基于不同取样尺度的油松针阔混交林物种多样性
2015-03-23邓宏兼李卫忠王广儒
邓宏兼,李卫忠,曹 铸,王 庆,王广儒
基于不同取样尺度的油松针阔混交林物种多样性
邓宏兼1,李卫忠1,曹 铸1,王 庆1,王广儒2
(1.西北农林科技大学 林学院,陕西 杨凌712100;2.陕西省延安市黄龙山林业局,陕西 黄龙716000)
物种多样性是描述群落组成结构的重要指标,其空间格局及成因是物种多样性研究的基础和重要内容。利用陕西省黄龙山林区油松Pinus tabulaeformis针阔混交林1 hm2样地数据,对物种多样性随尺度的变化趋势,不同尺度下各功能层物种多样性特征,乔木层结构与林下植被物种多样性的关系进行了研究,得到以下结论:①丰富度指数、Simpson指数及Shannon-Wiener指数均随尺度增大先快速增大后趋于平缓,丰富度指数空间变异呈现为不规则变化,Simpson指数及Shannon-Wiener指数空间变异则随尺度增大先减小后趋于稳定,Pielou均匀度指数随尺度增大先急剧减小后趋于稳定,其空间变异则随尺度增大而逐步减小。②各功能层物种多样性的关系在5 000 m2及更大尺度上保持一致,灌木层具有最高的物种多样性,草本层物种丰富度较乔木层低,其物种分布均匀度则比乔木层高。③通过对乔木层密度及混交度与灌草层多样性各指数进行灰色关联度分析,得出乔木层结构与灌草层物种多样性具有明显的关联。图2表3参30
森林生态学;油松林;物种多样性;取样尺度;灰色关联度分析;混交度;黄龙山林区
物种多样性是群落结构的重要特征,其空间格局及其成因是研究物种多样性的基础和重要内容[1]。国内外关于物种多样性的研究主要集中于环境梯度对物种多样性的影响,不同演替阶段物种多样性的变化,不同群落类型物种多样性的差异,以及人为干扰、放牧与外来入侵等对物种多样性的影响及其响应。生态学模式和生态学过程与取样尺度都密切相关[2],尺度对于物种的空间分布具有一定的影响[3],而物种多样性各指数对尺度亦均有一定程度的依赖性。取样尺度的变化对Simpson指数、Shannon-Wiener指数及Pielou指数均具有明显的影响。具体表现为:随尺度增大,各指数表现出不连续变化,其变异系数随尺度的增加而逐渐减小[1,4-5]。在不同尺度下物种多样性呈现出较大的差异,单一尺度下的物种多样性难以全面客观地描述整个群落的组成结构特征[6]。物种多样性空间格局在不同尺度下也同样呈现出一定差异,物理环境因子与物种多样性的关系在不同尺度下也不尽相同[7]。因此,在研究物种多样性时,不应忽略取样尺度与物种多样性的关系,也不能以单一尺度下的物种多样性测度结果代表整个区域的物种多样性特征。群落组成树种的数量和空间配置不同,形成了不同的结构格局,其物种多样性也会有一定差别[8]。乔木层的植被结构在某种程度上可以控制草本层的出现和生长[9],林分密度对林下植被的种类和组成影响显著[10],不同的林分类型,其林下植被物种多样性也有一定差异,混交林的物种多样性要明显高于纯林[11]。李利平等[12]通过对北京山区不同区域油松Pinus tabulaeformis林植物多样性的影响因子进行主成分分析,也表明林分密度是影响植物多样性的主要因素。目前,基于大样地数据的黄土高原地区物种多样性及其尺度效应的研究尚未见报道,关于乔木层结构与林下植被物种多样性关系的探讨也少见涉及。本研究利用黄龙山林区油松针阔混交林1 hm2大样地数据,对不同取样尺度下物种多样性的变化进行了研究,分析了物种多样性随取样尺度的变化趋势,并引入空间结构参数混交度,探讨了乔木层结构与林下植被物种多样性的关系。
1 研究区概况
