诱导耐药型宋内志贺菌中主动外排阻遏基因acrR、marR的变化
2015-05-06高佩增姚明晓
高佩增,姚明晓
(1.济南市第四人民医院检验科,山东济南250031;2.山东省疾病预防控制中心病原生物研究所)
诱导耐药型宋内志贺菌中主动外排阻遏基因acrR、marR的变化
高佩增1,姚明晓2
(1.济南市第四人民医院检验科,山东济南250031;2.山东省疾病预防控制中心病原生物研究所)
目的分析宋内志贺菌在被环丙沙星诱导耐药后其主动外排阻遏基因acrR、marR突变与表达量的变化。方法 通过1/2MIC逐级诱导宋内志贺菌对环丙沙星耐药,通过提取诱导株诱导前后DNA和总RNA,采用基因测序检测突变和实时荧光定量PCR方法检测acrR、marR基因表达量的变化。结果 2株诱导耐药株中acrR、marR中均不存在基因突变,相对于诱导前acrR、marR Ct值增加,基因表达量降低。结论 acrR、marR基因表达量降低间接导致主动外排基因acrAB-tolC转录水平提高,证实了诱导剂环丙沙星激活了菌株的主动外排系统。
诱导耐药;主动外排;阻遏基因;acrR;marR
Key words:Induced resistance;Active efflux;Repressor gene;acrR;marR
(Chin J Lab Diagn,2015,19:1625)
随着抗生素的应用,志贺菌耐药现象逐年增加,近年来,宋内志贺菌分离的比率逐年增高,有逐渐取代福氏志贺菌的趋势。志贺菌耐药机制涉及多个方面,其中,主动外排系统,尤其是耐药结节分化家族(resistance nodulation division,RND)成员对革兰阴性菌获得性耐药起重要作用,acrAB-tolC是RND家族的典型代表,由药物质子转运子AcrB、膜融合蛋白AcrA和外膜通道蛋白TolC构成,抗菌药物进入细菌的周质或细胞质后,会被内膜上的AcrB捕获,然后AcrB与AcrA形成AcrAB复合体,再与TolC结合形成三联复合体[1,2],AcrAB的表达受多种调节因子的调节,其中阻遏子基因acrR和mar-RAB操纵子的作用最为重要,AcrR抑制子是由位于泵基因上游的阻遏子基因arcR编码的,对acrAB转录起一定程度的抑制作用,MarR是marRAB操纵子的阻遏基因,主要抑制marA,一旦MarA表达增加,同时引起acrA、acrB过量表达,使细菌的耐药性显著增加[3,4]。虽然国内外很多单位对AcrAB-TolC进行了研究,研究对象主要是大肠埃希菌,也有少数研究福氏志贺菌和伤寒沙门菌等[5,6],本文通过检测acrR和marR在菌株被环丙沙星诱导耐药后基因突变与表达水平的变化,来验证宋内志贺菌中主动外排泵的表达变化。
1 材料与方法
1.1 菌株来源
采用我院分离的宋内志贺菌敏感菌株(编号:22、38),以及诱导成功后的宋内志贺菌耐药菌株(22R、38R)。
