ERK信号通路在人牙髓干细胞增殖与分化中的作用
2015-04-20马逢乐刘宝刚何欣遥王志华何文喜
马逢乐,刘宝刚,王 娟,何欣遥,王志华,何文喜
(1.军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院牙体牙髓病科,陕西西安710032;2.第二炮兵总医院礼士路门诊部,北京100820)
ERK信号通路在人牙髓干细胞增殖与分化中的作用
马逢乐1,刘宝刚2,王 娟1,何欣遥1,王志华1,何文喜1
(1.军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院牙体牙髓病科,陕西西安710032;2.第二炮兵总医院礼士路门诊部,北京100820)
目的:探讨细胞外信号调节激酶(ERK)通路是否参与对人牙髓干细胞(hDPSCs)增殖及分化过程的调控。方法:用不同浓度的ERK通路抑制剂U0126干预hDPSCs,用CCK-8法检测细胞增殖;在矿化液诱导条件下,用不同浓度U0126干预hDPSCs 14 d,茜素红染色观察矿化结节的形成,RT-PCR检测成骨/成牙相关基因ALP、OCN、DSPP及BSP的表达;Western blot检测U0126(25μmol/L)干预后,ERK通路活性的变化。结果:不同浓度的U0126对hDPSCs的增殖能力无显著影响(P>0.05);与对照组相比,不同浓度的U0126对矿化结节的形成和成骨/成牙相关基因ALP、OCN、DSPP及BSP的表达均有抑制作用,25μmol/L U0126的抑制作用最明显(P<0.05);Western blot结果显示随着时间的延长,p-ERK的表达量逐步增加,在加入25μmol/L U0126后,p-ERK表达量下降(P<0.05)。结论:ERK信号通路可能不参与对hDPSCs增殖的调控,而在促进hDPSCs成骨/成牙分化过程中起调控作用。
人牙髓干细胞(hDPSCs);ERK信号通路;增殖;分化
[Chinese Journal of Conservative Dentistry,2015,25(5):277]
Gronthos等[1](2000)利用collagenaseⅠ和dispase消化法,成功的从牙髓组织中分离出了间充质干细胞,并提出了牙髓干细胞(dental pulp stem cells,DPSCs)的概念。研究表明,牙髓干细胞在不同的诱导剂作用下,可以分化为成牙本质细胞、脂肪细胞和神经样细胞等多种细胞[2-3]。碱性磷酸酶(ALP)、牙本质涎磷蛋白(DSPP)等常被用于鉴定DPSCs分化的特异性指标[4-5]。近年来,DPSCs作为牙齿再生的种子细胞,已成为组织工程应用的重要细胞来源。
研究表明,干细胞的增殖和分化受到多种信号通路的调控,分子机制复杂。Wnt/β-catenin信号通路[6-7]和Notch信号通路[8]在调控神经干细胞增殖、分化中发挥重要作用;NF-κB通路能够调控间充质干细胞[9]和牙周膜干细胞[10]的成骨分化。那么,在DPSCs的增殖和分化过程中有哪些通路参与调控,目前还不清楚。
丝裂原活化蛋白激酶(mitogen activated protein kinases,MAPKs)是细胞内的一类丝氨酸和(或)苏氨酸蛋白激酶,在细胞应激和损伤反应中发挥重要作用。目前为止,已发现了 4种不同的MAPK[11],其中细胞外信号调节蛋白激酶(extracellular signal regulated protein kinase,ERK)是最早被发现的,且被认为与细胞的增殖和分化相关。近年来研究发现,ERK信号通路在调控干细胞增殖、分化过程中发挥重要作用。有研究表明,血管内皮生长因子可以激活MEK/ERK通路和PI3K/Akt通路,从而促进大鼠海马细胞的增殖[12];还有研究表明,IL-2可以激活ERK通路,促进淋巴B细胞的增殖和向浆细胞的分化[13];三氧化矿物凝聚体(MTA)能够激活JNK和ERK通路,促进骨髓间充质干细胞的成骨/成牙本质分化[14];但ERK信号通路是否参与人牙髓干细胞(hDPSCs)的增殖、分化调控,目前研究甚少。本研究通过观察ERK通路抑制剂U0126对hDPSCs的增殖和成骨/成牙分化的影响,从而探讨ERK信号通路在hDPSCs增殖和分化中的作用。
1 材料和方法
1.1 主要材料、试剂和仪器