研究样地位于陕西延安市黄龙山林业局虎沟门林场,地处陕北黄土高原南部,位于延安市黄龙、宜川两县交界处。地理坐标为35°37′46.6″N,109°58′39.6″E,平均海拔1 450 m。样地所在区域气候类型为中纬度暖温带大陆性季风气候区,年平均气温为8.6℃,极端最高温度36.7℃,极端最低温度-27.2℃,无霜期175 d左右,年平均降水量602.7 mm。土壤类型主要为褐土,地带性植被为暖温带落叶阔叶林带,主要成林树种为油松,辽东栎Quercus wutaishanica,白桦Betula platyphylla,山杨Populus davidiana,茶条槭Acer ginnala,山桃Prunus davidiana等,以这些树种为主形成的纯林或混交林呈镶嵌性分布,形成当地主要的森林植被。
2 研究方法
2.1 样地设置及调查
于2013年8月在黄龙山林区虎沟门林场设置100 m×100 m大样地,林分类型为天然油松针阔混交林,树木平均年龄32 a,主要树种为油松、辽东栎、白桦,伴生有小叶杨Populus simonii和茶条槭等。样地平均海拔为1 450 m,平均坡度25°,总体郁闭度0.7左右。以20 m为间隔将其划分为25个20 m×20 m小样地作为调查单元。对各个20 m×20 m小样地内胸径大于等于3 cm的乔木进行每木检尺,分别记录树高、胸径、冠幅、枝下高,并用铝牌进行编号。以东西方向边界为x轴,南北方向边界为y轴,分别测量每一乔木到x轴和y轴的垂直距离,记录坐标,作为尺度划分的依据。在每一样地中心位置用罗盘仪测定坡向和坡度,用手持全球定位系统(GPS)测定海拔及地理坐标。各个20 m×20 m小样地内随机设置4个5 m×5 m的灌木样方,4个1 m×1 m的草本样方,分别记录种名、高度、多度、冠(丛)径及盖度,并记录样方中心坐标。
2.2 研究方法
2.2.1 尺度划分及分析方法 根据乔木坐标数据,将样地划分为不同的尺度(尺度划分及划分后样地数见表1),分别计算400~5 000 m2各尺度的物种多样性均值和变异系数及6 400~10 000 m2各尺度物种多样性均值,绘出均值—尺度曲线,以描述物种多样性随尺度的变化趋势,绘出变异系数—尺度曲线,以描述尺度内划分后样地间空间变异程度。灌草层则根据灌草样方中心坐标进行尺度划分。用公式(1)和公式(2)计算均值及变异系数。均值:

式(1)中:Mi为第i尺度下的物种多样性各指数均值,Mij为第i尺度下第j样方内物种多样性各指数值,n为第i尺度下样方数。变异系数:

式(2)中:sd为标准差,μ为平均值。
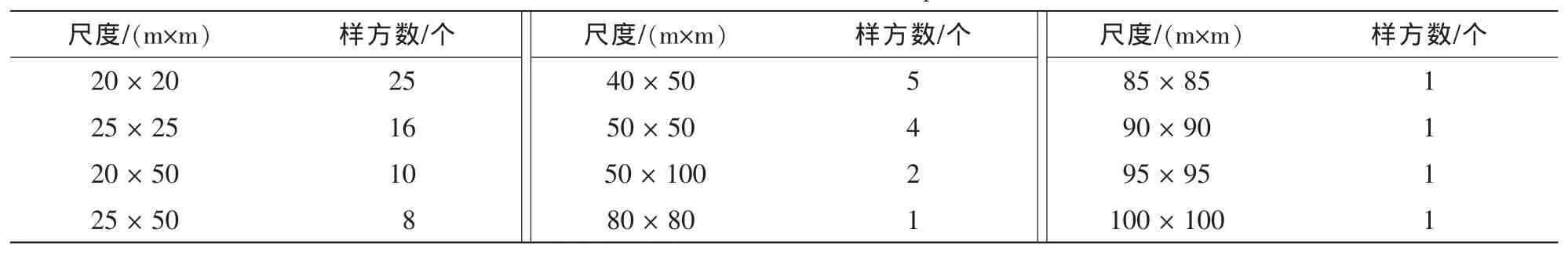
表1 尺度划分及划分后样地数Table 1 Scale division and the number of plots after division
2.2.2 重要值测定 利用重要值作为多度指标能够比较准确的测定多样性各指数。乔木层将相对密度、相对优势度及相对高度的平均值作为其重要值,灌草层由于个体间差异较大,存在较多的丛生及簇生个体,因此利用相对盖度和相对高度的平均值作为其重要值,公式如下[11]:

2.2.3 树种混交度测定 树种混交度(M)被定义为参照树i的4株最近相邻木与参照树不属同种的个体所占的比例,其公式如下[13]:

2.2.4 多样性指数测定 本研究多样性测度采用物种丰富度指数(R),多样性指数(Simpsion指数,Shannon-Wiener指数)及Pielou均匀度指数,公式分别如下[14-16]:

式(6)~式(9)中:S为一定样方内的物种数,Pi=Ni/N,其中Ni为第i个物种的重要值,N为所有物种的重要值之和。
