胶州湾营养盐限制作用空间差异和长期变化的数值研究*
2015-03-20贾守伟高会旺
贾守伟, 史 洁, 高会旺
(中国海洋大学海洋环境与生态教育部重点实验室,山东 青岛 266100)
胶州湾营养盐限制作用空间差异和长期变化的数值研究*
贾守伟, 史 洁**, 高会旺
(中国海洋大学海洋环境与生态教育部重点实验室,山东 青岛 266100)
以胶州湾为研究区域,建立箱式生态动力学模型,模拟得到胶州湾浮游植物生长受营养盐限制情况的季节变化;通过数值实验分析了胶州湾营养盐限制的区域差异,并进一步比较了1960、1980年代及本世纪初营养盐限制情况的区别。结果表明,胶州湾内的营养盐限制作用有显著的季节变化。春季由于浮游植物旺发消耗大量营养盐,使得湾内营养盐限制春末最强;夏季是河流丰水季,营养盐补充充足,湾内营养盐限制最弱。从不同区域来看,养殖区全年表现为硅限制;河口区和湾口区夏季为磷限制,其他季节为硅限制。营养盐限制作用湾口区最强,养殖区次之,河口区最弱。受此影响,浮游植物生物量河口区最高,湾口区最低。从长期变化来看,1960年代,胶州湾内的营养盐限制作用以氮限制为主,1980年代呈硅和磷交替限制,逐渐演变到本世纪初的以硅限制为主,而且由于营养盐限制作用的逐渐增强,浮游植物生物量也呈现不断下降的趋势。
胶州湾; 营养盐限制; 区域差异; 长期变化
海洋浮游植物是海洋生态系统中的初级生产者和食物链的基础,在海洋生态系统的物质循环和能量转化过程中起着重要作用,其动态变化直接影响到海洋生态系统的功能与结构。浮游植物的生长受到光照、温度和营养盐等多种因素的影响。其中,营养盐水平受到人为活动的影响,关于营养盐对浮游植物生长控制作用的研究有助于污染物减排措施的制定和富营养化的治理,因此得到了国内外学者的广泛关注[1]。若环境中某种营养盐浓度低于浮游植物生长所需的最低阈值,则会产生该种营养盐的限制作用,营养盐限制直接影响海洋的初级生产力和浮游植物群落结构[2-3],从长远来看还可能导致食物链的改变[3]。近岸海域作为从陆地到海洋的过渡区域,受到人类活动的影响最为显著,而且其生产力水平也往往较高。随着气候变化和人类活动的影响,在近岸海域,尤其是半封闭海湾,营养盐各来源的量和比例往往具有差异,因此会导致营养盐限制作用产生显著的时空变化,并对浮游植物的生长造成影响[4-7]。
胶州湾是受人类活动影响显著的半封闭海湾,外与黄海相通,水体平均存留时间为52d[8],其营养盐结构在北方海湾中具有较强的代表性[9]。胶州湾有十余条入海河流,其中位于西北部的大沽河占入海总流量的70%[10],是胶州湾重要的营养盐来源,其带来的营养盐直接影响河口区域。胶州湾内水产养殖业发达,养殖区多集中在东北部,是典型的对虾和蛤蜊养殖区,饵料的过量投放和养殖生物的排泄为该区域输送大量的氮、磷营养盐,使该区域内这两种营养盐浓度远远高于全湾平均水平[11]。胶州湾内的营养盐浓度常年高于外海,通过湾口处与外海的交换,大量营养盐被输送到黄海,因此湾口区域的营养盐结构也有别于其他区域[12]。
现有研究表明,在胶州湾内,相较于光照和水温,营养盐对浮游植物生长的限制作用更为重要[9]。基于观测资料,已有研究阐明了胶州湾内营养盐浓度及浮游植物生物量的季节、长期变化和空间分布特征[12,14-18]。王勇等[19]在胶州湾通过营养盐添加实验发现,网采浮游植物对铁或氮、磷的添加有比较明显的响应。姚云[20]通过分析氮磷比探讨了胶州湾内限制性营养盐的种类及季节变化,认为相较于氮和磷而言,带有硅的评价结果能够更好地反映胶州湾的富营养化状况。观测和实验能够反映胶州湾营养盐结构的现状,但却存在着时间不连续性,并难以区分各个营养盐来源对胶州湾营养盐结构的影响。海洋生态动力学模型是定量研究生态系统中各过程相互作用的有效方法,不仅可以反演现场观测结果,还可以进行机制分析,模拟和预测生态系统的变化。现有的胶州湾生态系统动力学模型研究能够模拟胶州湾营养盐浓度和浮游植物生物量的季节变化,计算得到胶州湾内的营养盐收支状况,但是没有分析不同营养盐来源对胶州湾营养盐限制情况的影响,缺少源的长期变化导致胶州湾生态系统演变的研究[21-24]。