二氧化碳细胞培养箱(Thermo,美国);超净工作台(上海净化设备有限公司);倒置相差显微镜和照相系统(Olympus,日本);Real time PCR仪(ABIPrism 7500,美国);电泳仪(BIO-RAD,美国);红外成像仪(Odyssey,美国);α-MEM、Ⅰ型胶原酶、Dispase酶、胰蛋白酶(Gibco,美国);胎牛血清(HyClone,美国);β-甘油磷酸钠、地塞米松、维生素C、茜素红(Sigma,美国);U0126(Invivo Gen,美国);CCK-8(南京恩晶生物);RIPA裂解液、5×SDSPAGE Loading Buffer、β-actin单克隆抗体(北京康为世纪);ERK单克隆抗体、p-ERK单克隆抗体(CST,美国);PVDF膜(Millipore,美国);细胞总RNA提取试剂盒(Omega,美国);反转录试剂盒、Real Time-PCR试剂盒(Takara,日本);Tris、甘氨酸、吐温-20(MP,美国);BSA(Thermo,美国);氯化钠、甲醇(国产分析纯)。
1.2 ERK通路抑制剂U0126在hDPSCs成骨/成牙分化过程中的作用
1.2.1 实验分组及矿化诱导
取3代hDPSCs,当细胞长至80%~90%融合时,在矿化液 (2.16 g/Lβ-甘油磷酸钠 、0.05 g/L维生素C、0.785 g/L地塞米松)的条件下,分别加入5、10、25μmol/L的U0126作为实验组,以含100 mL/L胎牛血清的α-MEM作为空白组,以单纯矿化液诱导为对照组,进行后续实验。
1.2.2 CCK-8细胞增殖情况
取3代hDPSCs,当细胞长至约80%~90%融合时,消化离心后收集细胞,以每孔2 000个细胞接种至96孔板,每孔100μL,用不含胎牛血清的α-MEM饥饿24 h,分别加入5、10、25μmol/L的U0126,用含100mL/L胎牛血清的α-MEM作为对照,每组设置5个副孔,分别刺激1、3、5、7 d,每孔加入10μL CCK-8,37℃孵育4 h后酶标仪检测562 nm波长下的吸光值。
1.2.3 茜素红染色
取3代hDPSCs,当细胞长至80%~90%融合时,矿化诱导14 d,弃去诱导液用PBS漂洗3次,40 g/L多聚甲醛固定20 min,去离子水漂洗3次,茜素红染色10 min后肉眼及倒置显微镜下观察矿化结节的形成。
1.2.4 RT-PCR检测
取3代hDPSCs,当细胞长至80%~90%融合时,矿化诱导14 d,弃去诱导液用PBS漂洗3次,用RNA提取试剂盒提取mRNA,用反转录试剂盒反转录成cDNA,以GAPDH(人磷酸甘油醛脱氢酶)作为内参基因,并以GENE BANK数据库设计骨钙素(OCN)、碱性磷酸酶(ALP)、牙本质涎磷蛋白(DSPP)、骨唾液蛋白(BSP)、GAPDH基因的引物(具体引物序列见表1),由上海生工生物合成。按说明书进行Real-Time PCR,每组3个副孔并重复试验3次。

表1 实时定量PCR作用引物名称及序列
1.3 ERK通路抑制剂U0126调控hDPSCs成骨/成牙分化的分子机制
1.3.1 实验分组及矿化诱导
取3代hDPSCs,当细胞长至80%~90%融合时,分别进行矿化诱导,分组为矿化诱导5、15、30、60 min,矿化诱导+25μmol/L U0126,以含100 mL/L胎牛血清的α-MEM作为对照组。
1.3.2 Western Blot检测
上述细胞弃去诱导液用PBS漂洗3次;加入150μL RIPA裂解液裂解细胞,4℃12 000 r/min离心15 min,取上清;加入5×SDS PAGE Loadling Buffer沸水浴10 min后,80 V、30 min,120 V、1 h进行PAGE电泳;电泳结束后2 h、200 mA冰浴转膜;转膜完毕后用50 g/L的BSA室温封闭2 h;一抗4℃孵育过夜,TBST漂洗3次,每次15 min;二抗室温孵育2 h,TBST漂洗3次,每次15 min;用红外成像仪进行结果分析。
1.4 统计学分析
采用SPSS 17.0统计软件进行统计分析,两两比较用t检验,检验水准α=0.05。
2 结果
2.1 ERK信号通路抑制剂U0126对hDPSCs增殖的影响
取3代hDPSCs,CCK-8法检测其1、3、5、7 d细胞增殖情况,结果显示:与对照组比较,5、10、25μmol/L U0126组细胞增殖均无明显差异(P>0.05)(图1)。

图1 CCK-8细胞增殖情况
2.2 ERK信号通路抑制剂U0126对hDPSCs成骨/成牙分化的影响
2.2.1 茜素红染色结果
取3代hDPSCs,矿化诱导14 d后,茜素红染色结果显示,对照组矿化结节明显多于空白组,而与对照组相比,实验组矿化结节明显减少,25μmol/L U0126组减少最明显,而5μmol/L U0126与10μmol/L U0126组间无显著差异(图2),定量结果与肉眼观察结果一致(图3)。
2.2.2 RT-PCR检测结果
