因频繁的人类活动导致的贝类入侵问题,往往使该过程不能正常运转。大量研究表明,入侵贝类会抑制原著贝类或其他生物的生长及拓殖,形成单一优势群落,破坏当地生态系统的平衡,也会对海洋渔业生产和近海工程及作业等造成不可预测的危害。目前,欧美等国家针对该问题已开展了大量工作,十分重视入侵贝类的生物学、生态学与行为学特征及特性的研究,并根据其入侵途径与方式的不同开展了检测、监测、防控及管理工作。但在我国,入侵贝类还未引起相关部门及研究机构的足够重视。本文主要简述了我国近海的3种入侵贝类沙筛贝、指甲履螺和地中海贻贝的生物学、生态学及行为学等特征及其对当地生态系统的影响;同时,根据其特性及入侵过程控制的难易,建议在完善我国入侵贝类数据库的基础上加强早期预警与检测,以及安全防控和管理等工作。
入侵生物; 贝类; 生物多样性; 沙筛贝; 指甲履螺; 地中海贻贝
追溯华莱士线(Wallace′s Line) 下不同动、植物的变迁历史就会发现生物入侵过程一直存在,不同地质时期的板块运动、气候变化等都会将外来物种带入新生境。资料显示,陆地、河流等生态系统受到的影响较大,而海洋生态受到的影响较少(Elton,1958; Herderetal.,2012)。但伴随着全球化进程的加快,日益增多的航运、水产养殖等活动把经过亿万年长期隔离的物种带到世界各个海域,使生物入侵频发(Lockwoodetal.,2013)。许多研究表明,一个物种被有意或无意引进一个新的生境中完成建群及拓殖成为入侵生物的过程往往伴随当地生态过程、生态系统结构的急剧变化,且会危及当地的生物多样性甚至导致当地物种灭绝(Donlan,2010; Royetal.,2014)。目前,由于生物入侵导致各种海洋生态系统生物多样性锐减的事件不断发生,有超过84%的海洋生态区(Ecoregion)遭受入侵生物的威胁(Baxetal.,2003; Molnaretal.,2008)。
早在19世纪,就有研究表明,太平洋海域因生物入侵发生过一系列生态学变化,其中,夏威夷群岛、北美、澳大利亚及亚洲沿海等区域是主要的生物入侵地(Carlton,1987)。大部分外来生物通过包括航运、经济贝类和鱼类贸易等人类活动的介导到达新的区域,如贻贝、藤壶、水螅、海绵、藻类等是可附着在船只上的污损生物。虽然在诸多人类介导的物种传播事件中,只有少数海洋生物可成功入侵到新的生态系统,成功定殖进而成为入侵生物;但是其危害极其显著,一般包括:取代原有物种,改变当地群落结构,并通过食物链改变当地基本的生态学过程;损害海水养殖业、腐蚀船只、堵塞进水管道等,给当地经济带来很大的损失。此外,一些入侵物种能够携带致病菌,对其他生物甚至人类构成直接威胁(Diaz & Rosenberg,2008; Molnaretal.,2008; Perringsetal.,2002; Pimenteletal.,2005; Ruizetal.,1997)。
北美、欧洲及澳大利亚等国家和地区对海洋生物入侵问题关注较早,对一些主要海洋入侵生物的入侵途径、建群机制及生态危害做了较为详细的研究(Carlton,1987; Freeman & Byers,2006; Lockwoodetal.,2010; McQuaid & Phillips,2000; Paoluccietal.,2014; Willan,1985)。但相比而言,我国对海洋生态系统生物入侵的研究则较少(李小慧等,2009; 万方浩等,2012; Caietal.,2014; Pimenteletal.,2005; Weberetal.,2008)。
我国海岸线长、港口众多,沿海人口密度大、人类活动较为频繁,为入侵生物的引进、定殖、归化等提供了非常有利的条件。因此,目前亟需了解已入侵或及有可能入侵我国近海的外来物种,并开展这些入侵物种预警、防控、消除等研究工作。本文拟分析我国近海入侵贝类及其对近海环境的影响,并提出近海贝类生物入侵的防控措施及注意事项。
1 我国近海的入侵贝类