2.2.5 邓氏灰色关联分析及其计算方法 灰色关联分析是灰色理论中的一种分析方法,用来分析系统中母因素与子因素的关系密切程度,从而判断引起该系统发展的主要和次要因素[17]。邓氏关联度分析理论由中国学者邓聚龙创立。该理论提出了灰色关联空间,并在此空间建立非函数型的序模型,以克服一般统计方法追求大样本,渴望典型分布,计算工作量大,有时与定性分析大相径庭的弱点[18]。下面介绍其计算步骤[19]。①无量纲化。对子因素进行无量纲化,其公式为:m。②求差序列、最大差和最小差。据计算母因素序列与各子因素序列的绝对差值,形成如下绝对差值矩阵:同时找出差值矩阵中的最大数(最大差)与
最小数(最小差),并分别表示为Δmax与Δmin。③计算关联系数其中:ρ为分辨系数,在[0,1]内取值,一般取0.5,得关联矩阵:④计算关联度。
2.3 数据处理
采用SPSS 19.0和Excel 2003进行统计分析和制图,利用空间格局分析软件进行混交度计算,用灰色关联度分析软件进行邓氏灰色关联度计算及分析。
3 结果与分析
3.1 取样尺度对物种多样性的影响
为探索研究区域物种多样性各指数对于尺度变化的敏感程度,本研究划分出12个不同的取样尺度分别对多样性各指数及其变异系数进行了测度,其结果如图1所示。由图1可以看出:①丰富度指数随取样尺度的增大先均匀增大后趋于平缓,其空间变异随尺度的变化趋势较不规则,其原因可能是本研究选取的研究样地为混交林,物种数较多且分布不十分均匀。②Simpson指数及Shannon-Wiener指数随取样尺度的增大均表现为先快速增大后较为平缓,其空间变异则均随尺度增大先减小后趋于稳定。③Pielou指数随尺度增大呈现出先减小后保持稳定趋势,其空间变异随尺度的增大而逐步减小。
根据尺度分析结果,丰富度指数和Pielou均匀度指数在5 000 m2后趋于稳定,Simpson多样性指数和Shannon-Wiener多样性指数则从2 000 m2后就逐步趋于稳定。综合而言,研究区域物种多样性在取样尺度达到5 000 m2后稳定性较好。
3.2 物种多样性层间关系
各功能层由于其生态学过程、生活史对策各不相同,其群落组成及群落结构也存在较大差异。为了探讨各层间物种多样性的关系,分别对乔木层及灌草层的物种多样性进行了测度,其结果如图2所示.由图2可以看出:在5 000 m2及更大尺度上,各功能层物种多样性关系保持一致,但不同指数的测定结果不尽相同。丰富度指数与Simpson指数表现为灌木>乔木>草本,而Shannon-Wiener指数及Pielou指数则表现为灌木>草本>乔木。
Simpson指数以一定区域内出现不同种的概率,即物种丰度作为测度依据,而Shannon-Wiener指数则综合考虑种的丰度及各物种分布的均匀度。根据2种多样性指数测定结果出现的差异可以得出:乔木层物种丰富度要高于草本层,但草本层物种分布均匀度较乔木层高。
3.3 乔木层结构对林下植被物种多样性的影响
本研究选取了乔木层密度及混交度2项指标,以探讨乔木层结构与灌草层物种多样性的关系。密度主要影响林内光照,而混交度则主要影响种群竞争,进而影响土壤养分。较小的环境跨度对物种多样性不会产生明显的影响[20]。由于研究样地海拔跨度较小,坡度、坡向、水分等环境因子相对均匀,差异不大,因此,可以忽略环境因子的影响而单独考虑乔木层结构与灌草层物种多样性的关系。由于涉及到空间结构参数混交度的测定,而空间结构的研究需要在大尺度条件下进行,一般要求最小取样面积为2 500 m2[21],因此,对于乔木层结构与灌草层物种多样性关系的研究在2 500 m2及5 000 m2尺度下进行。在2种不同尺度下分别对乔木层密度、混交度及灌草层物种多样性各指数进行了测定,其结果如表2所示。由表2可以看出:2种不同尺度下,各物种多样性指数随密度及混交度的变化略有差异,但大体呈现相同规律。