本文选取胶州湾为研究海域(见图1),建立胶州湾箱式生态动力学模型,区分河口区、养殖区和湾口区3个典型区域,模拟并探讨胶州湾内营养盐限制作用的区域差异,分析胶州湾不同营养盐来源对湾内营养盐限制作用以及浮游植物生物量的影响,模拟并分析了胶州湾营养盐结构随营养盐来源变化而产生的长期变化趋势。
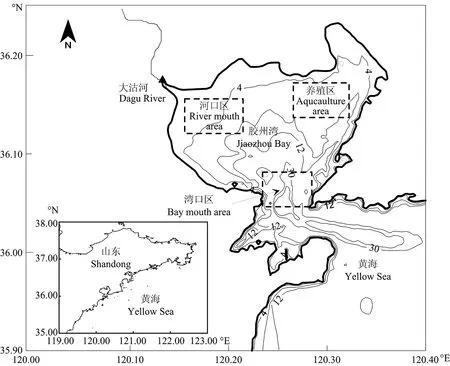
图1 胶州湾地理位置Fig.1 Location of Jiaozhou Bay
1 研究方法
1.1 模型介绍
本文在胶州湾建立了箱式生态系统动力学模型(见图2),来模拟营养盐浓度和浮游植物生物量的季节变化。模型包括无机氮(DIN: dissolved inorganic nitrogen)、无机磷(DIP:dissolved inorganic phosphorus)、硅酸盐(SIL:silicate)3种营养盐(N),浮游植物(P)和碎屑(D)共5个状态变量。浮游植物利用太阳能进行光合作用,摄取营养盐合成有机物,同时呼吸作用释放出营养盐,浮游植物死亡后成为碎屑的一部分,碎屑经过矿化作用向水体补充营养盐。
模型的控制方程如下:
(1)
(2)
(3)

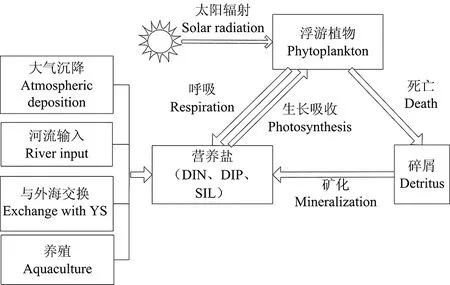
图2 模型概念图Fig.2 Schematic illustration of the model
浮游植物呼吸作用分为光呼吸和基础呼吸过程,并考虑温度对呼吸作用的影响。
(4)
rT=er·(T-T0)
(5)
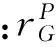
浮游植物的生长受光照条件、温度、营养盐的共同影响,其表达式为:
(6)
其中:g(T)为温度限制因子;VL、VN分别为光和营养盐限制因子。
温度对浮游植物生长的限制采用Q10法则,即:
(7)
光对浮游植物的生长有重要影响,其限制作用的表达函数为:
(8)
其中:Iopt为最优光强;I为水体有效太阳辐射。
本研究中采用Michaeli-Menten(M-M)关系来量化浮游植物生长受到环境中营养盐限制作用的强弱,定义营养盐限制因子为:
(9)
其中:N为营养盐的浓度;KW为浮游植物生长吸收营养盐的半饱和常数。由营养盐限制因子定义可知,在KW一定的情况下,营养盐浓度越高,VN值越接近于1,对浮游植物的限制作用越小。浮游植物生长受到营养盐的限制取决于VN值最小的营养盐,即:
VN=min(VDIN,VDIP,VSIL)
(10)
模型中参数的取值(见表1)采用前人在胶州湾及中国近海的生态模型研究结果[24-26]。

表1 模式中主要参数说明及取值Table 1 Values of parameters used in the model
1.2 模型设置和数据来源