取3代hDPSCs,矿化诱导14 d后,RT-PCR检测成骨/成牙相关基因ALP、OCN、DSPP和BSP的表达情况,结果显示与空白组相比,对照组ALP、OCN、DSPP和BSP的表达均明显上调,而除5μmol/L U0126组ALP的表达与对照组无显著差异(P>0.05),10μmol/L U0126与25μmol/L U0126组ALP、OCN、DSPP和BSP的表达和对照组相比均明显下调,且以25μmol/L U0126组下调最明显(P<0.05)(图4)。
2.3 ERK通路抑制剂U0126调控hDPSCs成骨/成牙分化的分子机制
取3代hDPSCs,短时间矿化诱导并加入ERK信号通路抑制剂U0126(25μmol/L)后,Western blot检测ERK、p-ERK蛋白表达结果显示,随着时间的延长,p-ERK的表达量明显增加,并在加入抑制剂U0126后,表达量显著下降(图5)。

图2 各组茜素红染色情况
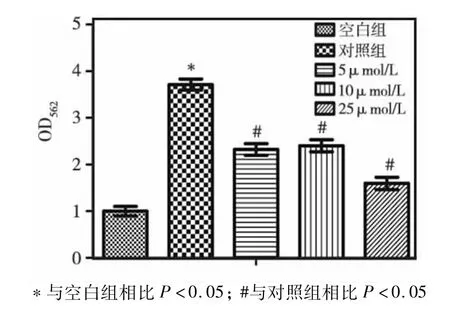
图3 茜素红染色定量结果

图4 成骨/成牙相关基因的表达水平比较

图5 ERK信号通路的蛋白表达和p-ERK相对表达量
3 讨论
大量研究表明,在干细胞增殖分化过程中有多种信号途径参与调控。而ERK信号途径作为最早发现的MAPK信号通路中的一条在调控干细胞的增殖分化中发挥重要作用。本研究通过CCK-8、茜素红染色、RT-PCR研究ERK信号通路抑制剂U0126对hDPSCs增殖、分化的影响,并进一步通过Western blot来验证ERK信号通路在hDPSCs分化过程中的激活情况。
有文献表明,同型半胱氨酸能够通过下调p-ERK的蛋白表达量来抑制神经干细胞NSCs的增殖[15];还有研究表明,上皮调节蛋白EREG能够增加牙乳头干细胞SCAPs中ERK1/2、MEK和JNK的磷酸化水平,从而促进SCAPs的增殖[16];多肽物质IKVAV可以通过促进磷酸化的MAPK/ERK和磷酸化的PI3K/Akt水平增高,来促进骨髓间充质干细胞BMMSCs的增殖,且呈浓度和时间依赖性[17];本结果显示与对照组相比,1、3、5、7 d不同浓度U0126刺激组的hDPSCs细胞增殖均无明显变化(P>0.05),表明ERK信号通路可能不参与对hDPSCs增殖的调控,这与文献报道有所差别,可能与组织来源,细胞种类,所处微环境及时间等综合因素的不同有关。
矿化结节是反应干细胞成骨/成牙分化的重要指标之一。本研究茜素红染色结果显示,与对照组相比,实验组矿化结节明显减少,25μmol/L U0126组减少最明显,而5μmol/L U0126与10μmol/L U0126组组间无显著差异,说明ERK通路抑制剂U0126能够抑制hDPSCs的成骨/成牙分化。
ALP、OCN、DSPP、BSP、CollagenⅠ、DMPs等是成牙本质细胞分泌的细胞外基质蛋白,与牙髓干细胞的成骨/成牙分化密切相关,故本研究选取ALP、OCN、DSPP和BSP作为研究对象。成骨诱导14 d后,RT-PCR检测结果显示,与空白组相比,经矿化诱导后,对照组的ALP、OCN、DSPP和BSPmRNA表达量均明显上调(P<0.05),而在加入不同浓度的ERK信号通路抑制剂U0126后,除5μmol/L U0126组ALP的表达与对照组无显著差异(P>0.05)外,10μmol/L U0126与25μmol/L U0126组ALP、OCN、DSPP和BSP的表达和对照组相比均明显下调,且以25μmol/L U0126组下调最明显(P<0.05),这与茜素红染色结果趋势基本一致,说明ERK通路抑制剂U0126能够抑制hDPSCs的成骨/成牙分化。5μmol/L U0126组ALP的表达与对照组无显著差异,可能是因为ALP为早期的成骨分化指标,经14 d长时间矿化诱导后表达变化可能不明显,也可能与5μmol/L的浓度较低,抑制剂效果不明显有关。
为了进一步验证ERK信号通路在hDPSCs成骨/成牙分化中的作用,我们通过Western blot检测p-ERK的蛋白表达量。结果显示,随着时间的延长,p-ERK蛋白表达量逐渐增加,在加入25μmol/L U0126后,表达量明显下降(P<0.05),进一步说明了ERK信号通路参与调控hDPSCs的成骨/成牙分化。有文献报道,醋酸氯地孕酮(CMA)能够激活ERK信号通路,促进骨髓间充质干细胞BMMSCs向成骨细胞分化且出现钙沉积,而抑制BMMSCs的成脂分化[18];在胚胎干细胞中,成纤维生长因子7能够激活ERK/RUNX2信号通路来促进成骨分化[19];这些与本研究中ERK信号通路可能参与调控促进hDPSCs的成骨/成牙分化的结果相一致。然而,Suzuki等[20]研究发现,ERK途径对小鼠胚胎成骨细胞MC3T3-E1的增殖起重要作用,对其分化作用不明显;廖清船等[21]发现,阻断ERK通路后,对已分化的成骨细胞ALP活性,钙沉积量无抑制作用,表明ERK可能只参与了该细胞的增殖调节;孔维霞[22]等采用骨片法培养C57/BL小鼠骨实质来源的MSCs,取第4代MSCs进行成骨诱导,发现ERK通路可能在早期有负调控作用;这些与本结果相反,可能是由于细胞种属来源不同和抑制剂作用浓度不同造成的。本结果表明ERK信号通路可能在hDPSCs的成骨/成牙分化过程中起调控作用,而不参与对其增殖的调控。那么,是否有其他信号通路参与,它们是如何相互作用进行调控的,还需进一步研究讨论。