1.2 主要试剂
TaKaRa Taq kit,Marker D2000,RNAiso Plus,PrimeScriptTMRT reagent Kit with gDNA E-raser(Perfect Real Time),SYBR○RPremix Ex TaqTMII(Tli RNaseH Plus)均购自宝生物工程(大连)有限公司。E-test试条(氨苄西林、头孢噻肟、庆大霉素、环丙沙星、氯霉素、四环素、复方新诺明)购自梅里埃公司。
1.3 主要仪器
实时荧光定量PCR仪GeneLight 2400(厦门安普利),电泳仪JY300C(北京君意),紫外分析仪JY02S(北京君意),高速低温冷冻离心机Eppendorf 5430R(艾本德中国有限公司)。
1.4 方法
1.4.1 宋内志贺菌的诱导 取对数生长期宋内志贺菌,用M-H肉汤校正浓度为0.5麦氏单位,使菌液浓度为1×108CFU/mL,稀释200倍后终浓度为5×105CFU/mL。向菌液中加入浓度为1/2MIC0环丙沙星溶液,充分混匀后塞紧塞子,放入恒温培养箱中37℃振摇培养至细菌生长的对数期,重复以上步骤,不断提高诱导剂环丙沙星的浓度,当环丙沙星的浓度达到1mg/L后,浓度以1mg/L递增,且每一浓度诱导两代以确保耐药,直至诱导剂环丙沙星浓度大于或等于128×MIC0。诱导过程中每诱导3代接种一次M-H琼脂平板,置35℃孵育16-18h,按菌株鉴定步骤进行生化反应和血清型鉴定,确保诱导过程中菌株不被污染。
1.4.2 E-test法测诱导前后菌株的MIC 挑取经35℃培养16-18h后的菌落,用生理盐水调整至0.5McF。在15min内用无菌棉试子蘸取菌液,管壁上挤去多余液体,涂布于M-H平板,平板旋转60度涂布3次并沿边缘涂布一圈,放置数分钟后小心贴上E-test试条。35℃培养16h-18h后读取结果。结果根据2013年版CLSI(M100-S19)标准判读。
1.4.3 宋内志贺菌总DNA的提取 用无菌棉棒挑取适量孵育16-18h诱导株的菌苔溶于50μl双蒸馏水中,95℃水浴5min,12 000rpm离心30秒,含有DNA的上清液于-20℃冰箱保存备用。
1.4.4 acrR与marR基因扩增与测序 根据Gen-Bank公布的acrR和marR基因序列,采用软件Primer Premier 5自行设计引物,见表1中acrR-1与marR-1的引物序列,以总DNA为模板进行PCR反应,PCR反应体系为:5U/μl TaqDNA聚合酶0.5μl、10×缓冲液(Mg2+plus)5μl、10mmol/L dNTP 4μl、25mmol/LMgCl22μl、10μmol/L引物5μl、模板DNA5μl、无菌双蒸水补足总反应体积为50μl,反应条件:95℃预变性5min;94℃变性25s,55℃退火20s,72℃延伸25s,共35个循环;最后72℃延伸5min。反应结束后进行琼脂糖凝胶电泳,扩增产物送宝生物工程(大连)有限公司进行基因测序。