营养盐的源汇项包括河流输入、大气沉降、养殖活动和与黄海交换。其数值大小均采用本世纪初胶州湾的模拟和观测结果(见表2)。入湾河流中大沽河对胶州湾内的营养盐贡献最大,故模型中河流对胶州湾的营养盐贡献仅考虑大沽河一条径流,大沽河的营养盐输入量由流量乘以营养盐的浓度计算得出。大沽河流量采用浦祥[28]模拟得到的2003年大沽河日均径流量结果(见图4)。大沽河营养盐浓度为2002年干、湿季的观测结果[29]。模型计算时将养殖活动和大气沉降一年所贡献的营养盐总量平均分配到各时间步长。养殖活动对胶州湾每年贡献无机氮54×106mol、无机磷3.8×106mol[30]。大气沉降每年为胶州湾贡献无机氮56×106mol,无机磷0.42×106mol,硅酸盐0.93×106mol[31]。考虑胶州湾与黄海之间的营养盐交换,将黄海水中各种营养盐的浓度视为常数,其营养盐通量取决于黄海与胶州湾内营养盐浓度的差值和海水交换率[8, 27]。黄海的营养盐浓度参考已发表的观测资料[12,32-33]。
模型中的物理强迫包括海水温度和海面光强,二者均用余弦函数来拟合,模型采用以下公式进行计算[23]:
Temperature=13.5-11.0×(2π×(t-40)/365)
(11)
Rad=240-140×cos(2π×(t+10)/365)
(12)
其中:Temperature为水温,单位为℃;Rad为海表面的光照强度,单位为W/m2;t的单位为d,t=1代表1月的第1天。
模型采用冬季观测的平均浓度作为初始场,无机氮、无机磷和硅酸盐的浓度分别设为10、0.5、3.5μmol/L,叶绿素浓度为0.8mg/m3[14-15]。开始时间为1月1日,模型时间步长为1h,模型在以上强迫下连续运行3年,用第3年的结果进行分析。
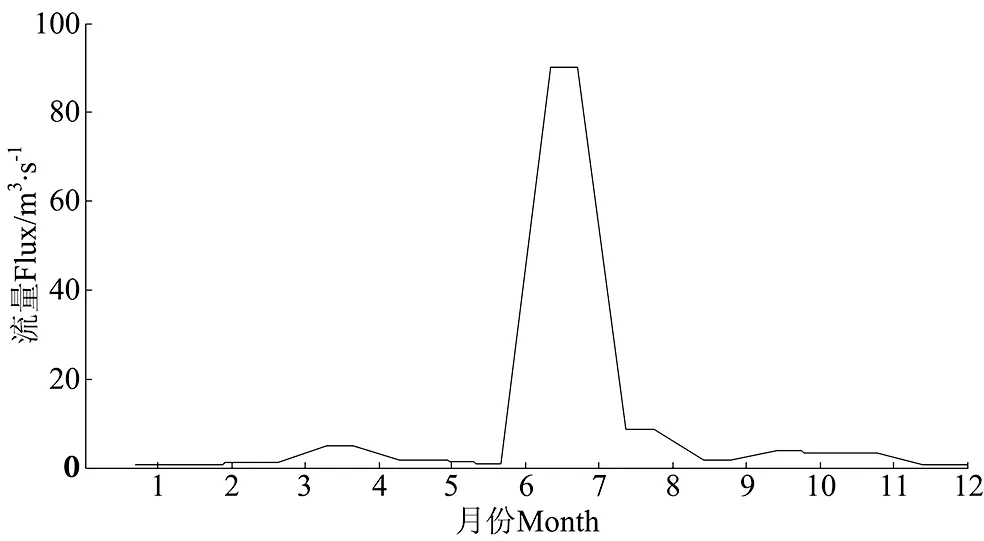
图3 本世纪初大沽河径流量季节变化Fig. 3 Seasonal variations of Dagu River discharge in the beginning of this century
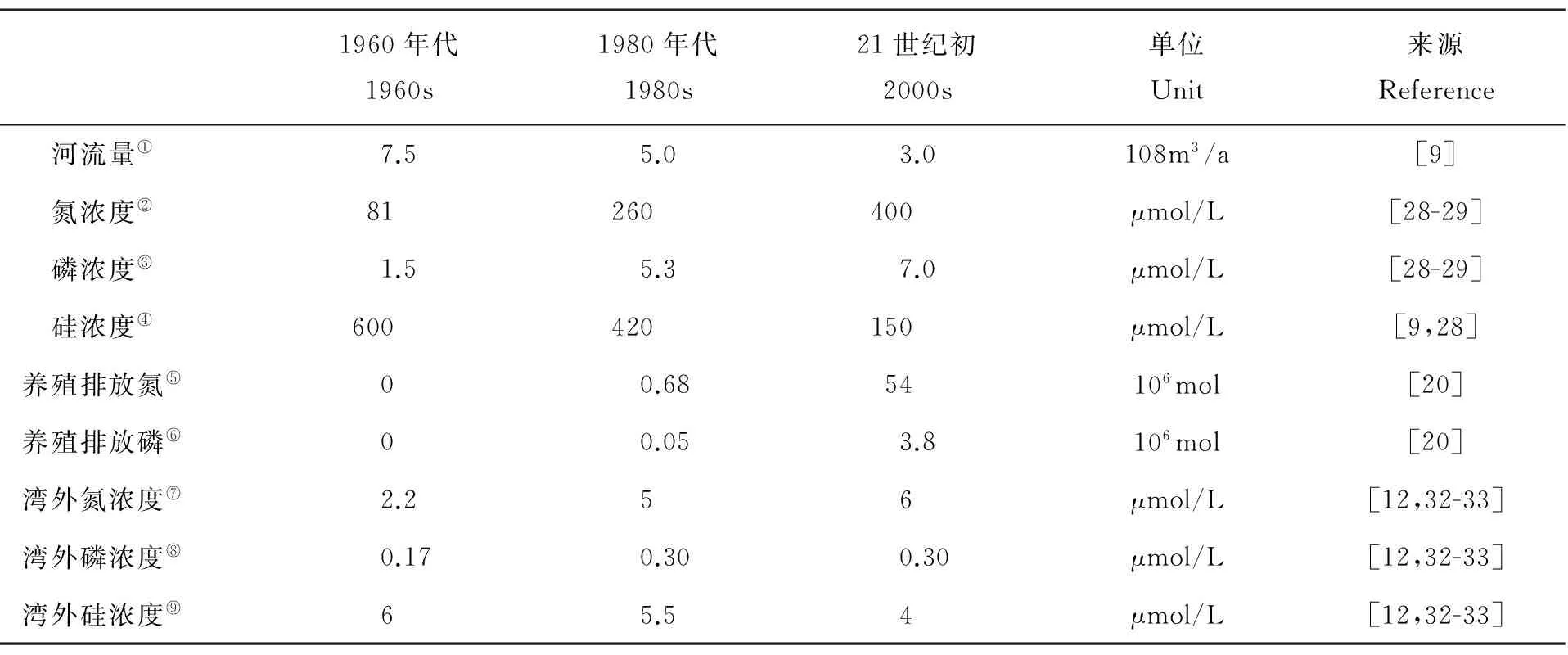
1960年代1960s1980年代1980s21世纪初2000s单位Unit来源Reference河流量①7.55.03.0108m3/a[9]氮浓度②81260400μmol/L[28-29]磷浓度③1.55.37.0μmol/L[28-29]硅浓度④600420150μmol/L[9,28]养殖排放氮⑤00.6854106mol[20]养殖排放磷⑥00.053.8106mol[20]湾外氮浓度⑦2.256μmol/L[12,32-33]湾外磷浓度⑧0.170.300.30μmol/L[12,32-33]湾外硅浓度⑨65.54μmol/L[12,32-33]
Note:①Flow;②Nitrogen concentration;③Phosphorus concentration ;④Silicion concentration;⑤Nitrogen flux of aquaculture;⑥Phosphorus flux of aquaculture;⑦Nitrogen concentration of Yellow Sea;⑧Phosphorus concentration of Yellow Sea;⑨Silicion concentration of YS
2 结果
2.1 敏感性分析
在一个模式中,参数的变化对模拟结果的影响程度反映了模式结果对该参数的敏感程度。由于生物过程本身的非线性特征,生态变量的计算值会对模型中一些参数的变化非常敏感。因此,参数的敏感性分析是生态模型研究中必不可少的步骤。通过敏感性实验,找出生态系统变化敏感参数,为了比较不同参数的敏感程度,定义敏感度为:
(12)