生物入侵主要包括3种渠道:(1)自然入侵,是非人为因素引进,主要通过风媒、水流或昆虫、鸟类等媒介发生自然迁移;(2)无意引进,伴随着进出口贸易、船舶航运或入境旅游等人类活动而被无意引入;(3)有意引进,主要指各国出于发展农业、林业、渔业、观赏等人为需求,有意识引进优良动、植物品种等。海洋入侵生物也通过上述3种主要渠道到达世界各地(Lockwoodetal.,2013)。目前,我国已确认的海洋入侵生物共计约43种,主要为无脊椎动物(25种),其中72%的无脊椎动物均由船舶船体或压舱水等航运介导而无意带入(表1),且对当地环境造成了一定危害(黄宗国,1984; 梁玉波和王斌,2001; 陆琴燕等,2013; 徐海根等,2004; Xuetal.,2006)。在上述无意引进的物种中,入侵贝类有3种,且已对沿海的生物多样性、水产养殖及当地生态系统等造成破坏,需要重点关注。
1.1 沙筛贝Mytilopsissallei及其危害
沙筛贝俗称黑条纹贻贝,成体壳长8~25 mm,颜色从黑色到浅色变化较大,左右壳重叠、右壳稍大,一般栖息于潮间带和浅海区域。该类生物雌雄异体,繁殖力极强,且对温度、盐度和溶解氧等有较宽的适应范围和耐受性。其可形成大量群体且固着在一起,是一种生存模式符合r策略的海水入侵生物(GISD, Global Invasive Species Database, http:∥www.issg.org/database/welcome/)。沙筛贝原产于中美洲热带水域,最早可能是通过巴拿马运河入侵到斐济岛,在被引进印度之前一直没有相关报道。19世纪七八十年代,该物种又相继被引进日本和我国台湾;之后随船舶进入维多利亚港,并迅速大量繁殖(Morton,1980、1989)。目前,其已扩散至南部大部分沿海地区,成为近海主要的入侵生物。
虽有报道称沙筛贝滤水量大,对澄清水质有一定贡献,但由于其增长快速,会加速环境氧耗,形成死亡区(Dead zones),不利于其他生物生存,导致原有生物群落的衰退或死亡,进而造成所处水域严重的有机污染;同时,其繁殖力强,会掠夺食物、空间资源,所到之处迅速形成单一优势群落,排挤原生境中的大部分生物,使当地生物多样性严重下降,损害当地生态系统(Caietal.,2014; Wongetal.,2011)。在福建一些海湾,沙筛贝大量繁殖,占据了海面上的浮筏、桩柱等养殖设施,造成当地养殖的菲律宾蛤仔Ruditapesphilippinarum、翡翠贻贝Pernaviridis和牡蛎Crassostreaspp.等经济贝类产量大幅度下降,甚至使原有数量很大的藤壶等本土底栖生物大量减少甚至绝迹(王珊等,2011)。另外,其常附着在船舶、码头、石油平台及其他生产活动设施上,对其污损或腐蚀,严重影响了当地经济和人民生活(Wongetal.,2011)。

表1 经压舱水或航运途径入侵中国的海洋无脊椎动物Table 1 Invasive marine invertebrates transported to China via ballast water
*指海洋入侵贝类。
*are marine shellfish.
1.2 指甲履螺Crepidulaonyx及其危害
指甲履螺,属滤食性腹足类。外壳腹面有一块横隔板,板的前缘有中央白色小凹痕,背面呈椭圆形或指甲形。身体呈深褐色,成体壳长25~40 mm。除较小幼体会移动外,雌性和大个体都终生不动。但由于其对盐度、温度及溶解氧有较宽的适应幅,浮游期幼体很容易在一个地方固定并迅速繁殖(Gomoiuetal.,2002; Li & Chiu,2013)。其繁殖策略较为独特:成年个体会叠加在一起形成一座塔,相互附着形成一个交配链;底部通常为雌性,如果底部雌性消失,它们会用独特的性别调控模式调控上面的雄性变为雌性,让这个链条得以维系,因此它们能保持较高的繁殖率且很快能达到很高的种群密度(Coe,1953)。该类生物原产于中美洲波多黎各到美国加利福尼亚等地,于19世纪70年代末期随航运船只抵达我国香港并在维多利亚港快速完成高密度建群,继而迅速扩散到深圳湾,成为我国近海的主要入侵贝类(黄宗国,1984; Woodruffetal.,1986)。