1.4.5 宋内志贺菌总RNA的提取及转录 宋内志贺菌总RNA提取参照RNAiso Plus试剂盒说明,取1μg总RNA按照PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)进行DNA消除,然后进行逆转录。
1.4.6 宋内志贺菌主动外排基因表达水平检测(RT-PCR) 根据GenBank公布的acrR和marR基因序列,采用软件Primer Premier 5自行设计引物,PCR引物合成委托宝生物工程(大连)有限公司完成。RT-PCR引物序列见表1中acrR-2,marR-2。荧光定量PCR反应体系与反应条件。25μl反应体系中:SYBR○RPremix Ex Taq(Tli RNaseH Plus)12.5μl,上下游引物10μM各1μl,cDNA 2 μl,dH2O 8.5μl,RT-PCR循环参数;反应条件:95℃预变性5min;94℃变性25s,52℃退火20s,72℃延伸25s,共35个循环;最后72℃延伸5min。分别读取每个菌株的acrR、marR和16sRNA的Ct值,acrR、marR的Ct值与16sRNA的Ct值之比为其相对含量,比较敏感株与耐药株中acrR和marR的相对含量。
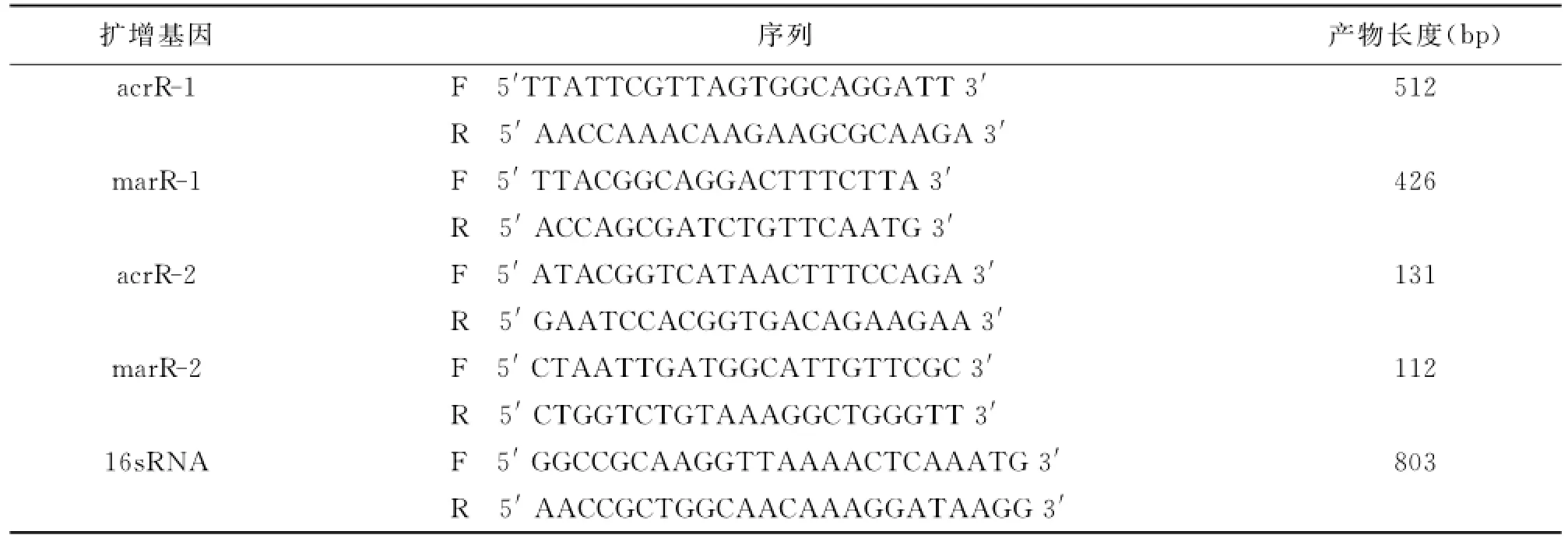
表1 PCR引物序列
2 结果
2.1 环丙沙星诱导结果
经过69代传代诱导,环丙沙星对诱导菌株的MIC达64μg/mL。E-test法测定诱导后菌株的MIC,发现经过诱导的敏感菌株除了对环丙沙星产生耐药外,还对与环丙沙星在结构和作用机制上都不同的其他几种抗菌药(氨苄西林、头孢噻肟、庆大霉素、氯霉素、四环素、复方新诺明)也产生了耐药,经过环丙沙星的诱导,原来的敏感菌株均逐步转变为多重耐药株。诱导前后各抗生素MIC值见表2。
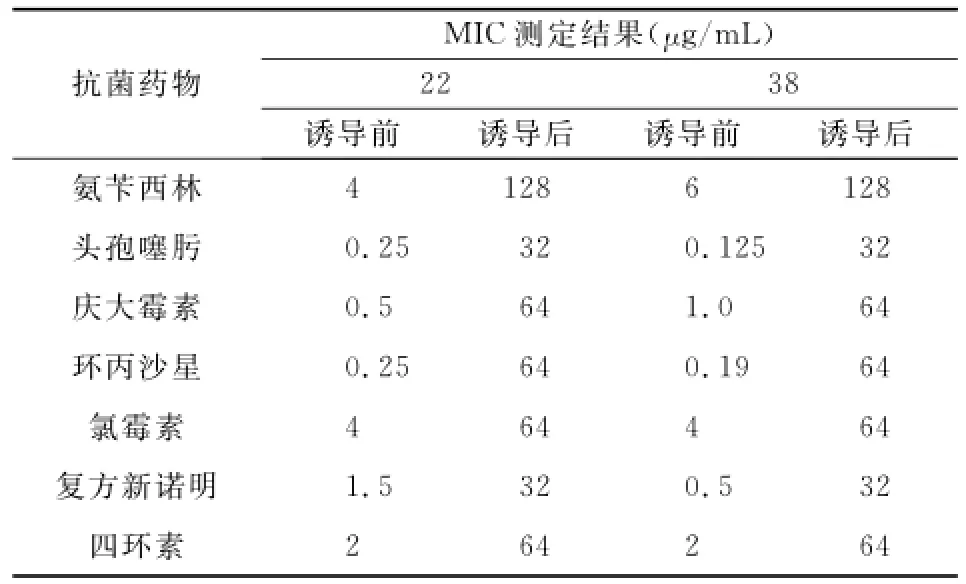
表2 两株宋内志贺菌诱导前后的MIC结果
2.2 宋内志贺菌acrR和marR的PCR扩增产物及其测序结果
2株诱导株均扩增出大小分别为512bp和426 bp的片段,acrR和marR基因扩增产物,经测序与BLAST比对,均未发现有基因突变。
2.3 宋内志贺菌acrR、marR基因表达水平分析结果
读取每个菌株诱导前后acrR、marR的Ct值与16sRNA的Ct值,实时荧光定量曲线图见图1。根据Ct值的比值,诱导耐药株的acrR、marR表达水平明显低于敏感株,见表3。这表明诱导耐药后菌株的主动外排阻遏基因表达水平降低,相应的主动外排基因acrAB-tolC转录水平提高,证实了诱导剂环丙沙星激活了菌株的主动外排系统。