其中:ΔF/F为生态模型中浮游植物生物量的变化率;Δα/α为模型中与生态变量F有关的某个生物参数的变化率。表1提供了10个参数的敏感性数值,浮游植物生物量对最大生长率、浮游植物基础呼吸率、最优光强等参数较敏感,而对矿化率、氮营养盐半饱和常数等参数相对不敏感。各参数分别变化或减小相同比例,其敏感度并不相同,表明各参数的变化是非线性变化,如浮游植物死亡率,增大50%其敏感度为0.65,减小50%其敏感度为1.91。75%的参数敏感度小于0.5,因此本文建立的模型是稳定的。
2.2 本世纪初营养盐浓度和浮游植物生物量的季节变化
应用所建立的胶州湾箱式生态动力学模型,模拟本世纪初的胶州湾营养盐浓度和浮游植物生物量的季节变化规律(见图4)。
胶州湾内3种营养盐的浓度均为夏季最高,春季最低。春季,由于浮游植物生长摄取大量营养盐,导致胶州湾内营养盐浓度一直维持在较低水平。夏季,大沽河进入汛期,河流输入带来的营养盐显著增加,使湾内各营养盐浓度迅速升高,到8月中旬达到峰值,无机氮、无机磷和硅酸盐的浓度峰值分别为41.6、1.0和10.3 μmol/L,出现时间均在第220天左右。丰水期之后,浮游植物生长消耗大量的营养盐,使其浓度不断下降。胶州湾内浮游植物生物量的季节变化特征明显,模拟结果显示呈单峰状,叶绿素年平均浓度为1.4mg/m3。冬季,太阳辐射较弱,水温较低,不利于浮游植物的生长,叶绿素浓度比较低。夏季,营养盐充足,温度和光照条件适宜,浮游植物大量繁殖,叶绿素浓度达到峰值4.1mg/m3,稍微滞后于营养盐浓度峰值。此后,由于营养盐大量消耗及太阳辐射的限制作用不断增强,抑制了浮游植物的生长,叶绿素浓度降低,冬季达到最低。模拟得到的本世纪初胶州湾营养盐浓度和浮游植物生物量的季节变化规律与观测结果较为吻合。现场观测发现某些年份浮游植物生物量在2月也存在峰值,有学者[14, 34]推测,这是由于某些冷水种藻类冬季大量生长造成的,但并未证实。在本模型中温度对浮游植物生长的限制采用Q10法则,因此难以体现冷水种浮游植物在冬季的旺发。
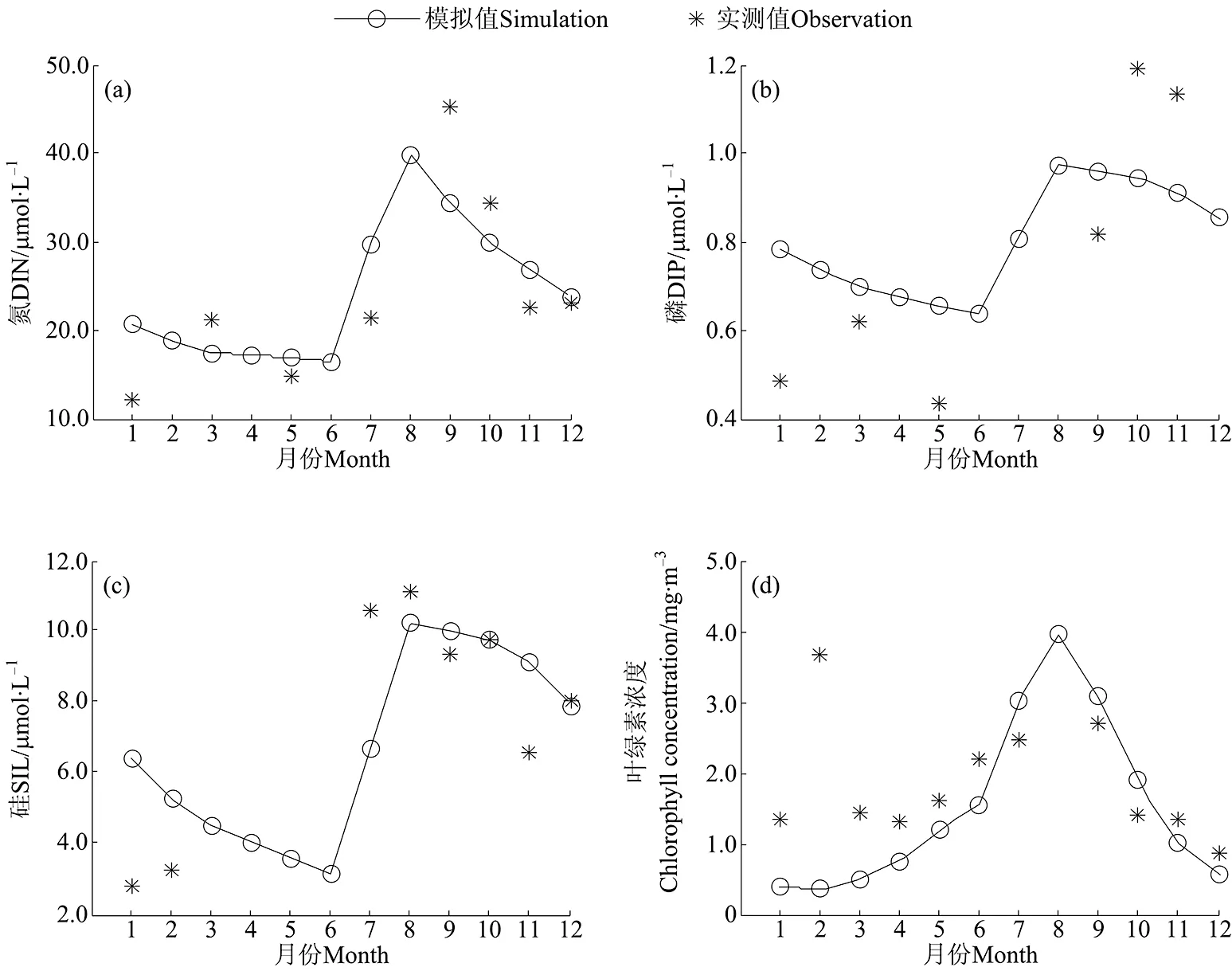
图4 营养盐浓度和浮游植物生物量的季节变化
2.3 本世纪初营养盐限制的季节变化
3种营养盐对浮游植物的限制作用存在强弱差异,因此探讨营养盐限制作用的季节变化对于理解营养盐结构对浮游植物生长的影响有重要作用。通过比较模拟结果中3种营养盐限制因子的数值大小,来确定限制浮游植物生长的营养盐种类,即营养盐限制因子数值最小的被认为是限制浮游植物生长的营养盐。
比较3种营养盐限制因子可以看出(见图5),3种营养盐限制因子的数值均表现为春季最小,夏季最大,这说明胶州湾内的营养盐限制作用夏季最弱,秋季和冬季次之,春季限制作用最强。夏季,大沽河径流量达到全年最大,带来充足的营养盐,削弱了营养盐限制作用。秋季,随着径流输入的减少,加上浮游植物的生长不断消耗营养盐,营养盐限制作用逐渐增强。经过冬、春季节的消耗,营养盐限制作用在春末夏初(180 d左右)达到最强。从数值上看,氮和磷限制因子的数值全年均在0.8以上,而硅限制因子常年小于氮、磷,仅在夏季营养盐丰富时能够达到0.75左右,由于春季浮游植物旺发的大量吸收,5~6月份期间硅限制因子达到最小值,约为0.5,即浮游植物在硅酸盐的限制作用下,其生长速率只能达到最大生长率的一半。由此可见,硅是限制胶州湾浮游植物生长的首要营养盐,而氮和磷相对充足,不限制浮游植物的生长。