表2 乔木层密度及混交度与灌草层物种多样性关系Table 2 Relationship between the density of tree layer and the mingling of shrub-grass layer

图2 各功能层物种多样性对比Figure 2 Comparison of species diversity between layers
为了量化乔木层结构与灌草层物种多样性间的关联程度,本研究采用邓氏灰色关联分析法,选取分辨系数ρ=0.5,利用灰色关联度分析软件分别计算了乔木层密度及混交度与灌草层物种多样性的关联度,其结果如表3所示。由表3可以看出:乔木层密度及混交度与灌草层各物种多样性指数的关联度均大于0.5,说明乔木层密度及混交度与灌草层物种多样性均有较明显的关联,即乔木层结构会对灌草层物种多样性造成影响。灌草层Simpson指数与混交度的关联度均大于其与密度的关联度,Shannon-Wiener指数与Pielou指数则在灌草层表现出不同的规律,灌木层Shannon-Wiener指数与密度的关联度大于其与混交度的关联度,草本层则相反,灌木层Pielou指数与密度的关联度小于其与混交度的关联度,而草本层亦表现出相反的规律。
各物种多样性指数与密度的关联度大小关系为Shannon-Weiner指数 (灌木)>Pielou指数 (灌木)>Simpson指数(灌木)>Pielou指数(草本)>Shannon-Wiener指数(草本)>Simpson指数(草本);各指数与混交度的关联度则表现为Pielou指数(灌木)>Simpson指数(灌木)>Simpson指数(草本)>Shannon-Wiener指数(草本)>Shannon-Wiener指数(灌木)>Pielou指数(草本)。相对于草本层,灌木层物种多样性与乔木层结构的关系更加密切。

表3 乔木层密度及混交度与灌草物种多样性关联度Table 3 Correlation degree of density and mingling of tree layer with species diversity of shrub-grass layer
4 结论与讨论
4.1 物种多样性的尺度效应
植物群落在空间上的分布并不是均一不变的,环境梯度和生境异质性的存在,使得群落的分布会发生一定程度的空间变异[22],对其物种多样性的空间分布和动态也必然会造成一定的影响。而由于物种多样性的空间异质性,不同尺度下的物种多样性测度结果也必然存在一定的差异[4]。小取样尺度下,由于样地面积较小,所容纳的样本量有限,因此,其物种空间分布差异性较小,从而导致物种多样性较低,而当取样尺度增大时,由于样本量的增加,其物种多样性也会随之增加,研究物种多样性的尺度效应是了解物种多样性空间分布动态的基础。黄忠良等[23]及王志高等[1]先后对鼎湖山地区群落多样性及物种多样性进行了研究,表明物种多样性的空间变异随尺度增大有减小的趋势,且在尺度达到一定阀值后趋于稳定。叶万辉等[4]对北京雾灵山植物群落多样性的研究也表明:在一定尺度范围内,尺度对α多样性具有强烈影响,α多样性的空间变异随尺度的增大逐渐减弱,Shannon-Wiener指数随尺度增大呈现出先急剧增大后趋于平缓的规律。本研究对黄龙山林区油松针阔混交林物种多样性尺度效应的研究结果表明:丰富度指数随尺度增大先均匀增大后趋于平缓,其空间变异随尺度的变化趋势较不规则;Simpson及Shannon-Wiener多样性指数随尺度增大均表现出先快速增大后趋于平稳的趋势,其空间变异随尺度增大先逐步减小后趋于平稳;Pielou指数则随尺度增大先减小后保持稳定,其空间变异随尺度增大先逐步减小后趋于稳定。本研究中物种多样性达到稳定的尺度阀值要大于以往其他区域相关研究的尺度阀值,说明物种多样性达到稳定的尺度阀值在不同区域及不同林分类型中存在一定差异。