与原产于北美沿岸且入侵英国沿海的近亲大西洋履螺Crepidulafornicate类似,指甲履螺会滤食各种浮游生物,并与当地贝类争夺生存空间,由于其繁殖迅速很快会成为入侵地的优势群落,严重降低当地生物多样性(Elton,1958)。同时,它们喜欢附着在贻贝、牡蛎的壳上,影响经济贝类的正常育种和繁殖,使养殖产业受到损失。另外,其喜欢附着在水泥柱、铁管、网箱等上,损害这些公共设施,是我国近海特别是南海区域主要的优势污损生物(黄宗国,1984)。
1.3 地中海贻贝Mytilusgalloprovincialis及其危害
地中海贻贝,是紫贻贝复合种M.edulisspecies complex 3个代表种之一,也称蓝贻贝。其属于双壳类,两片壳接近四边形,颜色呈深蓝色、褐色到几乎黑色,体长一般为5~8 cm,最长可达 15 cm。有关地中海贻贝的入侵机制尚不明确,但由于其繁殖迅速,在一些地区会比原著物种有生长优势(繁殖速率为本土物种的1.2或3.0倍),并能很快取代原有物种;且地中海贻贝可耐受7 d持续暴晒并依然保持较高的存活率,具有入侵生物繁殖力强、适应性强的共性(Branch & Steffani,2004)。该贝类原产于地中海海岸、黑海与亚得里亚海,在19世纪80年代被引进欧洲,随后通过航运或养殖等途径被引种到世界各地,并在20世纪初期成为入侵日本、韩国及俄国沿海的主要物种。目前,该贝类已成功在全球范围扩散并建立族群,几乎出现在所有温带地区与大港口附近(Carlton,1996)。在我国,地中海贻贝主要分布于黄海、渤海海区,经人工移养后在东海和南海也能生长发育。
有关地中海贻贝生态危害的报道较多。如在非洲南部海岸,该物种成为西海岸岩石潮间带的优势种,并且使入侵区域与非入侵区的生物地貌表现出明显差异(Robinson & Griffiths,2002);在希腊,由于其大量繁殖,体外胞质呼吸酸过量释放导致海湾水体酸化等(Michaelidisetal.,2005)。因此,该物种被列入世界百大外来入侵种名单(GISD, Global Invasive Species Database, http:∥www.issg.org/database/welcome/)。但在我国,其主要作为经济物种被培育,有一定的经济价值且还未造成大面积的生态危害,因此其对本土族群与原生贻贝的影响常被忽略(Branch & Steffani,2004; Geller,1999)。但该物种存在复合种且形态鉴定困难,会增加错误引进的风险,因此仍需要特别注意。另外,地中海贻贝对多种污染物有较强的生物富集能力,且能携带各种病原生物和污染物,因此作为食用贝类仍有很大的健康风险(颜天等,2001)。
综上,3种入侵贝类均具有增长迅速、对生境适应性强等生物特性,且它们殖入新的栖息地后缺少相应天敌,一般都能很快形成单种优势群落(Astudilloetal.,2014),因而会打破当地物种相互间的动态平衡,致使物种种群数目锐减并降低入侵地的生物多样性,进而改变或破坏原本的生态系统和生态功能(Curnutt,2000; Traveset & Richardson,2014),甚至会改变原有的生物地理分布和自然生态系统的结构与功能,对入侵地的生物地貌系统造成一定的影响(Feietal.,2014)。此外,上述3种贝类也是主要的污损优势种类,严重污损、腐蚀近海的基础设施及养殖网箱设备等,且它们往往携带很多病毒或病原微生物,对人们的生活和健康存在一定威胁。
2 入侵贝类的预防、管理与控制
海洋入侵生物的防控和管理工作已在很多国家得以开展,并取得一定成效。入侵贝类的预防和控制可以借鉴其他国家的成功经验,在入侵初期建立早期预警监测技术,并对各个阶段的入侵贝类进行综合治理,建立完善入侵贝类的档案库,预防具有潜在入侵风险贝类的引进,培养公众的科普和权利意识、消除有意引入途径带来的风险。
2.1 完善入侵贝类早期监测与预警技术
生物入侵的基本过程一般分为5个阶段(图1),一旦入侵生物成为入侵地的优势种并普遍存在之后就很难再对其进行处理和控制(Colautti & MacIsaac,2004)。由于多数入侵贝类具有繁殖速度快、适应范围广、耐受力强且喜欢附着船舶载体等生物特性,其很容易通过日益频繁的航运途径完成第Ⅰ和第Ⅱ阶段抵达新栖息地,且通过环境过滤后以指数级的速率大量繁殖和扩散。因此,为减少贝类生物入侵带来的危害,必须在其数量很少时进行控制,即在第Ⅲ阶段入侵贝类生物建群完成之前做好预防、检测、控制和消除工作。