胶州湾是位于黄海边缘的一个海湾,但与黄海中部海域相比,其营养盐限制作用却有其特性。黄海中部海域,营养盐限制作用夏季最强,冬季最弱,且主要受磷的潜在限制。王保栋等[36]观测发现,黄海大部分海域硅氮比值大于1,可以认为硅酸盐不会限制浮游植物的生长。Zhang[37]研究发现,黄河、滦河、鸭绿江、大辽河等几条黄海主要入海河流的氮磷比值在20~5000之间,远远高于Redfield系数。因此,黄海海域水体中无机氮相对过剩,而无机磷相对不足,黄海中部海域浮游植物的生长明显受磷限制。从季节上看,黄海中部水域中营养盐限制最强和最弱分别出现在夏季和冬季,而不像胶州湾分别出现在春季和夏季。黄海春季出现较强的浮游植物水华现象,大量消耗营养盐。夏季,随着海表面温度升高,形成较强的跃层,较弱的垂直混合阻碍了底部高营养盐的水向上补充到真光层,进而使得此时营养盐限制达到全年最强。秋季水华过后,大风混合增强使得上层得到营养盐的补充,并且温度降低和太阳辐射减弱,浮游植物生长速率减慢。到了冬季,低浮游植物生物量和高营养盐浓度,使得此时营养盐限制为全年最弱。黄海中部水域营养盐限制的季节变化是受到浮游植物生长和跃层生消的共同作用,而胶州湾水深较浅,跨跃层补充的季节变化并不显著,因此营养盐限制的季节变化主要取决于浮游植物的生长和河流的丰水、枯水期变化。可见,不同的营养盐补充机制,会导致不同的海域,即使是相互连通的海域,其营养盐限制情况也会呈现不同的季节变化规律。
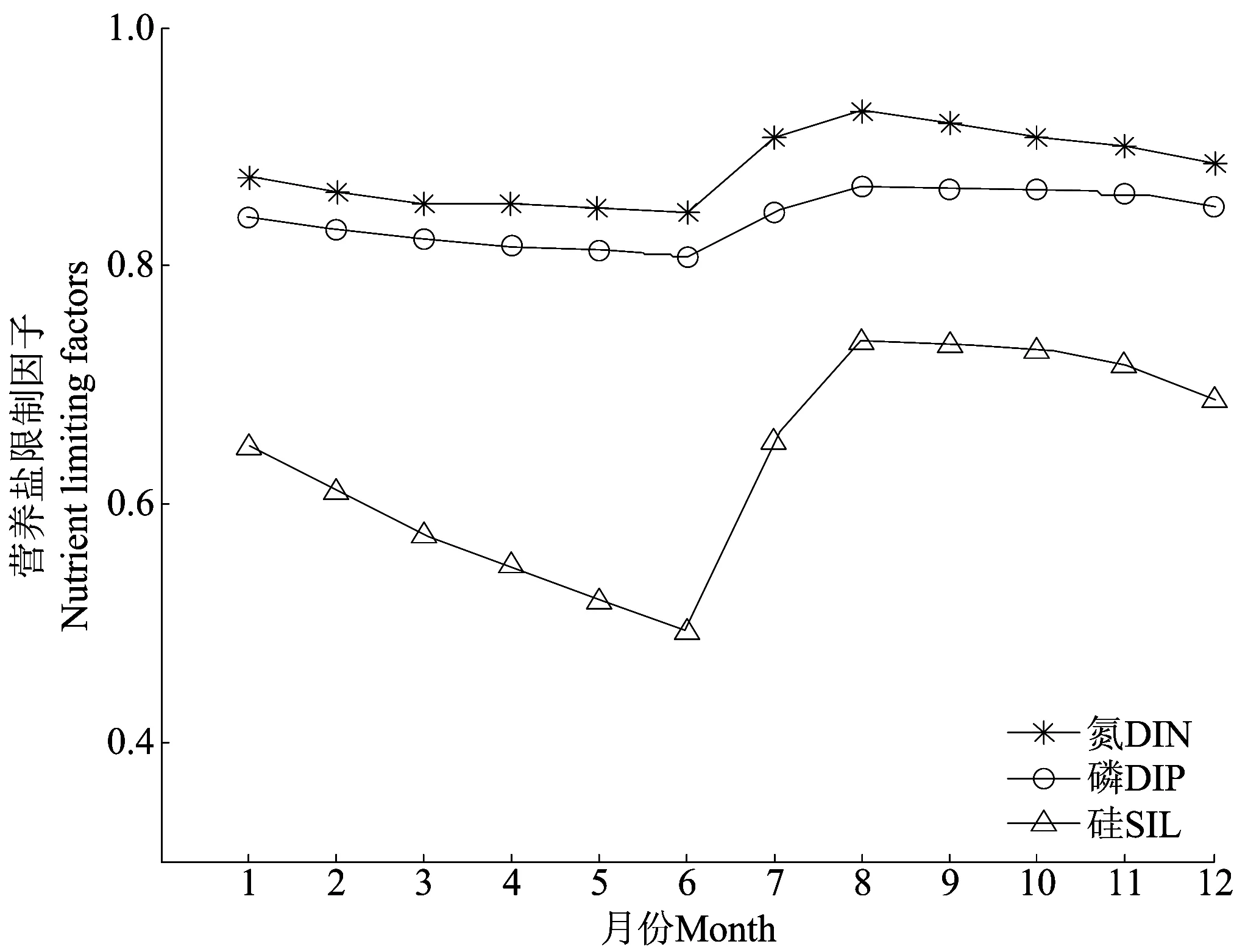
图5 营养盐限制因子的季节变化Fig.5 Seasonal variations of nutrient limiting factors
3 讨论
3.1 营养盐限制的区域差异
胶州湾内不同区域营养盐的来源不同,并且各个营养盐来源的量和比例也有较大区别,因此营养盐浓度及结构都存在着空间不均匀性,浮游植物生长受到的营养盐限制作用也会存在较大的区域差异。
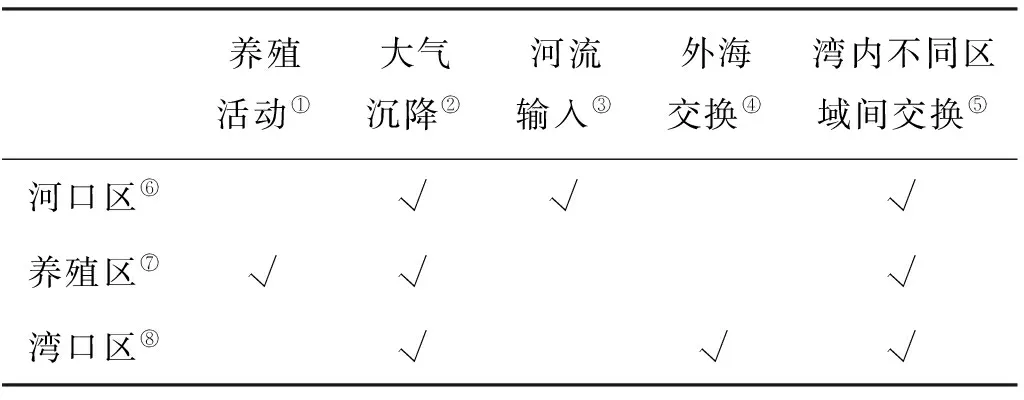
表3 不同区域的营养盐来源Table 3 Nutrient sources in different areas
Note:①Aquaculture;②Atmospheric deposition;③River input;④Exchange with YS;⑤Exchange among the different areas;⑥River mouth area;⑦Aquiculture area;⑧Bay mouth area
分别将河口区、养殖区和湾口区作为研究区域(见图1),考虑不同区域营养盐的来源差异(见表3),将生态模型应用于3个区域,其中湾内不同区域间营养盐的交换量取决于该区域与全湾水平的营养盐浓度差,模拟得到各区域营养盐限制因子的季节变化(见图6)。氮限制因子的数值在不同区域均大于0.8,始终是3种营养盐限制因子中最大的,说明3个区域都没有表现出氮限制,胶州湾内无机氮的现存量相对于浮游植物的生长需要来说过剩。河口区硅和磷限制因子的数值区间均为0.6~0.8,在3~7月份,硅限制因子小于磷限制因子,而在8~12月份,磷限制因子小于硅限制因子。在河口区由硅和磷交替限制浮游植物的生长。冬、春季,大沽河处于枯水期,硅酸盐的来源不足,浮游植物的生长主要受到硅的限制;夏季,径流急剧增加带来丰富的营养盐,而且硅酸盐的浓度在汛期为枯水期的12.3倍,无机磷仅为枯水期的7倍[29],大量的硅酸盐被输送到胶州湾,而无机磷相对不足,使得营养盐限制由硅限制转变为磷限制。在养殖区,硅限制因子的数值全年均在0.7以下,远小于另外两种营养盐,说明该区域呈显著的硅限制。养殖活动为该区域贡献大量氮和磷,而硅酸盐无充足来源,此外,养殖区位于胶州湾的东北角,潮流作用较弱,与其他区域的水交换相对缓慢,加剧了无机氮、无机磷的富集,阻碍了其他区域向此处硅酸盐的补充。从全年来看,硅对浮游植物的限制作用春末尤为突出,浮游植物的生长使硅酸盐几乎完全被消耗。湾口区,由于与外海水交换顺畅,大量营养盐被输运到外海,此处营养盐浓度低于全湾平均水平,无机磷和硅酸盐交替成为限制因子,春季为硅酸盐限制,到了秋季转为无机磷限制。取3种营养盐限制因子中的最小值(见公式10),绘制不同区域该值的全年变化曲线,可以发现湾口区的数值最小,全年均在0.6以下,而河口区和养殖区多大于0.6,其中以河口区的数值最大,因此胶州湾内河口区的营养盐限制作用最弱,养殖区次之,湾口区限制作用最强。