在物种多样性的早期研究中,其尺度效应并没有引起太多的关注,调查数据质量及空间设置方法的局限性也对相关研究造成了一定的制约[7]。只有极少数的生态学家对此进行过研究,如Palmer等[24]于1994年对不同尺度下物种多样性空间格局的探讨,尔后,陆续有一些关于物种多样性尺度效应的研究结果发表。虽然物种多样性存在尺度效应已经具备了有力的科学支撑,但在大部分的物种多样性研究中,考虑到尺度效应的仍然较少,且基本在单一尺度下进行,其研究结果也无法排除尺度的影响。随着群落结构及空间动态研究的深入,作为其重要内容之一,物种多样性的尺度效应也应受到重视。因此,在今后的物种多样性研究中,应将物种多样性的尺度效应分析作为研究的基础,并尽量避免在单一尺度下探讨物种多样性特征,以排除尺度效应对研究结果的影响。
4.2 各功能层物种多样性比较
地理区域是影响物种多样性的重要因素之一。在不同的区域,由于其环境条件的差异,物种多样性也存在较大差别[25]。在本研究中,灌木层表现出最高的物种多样性,草本层虽然物种丰度较乔木层低,但其物种分布的均匀度要比乔木层高,这与之前该区域单一尺度下油松群落物种多样性的研究结果略有差异[26],与以往其他地区关于物种多样性层间关系的研究结果也不尽相同[11,27-29]。这说明功能层间物种多样性的关系也受到取样尺度及研究区域的影响。
4.3 乔木层结构对林下指被物种多样性的影响
乔木层与灌草层作为一定空间内共存的植物群体,其生态学过程并非独立进行,而是存在一定的相互影响及相互制约。乔木层密度与林内光照具有密切的关系,林内光照条件随乔木层密度增大而变差,而光照条件的改变相应的会影响林下植被的生长。混交度同样会对林内生境条件造成影响,混交度越高,表示物种分布越均匀,对于资源的需求亦更加分散。由于同种树木对光照、水分和营养需求相同,理论上相互间对这些生长要素的竞争也最为激烈,因此,随着混交度的增大,林木竞争压力会逐渐减小,空间分布特征也更加优化[30]。而竞争程度的变化相应的会改变土壤、水分及营养条件,进而影响林下植被的生长发育。在本研究中,乔木层密度及混交度与灌草物种多样性表现出较为明显的关联,恰恰说明了乔木层结构与灌草层物种多样性具有明显的关系。因此,在森林经营管理过程中,应营造适度混交林,优化林分结构,以提高林下植被总体物种多样性。
由于本研究数据量有限,无法进行基于大量样本的相关性分析,要揭示乔木层结构与灌草层物种多样性是否具有显著的线性关系,仍有待进一步研究。
5 致谢
感谢王庆、王道亮、曹铸、牛硕、谈多多及黄龙山林业局对外业工作提供的帮助。
[1] 王志高,叶万辉,曹洪麟,等.鼎湖山季风常绿阔叶林物种多样性指数空间分布特征[J].生物多样性,2008,16(5):454-461. WANG Zhigao,YE Wanhui,CAO Hongling,et al.Spatial distribution of species diversity indices in a monsoon evergreen broadleaved forest at Dinghushan Mountain[J].Biodiversity Sci,2008,16(5):454-461.