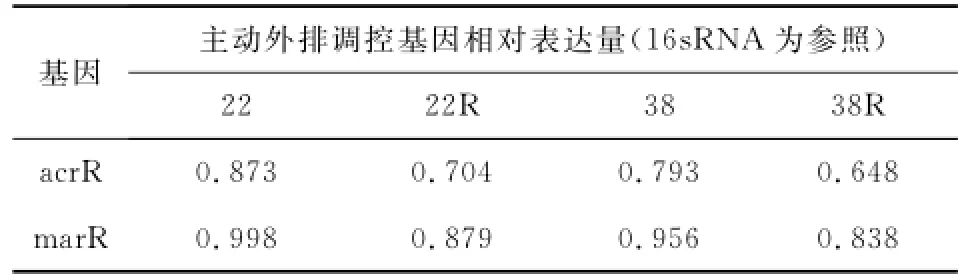
表3 诱导前后acrR和marR的相对表达量
3 讨论
自Levy[7]等提出主动外排系统是细菌主要耐药机制之一后,一个个新的主动外排系统不断被发现,主动外排系统在临床主要致病菌耐药中的重要作用不断被揭示,并为业界所认同,是目前细菌耐药机制研究中的新热点[8]。
图1 实时荧光定量PCR曲线图
本实验用环丙沙星对宋内志贺菌敏感菌株进行诱导耐药,建立环丙沙星耐药株,意外的是诱导后菌株除了对环丙沙星产生耐药外,还对与环丙沙星在结构和作用机制上都不同的其他几种抗菌药(氨苄西林、头孢噻肟、庆大霉素、氯霉素、四环素、复方新诺明)也产生了耐药,而且这几种药物为主动外排泵AcrAB的作用底物。推测这可能是由于诱导剂环丙沙星激活了敏感株的主动外排系统,使敏感株变为多重耐药菌株。
AcrAB的表达受多种调节因子的调节,其中阻遏子基因acrR和marRAB操纵子的作用最为重要。AcrR抑制子是由位于泵基因上游的阻遏子基因arcR编码的,对acrAB转录起一定程度的抑制作用[3,4]。AcrR通常以二聚体形式结合于acrA基因和acrB基因启动子附近的位点,抑制acrA和acrB基因的转录,防止其过量表达;marRAB操纵子包括启动子marO、阻遏基因marR、转录调节因子marA和marB。该操纵子可编码三种蛋白质,MarR(阻遏子)、MarA(转录激活物)和MarB(功能不明蛋白质),marRAB操纵子被激活后,可增强细菌多重耐药性。MarR是由marR基因编码的162个氨基酸大小的蛋白质,它可以结合到marRAB操纵子调控区域,抑制MarA的产生,从而抑制主动外排泵基因的转录[9]。有研究报道当acrR或marR基因突变后,不能有效阻遏acrAB的转录,导致acrAB表达增强,开启主动外排系统,导致因药物外排而产生的耐药[3,4,10]。本项目中对两株诱导株中的acrR和 marR基因进行测序,均未发现有基因突变发生,但通过定量分析,发现诱导耐药株中acrR表达水平下降时,细菌对多种抗生素产生耐药,推测可能是主动外排系统被激活。本实验中未发现acrR和marR的基因突变,可能与实验株有限相关。宋内志贺菌的主动外排泵AcrAB-TolC的影响因素很多,外排机制比较复杂。由于宋内志贺菌标本较少,耐药宋内志贺菌标本更难获得,本试验仅就两株诱导耐药宋内志贺菌的主动外排泵相关的阻遏基因做了研究,将来需广泛收集标本,扩大检测例数作进一步研究。
[1]Blair J M,Piddock LJ.Structure,function and inhibition of RND efflux pumps in Gram-negative bacteria:an update[J].Curr Opin Microbiol,2009,12(5):512.