[1]Gronthos S,Gronthos M,Mankani J,et al.Postnatal human dental pulp stem cells(DPSCs)in vitro and in vivo[J].Proc Natl Acad Sci U SA,2000,97(25):13625-13630.
[2] 刘宏胜,白小文,杨媛,等.人类年轻恒牙牙髓干细胞体外多向分化的能力[J].北京大学学报:医学版,2007,39(1):41-45.
[3] 何飞,谭颖徽,张纲.人牙髓干细胞的体外培养和鉴定[J].华西口腔医学杂志,2005,23(1):75-78.
[4]Shiba H,Mouri Y,Komatsuzawa H,et al.Enhancement of alkaline phosphatase synthesis in pulp cells co-cultured with epithelial cells derived from lower rabbit incisors[J].Cell BiolInt,2003,27(10):815-823.
[5]Papagerakis P,Berdal A,Mesbah M,et al.Investigation of osteocalcin,osteonectin,and dentin sialophosphoprotein in developing human teeth[J].Bone,2002,30(2):377-385.
[6]Brembeck FH,Rosário M,BirchmeierW.Balancing cell adhesion and Wnt signaling,the key role of beta-catenin[J].Curr Opin Genet Dev,2006,16(1):51-59.
[7]Munji RN,Choe Y,Li GN,etal.Wnt signaling regulates neuronal differentiation of cortical intermediate propenitors[J].J Neurosci,2011,31(5):1676-1687.
[8]Sang L,Coller HA,Roberts JM.Control of the reversibility of cellular quiescence by the transcriptional repressor HES1[J].Science,2008,321(5892):1095-1100.
[9]Hess K,Ushmorov A,Fiedler J,et al.TNFalpha promotes osteogenic differentiation of humanmesenchymal stem cellsby triggering the NF-kappaB signaling pathway[J].Bone,2009,45(2):367-376.
[10]Chen X,Hu C,Wang G,et al.Nuclear factor-kappaB modulates osteogenesisof periodontal ligament stem cells through competition with beta-catenin signaling in inflammatory microenvironments[J].Cell Death Dis,2013,4:e510.
[11]Wang L,Kwak JH,Kim SI,etal.Transforming growth factorbeta1 stimulates vascular endothelial growth factor 164 viamitogen-activated protein kinase kinase 3-p38alpha and p38delta mitogen-activated protein kinase-dependent pathway in murine mesangial cells[J].J Biol Chem,2004,279(32):33213-33219.