[2] WIENS J A.Spatial scaling in ecology[J].Funct Ecol,1989,3(4):385-397.
[3] BELLEHUMEUR C,LEGENDRE P,MARCOTTE D.Variance and spatial scales in a tropical rain forest:changing the size of sampling units[J].Vegetatio,1997,130(1):89-98.
[4] 叶万辉,马克平,马克明,等.北京东灵山地区植物群落多样性研究(Ⅸ)尺度变化对α多样性的影响[J].生态学报,1998,18(1):10-14. YE Wanhui,MA Keping,MA Keming,et al.Studies on plant community diversity in Donglingshan Mountain,Beijing, China(Ⅸ)The influence of scale on α diversity[J].Acta Ecol Sin,1998,18(1):10-14.
[5] HE Fangliang,LEGENDRE P,BELLEHUMEUR C,et al.Diversity pattern and spatial scale:a study of a tropical rain forest of Malaysia[J].Environ Ecol Stat,1994,1(4):265-286.
[6] WHITTAKER R H.Evolution and measurement of species diversity[J].Taxon,1972,21(2/3):213-251.
[7] KALLIMANIS A S,HALLEY J M,VOKOU D,et al.The scale of analysis determines the spatial pattern of woody species diversity in the Mediterranean environment[J].Plant Ecol,2008,196(1):143-151.
[8] 哀建国,梅盛龙,刘胜龙,等.浙江凤阳山自然保护区福建柏群落物种多样性[J].浙江林学院学报,2006,23 (1):41-45. AI Jianguo,MEI Shenglong,LIU Shenglong,et al.Species diversity of Fokienia hodginsii community in Nature Reserve of Mount Fengyang in Zhejiang Province[J].J Zhejiang For Coll,2006,23(1):41-45.
[9] 宫田逸夫,周瑞昌.次生林乔木层植被结构对草本层发育的影响[J].自然资源研究,1985(增刊1):66-73. MIYADA Itsuo,ZHOU Ruichang.Effects of tree layer structure on grass development in secondary forest[J].Nat Resour Stud,1985(supp 1):66-73.
[10] 康冰,刘世荣,蔡道雄,等.马尾松人工林林分密度对林下植被及土壤性质的影响[J].应用生态学报,2009,20(10):2323-2331. KANG Bing,LIU Shirong,CAI Daoxiong,et al.Effects of Pinus massoniana plantation stand density on understory vegetation and soil properties[J].Chin J Appl Ecol,2009,20(10):2323-2331.
[11] 苏日古嘎,张金屯,王永霞.北京松山自然保护区森林群落物种多样性及其神经网络预测[J].生态学报,2013,33(11):3394-3403. Suriguga,ZHANG Jintun,WANG Yongxia.Species diversity of forest communities and its forecasting by neural network in the Songshan National Nature Reserve,Beijing[J].Acta Ecol Sin,2013,33(11):3394-3403.
[12] 李利平,邢韶华,赵勃,等.北京山区不同区域油松林植物多样性比较研究[J].北京林业大学学报,2005,27(4):12-16. LI Liping,XING Shaohua,ZHAO Bo,et al.Comparative analysis of plant diversity of Pinus tabulaeformis forests in ten regions of Beijing mountainous areas[J].J Beijing For Univ,2005,27(4):12-16.