通过模型结果可以看出(见图7),各区域的浮游植物生物量季节变化均呈单峰状,峰值出现在夏季,但生物量存在明显差异。营养盐限制作用较弱的河口区,浮游植物生长旺盛,浮游植物生物量最大,叶绿素年平均浓度为1.7mg/m3,高出全湾平均水平20%。反之,营养盐限制作用较强的湾口区,其浮游植物生物量则大大低于平均水平,年平均浓度仅为1.1 mg/m3,比全湾平均值低21%。养殖区浮游植物生物量介于其他两区域之间。吴玉霖等[34]观测也发现,胶州湾内叶绿素高值区多集中在西北部水域,而该水域也是大沽河影响较大的区域。可见,营养盐限制作用的区域差异,最终导致不同区域间浮游植物生物量及初级生产力也有较大差别。
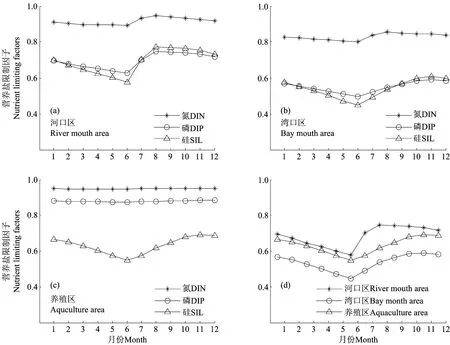
图6 不同区域营养盐限制因子的季节变化

图7 不同区域浮游植物生物量的季节变化
3.2 营养盐限制的年际差异
近40年来,胶州湾内的浮游植物生物量不断下降调查表明,胶州湾内的海水温度和盐度在此期间没有大的变化,而且大部分水域的水质和沉积环境质量较好[35]。因此,胶州湾内浮游植物生物量的减少,可能与胶州湾内的营养盐结构变化密切相关。表2给出了不同时期河流输入、养殖排放和外海交换对胶州湾营养盐的贡献,光照和水温仍然用余弦函数拟合,应用所建立的箱式生态动力学模型,分别模拟得到20世纪60年代、80年代营养盐浓度及限制因子的季节变化,结合已本世纪初的结果,比较分析40年来胶州湾营养盐限制情况的长期变化特征及其与浮游植物生物量的关系。
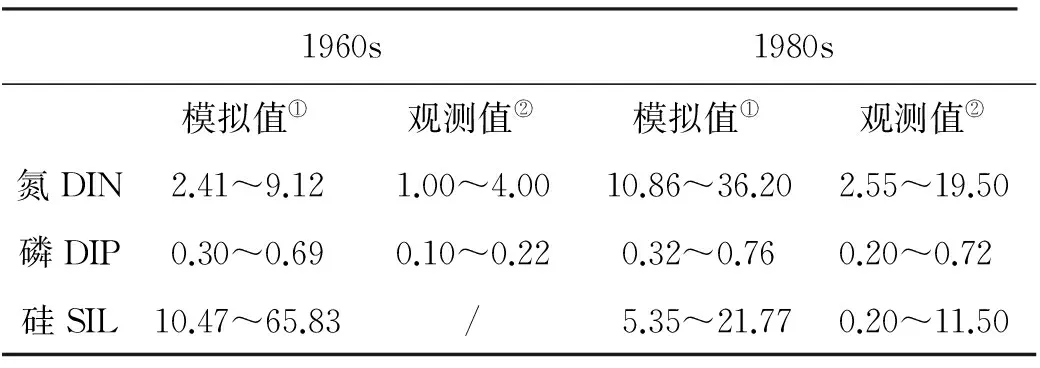
表4 营养盐浓度长期变化模拟结果与观测结果对比[35]Table 4 Comparison between the simulation and observation in the 1960s and the 1980s
Note:①Simulation;②Observation
从长期变化模拟结果来看,胶州湾营养盐浓度和限制作用变化显著(见表4;图8)。20世纪60年代,氮限制因子的数值在3种营养盐中最小,最小值约为0.7,磷限制因子略大于氮,硅的限制因子最大,其数值在夏季达到0.95,说明该时期胶州湾内浮游植物的生长主要受到氮的限制,其次为磷,硅的限制作用最小。这是因为在20世纪60年代,胶州湾内无机氮浓度相对较低,全年的氮磷比平均值仅为2.8,浮游植物的生长消耗大量的营养盐,海水中的氮常常被耗尽,特别是在生长高峰期,很多区域的站位检测不到无机氮[35]。20世纪80年代,硅限制因子的数值较小(0.6~0.8),仅在夏季磷限制因子的数值略小于硅,说明全年内浮游植物的生长受硅限制较强,仅在夏季受磷限制。这是由于1980年代人类活动排放导致胶州湾内无机氮的浓度迅速升高,氮限制作用大幅减弱,不再成为限制因子,而无机磷的来源相对较少,增速慢于无机氮。以胶州湾中东部水域为例,1960~1980年代,无机氮增加7.2倍,而无机磷仅增加2.2倍。与此同时,由于河流上游的水利工程,大沽河的径流量和输沙率呈下降的趋势,大量泥沙被截留。1960年代以前,大沽河年输沙量平均为170.03万t,到1970年代末仅为71.21万t,减少59%,导致硅酸盐浓度显著降低,成为限制浮游植物的主要因子,仅在汛期,大沽河的径流输入带来大量的硅酸盐,此时无机磷相对不足,呈现磷限制[33]。本世纪初,氮和磷的限制作用均大大减弱,浮游植物的生长受硅限制作用明显,近年来大沽河输沙量减少为36.59万t,仅为1960年代以前的22%,导致胶州湾内硅营养盐浓度的不断降低,进而成为浮游植物生物量的主要限制因子。