压舱水是外来贝类物种入侵的主要途径之一,每年穿行于各大洋的船舶在世界各地排放大量压舱水,其中夹带着数不清的外来物种,这些外来物种有的已在当地环境中泛滥成灾(王珊等,2011; 徐海根等,2004; Aniletal.,2002; Lockwoodetal.,2013)。尽管我国对船底和压舱水携带海洋外来物种的预警监测和评估做了一些工作,但这一体系仍待进一步健全和完善。另外,因很多入侵贝类生物被引入初期并没有可观的群体数量,不容易被传统技术监测,因而得不到及时控制,当大家开始关注时,其一般已完成拓殖和建群过程,因此建立灵敏的早期监测与预警的核心技术非常必要。
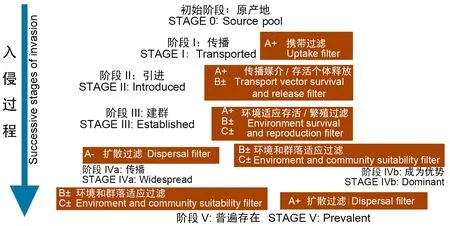
图1 生物入侵过程简略图(改自Colautti & MacIsaac,2004)Fig.1 Suggested framework of invasion process (revised from Colautti & MacIsaac,2004) A.繁殖体压力;B.生理化学因子;C.群落相互作用。 A. Propagule pressure; B. Physico-chemical factors; C. Community interaction.
随着分子技术的不断发展,高通量测序平台为水生入侵生物早期监测与预警核心技术的建立奠定了基础。目前,已有通过水生生物特定引物设计及序列扩增、基于高通量测序平台、针对水环境中极其稀有的特定物种的检测技术(Zhanetal.,2013)。应用该技术可以精确分析特定海域或压舱水是否存在特定入侵贝类生物,并用基因定量PCR技术对其数量和入侵风险进行评估,把入侵生物的危害遏制在萌芽状态。
2.2 建立合理规范的管理体系
生物入侵不能简单地靠技术检测和控制,还必须依靠法律约束和条例规范。目前,针对外来生物入侵的最有影响力的国际法律文件是1992 年的《生物多样性公约》。之后各个国家及组织机构也相继制定了生物入侵管理控制的相应对策,如环境科学问题委员会(SCOPE)、IUCN 和联合国环境规划署在1997 年共同发起的《全球入侵物种计划》;美国在1996 年颁布的“国家外来物种法”及在1999 年签署的总统令都强化了美国政府对入侵动、植物的研究和管理力度;澳大利亚在1996年制定了《澳大利亚生物多样性保护国家策略》并成立了国家生物安全局,强调并重视外来物种的管理问题;俄罗斯于2001 年创建了生物入侵研究与管理中心(闫小玲等,2012)。虽然我国于2000年发布了《全国生态环境保护纲要》,明确提出了“维护国家生态环境安全”的目标,但没有专门的国家级机构或部门针对我国外来物种建立生物信息库,制定控制与预防的计划和策略,健全检测、检疫制度并对不同入侵生物的入侵机理设立专项研究基金等(万方浩等,2002)。
目前,有关植物入侵问题已引起重视,但针对海洋入侵生物的管理和控制还较为薄弱。因此,必须制定相应的规章制度和建立特殊机构对海洋入侵生物进行检测、控制和管理,以将其危害控制在最小范围内。
2.3 完善入侵贝类生物档案库
目前,全球入侵物种数据库(Global Invasive Species Database, GISD)已收录入侵物种891种,其中已在世界范围内不同地区发现的入侵贝类有32种(包括淡水种和海水种,http:∥www.issg.org/database/welcome/)。GISD的建立在一定程度上为人们了解全球入侵物种的分布状况提供了一个平台,但仍然缺乏足够的数据描述全球入侵物种的具体分布图;我国对入侵生物的研究起步较晚,缺乏详细调查,对大多外来入侵种的本底资料还很不清楚;同时没有专门针对上述入侵贝类进行生态评估、安全评估(贝类毒素涉及的食用及疾病传播)及经济评估的深入研究。因此,需要在国外入侵贝类数据库的基础上,依据国际标准建立评估等级,对已经产生危害或有潜在危害的贝类进行等级划分和管理控制,建立针对我国本土的入侵贝类数据库。
2.4 宣传教育和公众参与