[12]Fournier NM,Lee B,Banasr M,et al.Vascular endothelial growth factor regulates adult hippocampal cell proliferation through MEK/ERK-and PI3K/Akt-dependent signaling[J].Neuropharmacology,2012,63(4):642-652.[13]Le Gallou S,Caron G,Delaloy C,et al.IL-2 requirement for human plasma cell generation:coupling differentiation and proliferation by enhancing MAPK-ERK signaling[J].J Immunol,2012,189(1):161-173.
[14]Wang Y,Li J,SongW,etal.Mineral trioxide aggregate upregulates odonto/osteogenic capacity of bone marrow stromal cells from craniofacial bones via JNK and ERK MAPK signalling pathways[J].Cell Prolif,2014,47(3):241-248.
[15]Yan H,Zhang X,Luo S,etal.Effects of homocysteine on ERK signaling and cell proliferation in fetal neural stem cells in vitro[J].Cell Biochem Biophysv,2013,66(1):131-137.
[16]Cao Y,Xia DS,Qi SR,etal.Epiregulin can promote proliferation of stem cells from the dental apical papilla via MEK/Erk and JNK signalling pathways[J].Cell Prolif,2013,46(4):447-456.
[17]Li B,Qiu T,Zhang P,et al.IKVAV regulates ERK1/2 and Akt signalling pathways in BMMSC population growth and proliferation[J].Cell Prolif,2014,47(2):133-145.
[18]Kim JM,Lee JE,Ryu SH,et al.Chlormadinone acetate promotes osteoblast differentiation of humanmesenchymal stem cells through the ERK signaling pathway[J].Eur J Pharmacol,2014,726:1-8.
[19]Jeon YM,Kook SH,Rho SJ,et al.Fibroblast growth factor-7 facilitates osteogenic differentiation of embryonic stem cells through the activation of ERK/Runx2 signaling[J].Mol Cell Biochem,2013,382(1-2):37-45.
[20]Suzuki A,Guicheux J,Palmer G,etal.Evidence for a role of p38 MAPK kinase in expression of alkaline phosphatase during osteoblastic cell differentiation[J].Bone,2002,30(1):91-98.
[21]廖清船,肖洲生,秦艳芳,等.P44/42和p38 MAPKs在骨髓间充质干细胞向成骨细胞分化中发挥不同的功能[J].中国骨质疏松杂志,2004,10(3):267-271.
[22]孔维霞,朱恒,江小霞,等.MAPK通路参与小鼠实质来源间充质干细胞向成骨的分化[J].中国实验血液学杂志,2010,84(4):981-985.
The effect of ERK signaling pathway on the proliferation and differentiation of human dental pulp stem cells
MA Feng-le*,LIU Bao-gang,WANG Juan,HE Xin-yao,WANG Zhi-hua,HEWen-xi
(*State Key Laboratory of Military Stomatology,Department of Operative Dentistry and Endodontics,School of Stomatology,The Fourth Military Medical University,Shaanxi Key Laboratory of Stomatology,Xi′an 710032,China)
AIM:To investigate the function of ERK signaling pathway in the regulation of proliferation and differentiation of human dental pulp stem cells(hDPSCs).METHODS:hDPSCs were treated with different concentrations of the ERK signaling pathway inhibitor U0126,and the proliferation wasmeasured by CCK-8 method.The mRNA expression of ALP,OCN,DSPP and BSPwas determined by RT-PCR.Alizarin red stainingwas used to detect themineralized nodules.ERK signaling pathway activation was analyzed by Western blot.RESULTS:ERK inhibitor U0126 had no significant influence on the proliferation ofhDPSCs.U0126 inhibited osteogenic differentiation and dentinogenic differentiation as observed by Alizarin red staining.U0126,especially at25μmol/L,down-regulated the expression of ALP,OCN,DSPPand BSP.U0126 at25μmol/L significantly inhibited ERK signaling pathway activation.CONCLUSION:ERK signaling pathway is probably not involved in the proliferation of hDPSCs,but plays an important role in the osteogenic and dentinogenic differentiation of hDPSCs.
human dental pulp stem cells(hDPSCs);ERK signaling pathway;proliferation;differentiation
R780.2
A
1005-2593(2015)05-0277-06
10.15956/j.cnki.chin.j.conserv.dent.2015.05.004
2014-12-19;
:2015-03-30
国家自然科学基金资助(81271125,81470733)
马逢乐(1989-),女,回族,陕西人。硕士生(导师:何文喜)
何文喜,E-mail:hewenxi@fmmu.edu.cn