比较不同时代营养盐综合限制因子的数值可以发现,近40年来该值逐渐减小,说明胶州湾内的营养盐限制作用有不断增强的趋势。图9为模拟得到的1960年代、1980年代和本世纪初的胶州湾浮游植物生物量的季节变化。相对于1960年代,1980年代略有降低,而21世纪初浮游植物生物量大约减少23%。由于本模型只考虑一种浮游植物,只能体现浮游植物生物量的变化,无法反映浮游植物群落结构的变化。但是,自1960年代以来,随着无机氮和无机磷输入的增加以及硅酸盐输入的减少,限制性营养盐发生转变,可以推测胶州湾内的大型浮游植物植物不断减少,而小型浮游植物生物量逐渐增加,并导致总体浮游植物生物量显著减少。
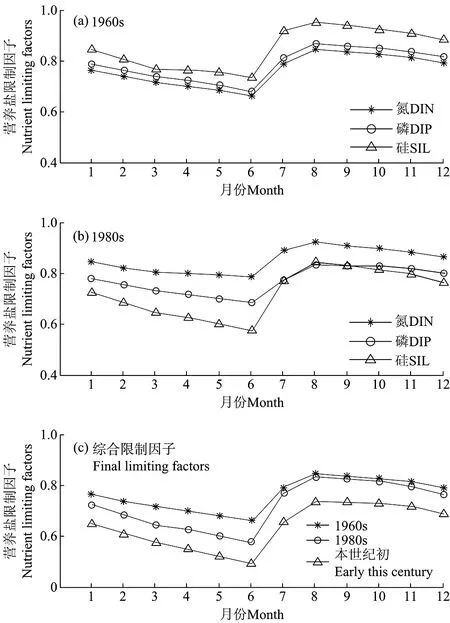
图8 营养盐限制因子的季节变化
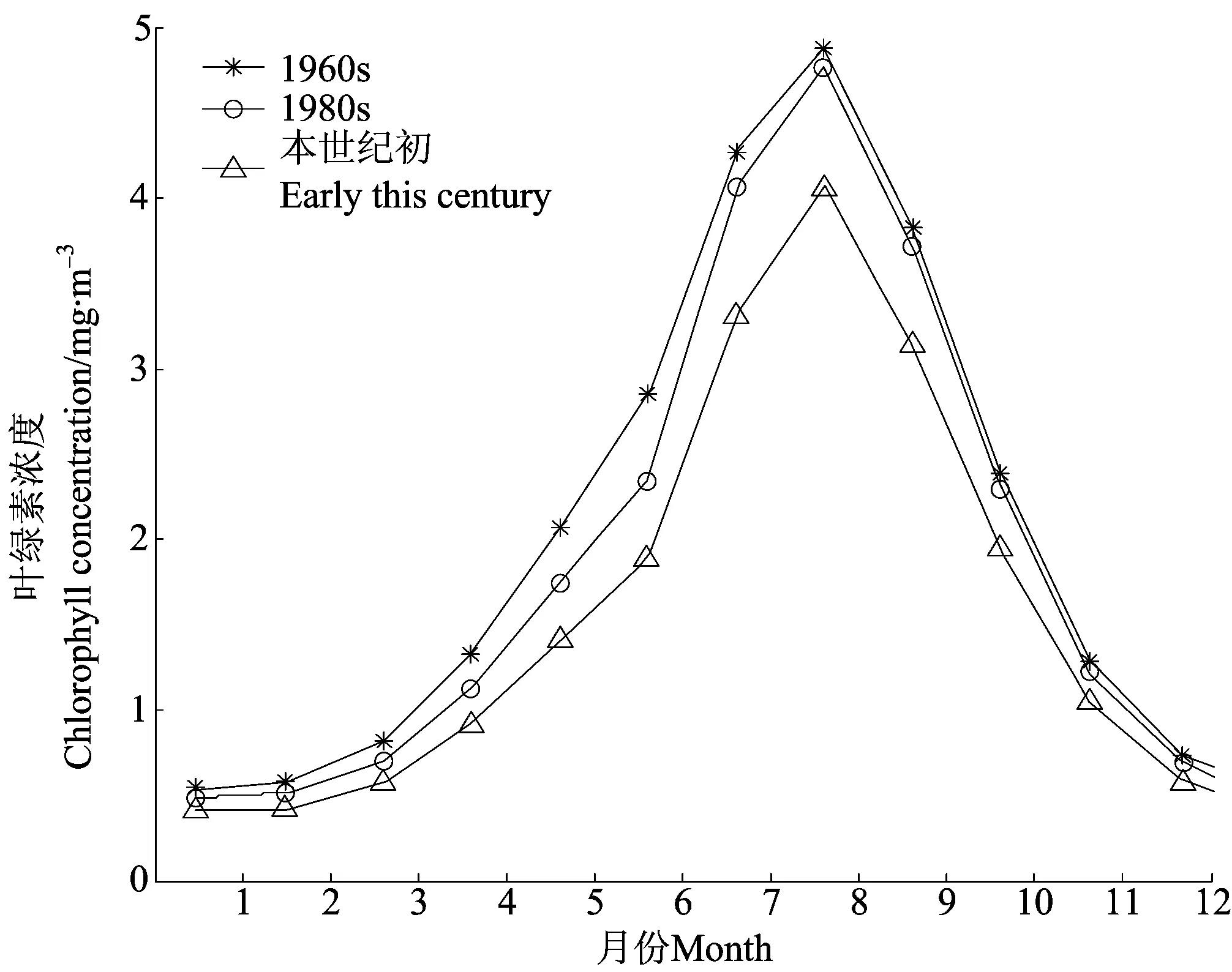
图9 不同年代浮游植物生物量的季节变化特征
4 结语
本研究建立胶州湾箱式生态动力学模型,模拟胶州湾内不同区域及不同年代营养盐限制浮游植物生长情况的差异。胶州湾内营养盐限制作用有显著的季节变化,春季最强,夏季最弱,并存在区域差异,河口区和湾口区的浮游植物生长受硅和磷的交替限制,养殖区呈现显著的硅限制,进而导致浮游植物生物量的空间分布不同。河口区的营养盐限制作用最弱,浮游植物生物量最高,养殖区次之,而湾口区营养盐限制作用最强,浮游植物生物量低于全湾平均水平。
从时间演变上看,1960年代,胶州湾内营养盐限制作用以氮限制为主;1980年代,磷和硅交替限制浮游植物的生长,夏季为磷限制,其他季节为硅限制;本世纪全年呈现显著的硅限制。近40年来,营养盐限制作用有逐渐增强的趋势,受此影响,胶州湾内的浮游植物生物量也不断下降,相对20世纪60年代,21世纪初浮游植物生物量降低约23%。营养盐限制作用的转变可能会导致藻类优势种的变化,并通过食物链向上层营养级传递,最终会影响胶州湾内海洋生态结构和渔业生产。
在近岸海域,尤其是胶州湾这样的半封闭海湾,其营养盐限制作用会随着营养盐来源的变化而改变,并会对生态系统产生影响。胶州湾与世界上其他类似海域相比,有其独特性,但也存在许多共性。Danielsson等[5]研究发现,在波罗的海内,由于不同区域营养盐来源的差异,营养盐限制作用也存在显著的区域差异,其中的芬兰湾,其营养盐来源与胶州湾相似,内有较大河流的输入,营养盐限制作用在夏季、秋初表现为氮限制,而其余时间均处于硅限制,这也与胶州湾营养盐限制作用的季节演替规律相似。而在没有河流来源的波的尼亚湾,全年均呈现显著的磷限制,没有季节更替。从长期来看,波罗的海有逐步向硅限制转化的潜在趋势,而且这种转化已经其浮游植物群落结构和初级生产力产生影响。同样,在地中海的河口区域,长期以来氮和磷增加显著,而硅的变化较小,也促进了该水域有害鞭毛藻的爆发[7],而胶州湾的长期变化趋势也与之类似,大量氮、磷的增加以及硅的减少,导致胶州湾内硅藻等大型藻类正在减少,取而代之的是不需要吸收硅的甲藻和鞭毛藻[18]。现阶段我们的模型仅能模拟出营养盐组成及浮游植物生物量的变化特征,下一步工作中,作者将在模型中增加不同浮游植物类型,综合讨论人类活动和气候变化影响下营养盐和浮游植物群落结构的演变。