生物入侵并非仅仅是一种自然现象,人为因素的影响远远超越了原本的自然过程,在世界范围内产生严重危害的入侵事件均是通过人类活动介导的(Lockwoodetal.,2013; http:∥www.issg.org/database/welcome/)。我国对人为介导的生物入侵所造成的生态和经济危害还没有足够重视,其中由于缺乏有效的科学知识与信息,缺乏对引进物种的利益与风险的正确评估,淡薄的生态意识与不顾生态后果的经济利益驱使,无法可依或有法不依与执法不严等因素造成的生物入侵事件频繁发生(万方浩等,2002)。因此,必须大力培养公众对入侵生物危害的正确认识,并加强其责任意识,监督和遏制盲目引进或利益驱动下的物种引进行为。另外,还需要号召国内各检疫或海关部门与科研单位或国外机构密切合作,对存在入侵风险的贝类做到心中有数并禁止入境,杜绝存在潜在风险的外来生物进入我国境内。
3 小结
生物多样性是海洋生态系统正常维系和运转的基础,贝类入侵导致的生物多样性锐减及生态结构单一等会打破这种生态平衡,进而损害当地生态系统。在补充和完善入侵贝类档案的同时,可以对有意引进的养殖贝类进行风险评估,并对无意引进的入侵贝类进行早期预警与检测。总之,随着人们对入侵生物危害意识的加强及相应法律法规的制定和完善,由有意引进导致的贝类入侵现象将逐渐减少;各科研机构及国际合作交流的加强对于我国近海入侵贝类防控和治理也具有重要意义。
黄宗国. 1984. 指甲履螺扩散到深圳湾. 海洋通报, 6(3): 91-92.李小慧, 胡隐昌, 宋红梅, 王培欣, 汪学杰, 牟希东, 刘超, 罗建仁. 2009. 中国福寿螺的入侵现状及防治方法研究进展. 中国农学通报, 25(14): 229-232.
梁玉波, 王斌. 2001. 中国外来海洋生物及其影响. 生物多样性, 9(4): 458-465.
陆琴燕, 刘永, 李纯厚, 魏小岚, 刘勇. 2013. 海洋外来物种入侵对南海生态系统的影响及防控对策. 生态学杂志, 32(8): 2186-2193.
万方浩, 郭建英, 王德辉. 2002. 中国外来入侵生物的危害与管理对策. 生物多样性, 10(1): 119-125.
万方浩, 刘全儒, 谢明. 2012. 生物入侵: 中国外来入侵植物图鉴. 北京: 科学出版社.
王珊, 刘瑀, 王海霞, 刘保占. 2011. 船舶压载水带来的生物入侵及其解决途径. 中国水产, (9): 24-26.
徐海根, 强胜, 韩正敏, 郭建英, 黄宗国, 孙红英, 何舜平,丁晖, 吴海荣, 万方浩. 2004. 中国外来入侵物种的分布与传入路径分析. 生物多样性, 12(6): 626-638.颜天, 傅萌, 李钧, 于仁诚, 周名江. 2001. 麻痹性贝毒PSP在紫贻贝体内的累积、转化与排出. 海洋与湖沼, 32(4): 420-427.闫小玲, 寿海洋, 马金双. 2012. 中国外来入侵植物研究现状及存在的问题. 植物分类与资源学报, 34(3): 287-313.Anil A, Venkat K, Sawant S, Dileepkumar M, Dhargalkar V, Ramaiah N, Harkantra S N and Ansari Z A. 2002. Marine bioinvasion: concern for ecology and shipping.CurrentScience, 83: 214-219.
Astudillo J C, Wong J C, Dumont C P, Bonebrake T C and Leung K M. 2014. Status of six non-native marine species in the coastal environment of Hong Kong, 30 years after their first record.BioInvasionsRecords, 3: 123-137.
Bax N, Williamson A, Aguero M, Gonzalez E and Geeves W. 2003. Marine invasive alien species: a threat to global biodiversity.MarinePolicy, 27: 313-323.