[13] 林勇明,洪滔,吴承祯,等.桂花次生林群落主要树种种间关联及其对混交度的响应[J].应用与环境生物学报,2007,13(3):327-332. LIN Yongming,HONG Tao,WU Chengzhen,et al.Interspecific association of major plants in Osmanthus fragrans secondary community and its response to mixing[J].Chin J Appl Environ Biol,2007,13(3):327-332.
[14] 郑晓翾,王瑞东,靳甜甜,等.呼伦贝尔草原不同草地利用方式下生物多样性与生物量的关系[J].生态学报,2008,28(11):5392-5400. ZHENG Xiaohuan,WANG Ruidong,JIN Tiantian,et al.Relationships between biodiversity and biomass under different regimes of grassland use in Hulunbeir,Inner Mongolia[J].Acta Ecol Sin,2008,28(11):5392-5400.
[15] 马克平.生物群落多样性的测度方法(Ⅰ)α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168. MA Keping.Measurement of biotic community diversity(Ⅰ)measurement of α diversity[J].Biodiversity Sci,1994,2 (3):162-168.
[16] 马克平,刘玉明.生物群落多样性的测度方法(Ⅰ)α多样性的测度方法(下)[J].生物多样性,1994,2(4):231 -239. MA Keping,LIU Yuming.Measurement of biotic community diversity(Ⅰ)measurement of α diversity[J].Biodiver-sity Science,1994,2(4):231-239.
[17] 胡淑萍,刘鹏举,高开通,等.北京九龙山自然保护区植物群落物种多样性分析[J].水土保持研究,2013,20(4):125-130. HU Shuping,LIU Pengju,GAO Kaitong,et al.Analysis of species diversity of plant communities of Jiulongshan Nature Reserve,Beijing[J].Res Soil Water Conserv,2013,20(4):125-130.
[18] 邓聚龙.灰色系统理论教程[M].武汉:华中理工大学出版社,1990.
[19] 刘新梅,徐润芳,张若勇.邓氏灰色关联分析的应用模型[J].统计与决策,2008(10):23-25. LIU Xinmei,XU Runfang,ZHANG Ruoyong.Application model of Deng’s grey relational analysis [J].Stat Decis, 2008(10):23-25.
[20] 魏天兴,王晶晶.黄土区蔡家川流域河岸林物种多样性研究[J].北京林业大学学报,2009,31(6):49-53. WEI Tianxing,WANG Jingjing.Diversity of riparian vegetation in the Caijiachuan Watershed of the Loess Plateau, southwestern Shanxi Province,northern China[J].J Beijing For Univ,2009,31(6):49-53.
[21] 惠刚盈,赵中华,胡艳波.结构化森林经营技术指南[M].北京:中国林业出版社,2010.
[22] LEGENDRE P,FORTIN M J.Spatial pattern and ecological analysis[J].Vegetatio,1989,80(2):107-138.
[23] 黄忠良,孔国辉,何道泉.鼎湖山植物群落多样性的研究[J].生态学报,2000,20(2):193-197. HUANG Zhongliang,KONG Guohui,HE Daoquan.Plant community diversity in Dinghushan Nature Reserve[J]. Acta Ecol Sin,2000,20(2):193-197.
[24] PALMER M W,WHITE P S.Scale dependence and the species-area relationship[J].Am Nat,1994,144(5):717-740.
[25] 索安宁,巨天珍,张俊华,等.甘肃小陇山锐齿栎群落生物多样性特征分析[J].西北植物学报,2004,24 (10):1877-1881. SUO Anning,JU Tianzhen,ZHANG Junhua,et al.Analysis of biodiversity characteristics of Quercus aliena var.acuteserrata community on Mt.Xiaolong in Gansu[J].Acta Boreal-Occident Sin,2004,24(10):1877-1881.
[26] 侯琳,雷瑞德,康博文,等.黄龙山林区油松林封育过程中植物物种多样性特征[J].西北植物学报,2004,24 (7):1165-1172. HOU Lin,LEI Ruide,KANG Bowen,et al.Traits of plant diversity in a hillclosing and afforestation sequence of Pinus tabulaeformis stands in Huanglong Mountains[J].Acta Boreal-Occident Sin,2004,24(7):1165-1172.