[2]王松泰,张继瑜,魏小娟,等.福氏志贺菌主动外排基因acrA的克隆及原核表达[J].中国人兽共患病学报,2008,24(10):933.
[3]Pourahmad JR,Jazayeri N.Expression of acrA and acrB Genes in Esherichia coli Mutants with or without marR or acrR Mutations[J].Iran J Basic Med Sci,2013,16(12):1254.
[4]Ballesté-Delpierre C,SoléM,DomènechÕ,et al.Molecular study of quinolone resistance mechanisms and clonal relationship of Salmonella enterica clinical isolates[J].Int J Antimicrob Agents,2014,43(2):121.
[5]Kim JY,Kim SH,Jeon SM,et al.Resistance to fluoroquinolones by the combination of target site mutations and enhanced expression of genes for efflux pumps in Shigella flexneri and Shigella sonnei strains isolated in Korea[J].Clin Microbiol Infect,2008,14(8):760.
[6]Yamasaki S,Nagasawa S,Hayashi-Nishino M,et al.AcrA dependency of the AcrD efflux pump in Salmonella enterica serovar Typhimurium[J].J Antibiot(Tokyo),2011,64(6):433.
[7]Levy SB.Active efflux mechanisms for antimicrobial resistance[J].Antimicrob Agents Chemother,1992,36(4):695.
[8]张 永,唐英春.病原菌质子驱动型外排泵分子机制研究进展[J].国外医药(抗生素分册),2004,25(1):6.
[9]Duval V,McMurry LM,Foster K,et al.Mutational analysis of the multiple-antibiotic resistance regulator MarR reveals a ligand binding pocket at the interface between the dimerization and DNA binding domains[J].J Bacteriol,2013,195(15):3341.
[10]Sato T,Yokota S,Uchida I,et al.Fluoroquinolone resistance mechanisms in an Escherichia coli isolate,HUE1,without quinolone resistance-determining region mutations[J].Front Microbiol,2013,24(4):125.
Changes of repressor gene acrR and marR induced resistant Sonnei Shigella
ObjectiveTo analyze mutation and expression of repressor gene acrR and marR for acrAB-tolC efflux system before and after induced resistant to Ciprofloxacin.Methods The resistance to Ciprofloxacin was induced step by step with 1/2MIC,The total DNA and RNA was extracted from the induced and the primary strains,and then detected the acrR and marR gene mutation and expression with sequencing and real time PCR.Results The results showed that the acrR and marR were not mutated,but declined in two induced sonnei shigellacompared with the wild strains.Conclusions Its indirectly means that the acrAB-tolC transcription declined and the active efflux was activated by Ciprofloxacin.
R378.2+5
A
2014-10-09)
1007-4287(2015)10-1625-05
GAO Pei-zeng,YAO Ming-xiao.
(ClinicalLaboratory of Ji’nan Fourth People’s Hospital,Ji’nan250031,China)