Branch G M and Steffani C N. 2004. Can we predict the effects of alien species? A case-history of the invasion of south Africa byMytilusgalloprovincialis(Lamarck).JournalofExperimentalMarineBiologyandEcology, 300: 189-215.Cai L Z, Huang J S, Dahms H U, Fu S J, Zhuo Y and Guo T. 2014. Effect of the invasive bivalve mytilopsis sallei on the macrofaunal fouling community and the environment of yundang lagoon, Xiamen, China.Hydrobiologia, 741: 101-111.Carlton J T. 1987. Patterns of transoceanic marine biological invasions in the Pacific Ocean.BulletinofMarineScience, 41: 452-465.Carlton J T. 1996. Pattern, process, and prediction in marine invasion ecology.BiologicalConservation, 78: 97-106.
Coe W R. 1953. Influences of association, isolation, and nutrition on the sexuality of snails of the genusCrepidula.JournalofExperimentalZoology, 122: 5-19.Colautti R I and MacIsaac H J. 2004. A neutral terminology to define ‘invasive’species.DiversityandDistributions, 10: 135-141.Curnutt J L. 2000. Host-area specific climatic-matching: similarity breeds exotics.BiologicalConservation, 94: 341-351.
Diaz R J and Rosenberg R. 2008. Spreading dead zones and consequences for marine ecosystems.Science, 321: 926-929.
Donlan C J. 2010. Aliens, globalization, and biodiversity.TrendsinEcology&Evolution, 25: 624-625.
Elton C S. 1958. The ecology of invasions by animals and plants. Chicago, USA: University of Chicago Press.
Fei S, Phillips J and Shouse M. 2014. Biogeomorphic impacts of invasive species.AnnualReviewofEcology,Evolution,andSystematics, 45: 69-87.
Freeman A S and Byers J E. 2006. Divergent induced responses to an invasive predator in marine mussel populations.Science, 313: 831-833.
Geller J B. 1999. Decline of a native mussel masked by sibling species invasion.ConservationBiology, 13: 661-664.
Gomoiu M T, Alexandrov B, Shadrin N and Zaitsev Y. 2002.TheBlackSea—ARecipient,DonorandTransitAreaforAlienSpecies. Netherlands: Springer.
Herder F, Schliewen U K, Geiger M F, Hadiaty R K, Gray S M, McKinnon J S, Walter R P and Pfaender J. 2012. Alien invasion in Wallace′s Dreamponds: records of the hybridogenic "flowerhorn" cichlid in Lake Matano, with an annotated checklist of fish species introduced to the Malili Lakes system in Sulawesi.AquaticInvasions, 7: 521-535.Li A and Chiu J. 2013. Latent effects of hypoxia on the gastropod crepidula onyx.MarineEcologyProgressSeries, 480: 145-154.Lockwood B L, Sanders J G and Somero G N. 2010. Transcriptomic responses to heat stress in invasive and native blue mussels (genusMytilus): molecular correlates of invasive success.TheJournalofExperimentalBiology, 213: 3548-3558.Lockwood J L, Hoopes M F and Marchetti M P. 2013.InvasionEcology. Malden, Netherlands: Blackwell Publishing.
McQuaid C D and Phillips T E. 2000. Limited wind-driven dispersal of intertidal mussel larvae: in situ evidence from the plankton and the spread of the invasive speciesMytilusgalloprovincialisin South Africa.MarineEcologyProgressSeries, 201: 211-220.
Michaelidis B, Ouzounis C, Paleras A and Pörtner H O. 2005. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine musselsMytilusgalloprovincialis.MarineEcologyProgressSeries, 293: 109-118.Molnar J L, Gamboa R L, Revenga C and Spalding M D. 2008. Assessing the global threat of invasive species to marine biodiversity.FrontiersinEcologyandtheEnvironment, 6: 485-492.Morton B. 1980.Mytilopsissallei(Recluz) (Bivalvia: Dreissenacea) recorded from Hong Kong: an introduction by Vietnamese refugees.MalacologicalReview, 13: 90-92.
Morton B. 1989. Life-history characteristics and sexual strategy ofMytilopsissallei(Bivalvia: Dreissenacea), introduced into Hong Kong.JournalofZoology, 219: 469-485.