[27] DUAN Renyun,HUANG Minyi,WANG Zhigao,et al.Species diversity of Pseudolarix amabilis population in Yaoluoping Natural Reserve of Anhui Province[J].Adv Mat Res,2012,518:5302-5305.
[28] 卢宝明,邢韶华,崔国发,等.北京山地植物群落的物种多样性比较[J].北京林业大学学报,2010,32(1):36-43. LU Baoming,XING Shaohua,CUI Guofa,et al.Comparison of species diversity of plant communities in mountains of Beijing[J].J Beijing For Univ,2010,32(1):36-43.
[29] 沈年华,万志洲,汤庚国,等.紫金山栓皮栎群落结构及物种多样性[J].浙江林学院学报,2009,26(5):696-700. SHEN Nianhua,WAN Zhizhou,TANG Gengguo,et al.Community structure and species diversity of a Quercus variabilis forest on Mount Zijinshan[J].J Zhejiang For Coll,2009,26(5):696-700.
[30] 徐海,惠刚盈,胡艳波.天然红松阔叶林不同径阶林木的空间分布特征分析[J].林业科学研究,2006,19 (6):687-691. XU Hai,HUI Gangying,HU Yanbo.Analysis of spatial distribution characteristics of trees with different diameter classes in natural Korean pine broad leaved forest[J].Fort Res,2006,19(6):687-691.
Species diversity based on sample size in a Pinus tabulaeformis mixed forest
DENG Hongjian1,LI Weizhong1,CAO Zhu1,WANG Qing1,WANG Guangru2
(1.College of Forestry,Northwest A&F University,Yangling 712100,Shaanxi,China;2.Forest Enterprise of Huanglongshan,Huanglong 716000,Shaanxi,China)
Species diversity is a key indicator of community composition with its spatial pattern and basic causes being important components of species diversity research.To study changing trends in species diversity on various scales,for characteristics of species diversity at all forest layers based on different scales,and for relationships between tree structure and species diversity of undergrowth vegetation,a 1 hm2plot of Pinus tabulaiformis mixed forest on Huanglong Mountain was utilized.Analysis was conducted with Richness,Pielou, Simpson,and Shannon-Wiener Diversity Indexes.Also,the relationship between density and mingling degree of tree layers as well as species diversity indexes of shrub and herb layers were analyzed with a grey relational analysis.Results showed that(1)as the scale increased,the Richness had no clear trend,whereas Simpson, Shannon-Wiener and Pielou Diversity Indexes all stabilized after a certain scale,with their spatial variation decreased.(2)At scales of 5 000 m2and larger,diversity index relationships between varying layers remained unchanged with the shrub layer having the greatest species diversity.The herb layer had the lowest species richness with higher uniformity for species distribution than tree layers.(3)The grey relational analysis showed that the structure of the tree layer was related to species diversity of the shrub and herb layers.The results showed that sampling scale should be taken into account when studying species diversity in a Pinus tabulae-formis mixed forest.[Ch,2 fig.3 tab.30 ref.]
forest ecology;Pinus tabulaeformis forest;species diversity;sampling size;grey relational analysis; mingling degree;Huanglong Mountain
S718.5;Q145
A
2095-0756(2015)01-0067-09
浙 江 农 林 大 学 学 报,2015,32(1):67-75
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.01.010
2014-04-16;
2014-05-15
国家自然科学基金资助项目(31170587)
邓宏兼,从事森林可持续经营与评价研究。E-mail:denghongjian@126.com。通信作者:李卫忠,副教授,博士,从事森林可持续经营与评价研究。E-mail:Wzhli6465@163.com
浙 江 农 林 大 学 学 报,2015,32(1):76-83
Journal of Zhejiang A&F University
doi:10.11833/j.issn.2095-0756.2015.01.011