Perrings C, Williamson M, Barbier E B, Delfino D, Dalmazzone S, Shogren J, Simmons P and Watkinson A. 2002. Biological invasion risks and the public good: an economic perspective.ConservationEcology, 6: 1.Pimentel D, Zuniga R and Morrison D. 2005. Update on the environmental and economic costs associated with alien-invasive species in the United States.EcologicalEconomics, 52: 273-288.Robinson T and Griffiths C. 2002. Invasion of langebaan lagoon, south Africa, byMytilusgalloprovincialis— effects on natural communities.AfricanZoology, 37: 151-158.Roy H E, Peyton J, Aldridge D C, Bantock T, Blackburn T M, Britton R, Clark P, Cook E,etal. 2014. Horizon scanning for invasive alien species with the potential to threaten biodiversity in great britain.GlobalChangeBiology, 20: 3859-3871.Ruiz G M, Carlton J T, Grosholz E D and Hines A H. 1997. Global invasions of marine and estuarine habitats by non-indigenous species: mechanisms, extent, and consequences.AmericanZoologist, 37: 621-632.
Traveset A and Richardson D M. 2014. Mutualistic interactions and biological invasions.AnnualReviewofEcology,Evolution,andSystematics, 45: 89-113.
Weber E, Sun S G and Li B. 2008. Invasive alien plants in China: diversity and ecological insights.BiologicalInvasions, 10: 1411-1429.Willan R C. 1985. Successful establishment of the Asian musselMusculistasenhousia(Benson in Cantor, 1842) in New Zealand.RecordsoftheAucklandInstituteandMuseum, 22: 85-96.Wong Y, Meier R and Tan K. 2011. High haplotype variability in established asian populations of the invasive caribbean bivalveMytilopsissallei(Dreissenidae).BiologicalInvasions, 13: 341-348.Woodruff D, McMeekin L, Mulvey M and Carpenter M. 1986. Population genetics of crepidula onyx: variation in a Californian slipper snail recently established in China.TheVeliger, 29: 53-63.Xu H, Ding H, Li M, Qiang S, Guo J, Han Z, Huang Z, Sun H, He S, Wu H and Wan F. 2006. The distribution and economic losses of alien species invasion to China.BiologicalInvasions, 8: 1495-1500.
Zhan A, Hulak M, Sylvester F, Huang X, Adebayo A A, Abbott C L, Adamowicz S J, Heath D D, Cristescu M E and MacIsaac H J. 2013. High sensitivity of 454 pyrosequencing for detection of rare species in aquatic communities.MethodsinEcologyandEvolution, 4: 558-565.
(责任编辑:杨郁霞)
Invasive shellfish and their impacts on Chinese coastal waters
Ji-wen SONG1, Yan-hong DONG2, Hai-tao LI2, Yan ZHAO3*
1ChinaNationalOffshoreOilCorporation,Beijing100094,China;2SouthChinaSeaEnvironmentalMonitoringCenter,StateOceanicAdministration,Guangzhou,Guangdong510300,China;3ResearchCenterforEco-EnvironmentSciences,ChineseAcademyofSciences,Beijing100085,China
As one of the most diverse and abundant taxonomic groups, shellfish plays important roles in energy flow and nutrient cycling in marine ecosystems. However, this process has been badly hampered by invasive shellfish. Invasive shellfish become the dominant species after their establishment, threaten native species and inhibit the growth of other species, erode fishing facilities and artificial structures, leading to ecosystem damage, economic loss and inconvenience to locals. The study of biological invasions by shellfish has
intensive attentions in the marine environment in many developed countries, and researches mainly focused on studying inherent traits of species, invasion pathway, developing technology for early warning and controlling, and improving ability to generate invasion theory and manage invasive species. However, the study of the shellfish invasions has not been well received enough attention in China as well as other aquatic invasion species. This review aims to summarize the main invasive shellfish (includingMytilopsissallei,Crepidulaonyx, andMytilusgalloprovincialis) of Chinese coasts and discuss their damage to local ecosystems based on their special traits or behavior. At the same time, according to the characteristics and the control of the invasion process, it was suggested that we should strengthen the early warning and detection, and the prevention and management.
invasive species; shellfish; biodiversity;Mytilopsissallei;Crepidulaonyx;Mytilusgalloprovincialis
2015-06-08 接受日期(Accepted): 2015-07-08
南海区海洋环境质量综合评估方法[DOMEP(MEA)-01-03]
宋积文, 男, 工程师。 研究方向: 近岸海洋环境变化及诱因。 E-mail: songjw@cnooc.com.cn
*通讯作者(Author for correspondence), E-mail: yanzhao@rcees.ac.cn
10. 3969/j.issn.2095-1787.2015.03.001