[1] Wu J T, Chou, T L. Silicate as the limiting nutrient for phytoplankton in a subtropical eutrophic estuary of Taiwan [J]. Estuarine Coastal and Shelf Science, 2003, 58: 155-162.
[2] Redfield A, Ketchum B, Richards F. 1963. The influence of organisms on the composition of seawater [M]. //Hill M, ed. The Sea. New York: Wiley, 1963: 26-77.
[3] Tyrerell T. The relative influences of nitrogen and phosphorus on oceanic primary production [J]. Nature, 1999, 400(6744): 525-531.
[4] Wan Z, Bi H, She J, et al. Model study on horizontal variability of nutrient N/P ratio in the Baltic Sea and its impacts on primary production, nitrogen fixation and nutrient limitation [J]. Ocean Science Discussions, 2012(9): 385-419.
[5] Danielsson Å, Papush L, Rahm L. Alterations in nutrient limitations—Scenarios of a changing Baltic Sea [J]. Journal of Marine Systems, 2008, 73(3): 263-283.
[6] Fisher T, Hagy J, Boynton W, et al. Cultural eutrophication in the Choptank and Patuxent estuaries of Chesapeake Bay [J]. Limnology and Oceanography, 2006, 51(1): 435-447.
[7] Domingues R, Anselmo T, Barbosa A, et al. Nutrient limitation of phytoplankton growth in the freshwater tidal zone of a turbid, Mediterranean estuary [J]. Estuarine, Coastal and Shelf Science, 2010, 91(2): 282-297.
[8] Liu Z, Wei H, Liu G L, et al. Simulation of water exchange in Jiaozhou Bay by average residence time approach [J]. Estuarine, Coastal and shelf Science, 2004, 61(1): 5-35.
[9] 张哲, 王江涛. 胶州湾营养盐研究概述 [J]. 海洋科学, 2009, 33(11): 90-94.
[10] 李莉, 梁生康, 石晓勇, 等. 2007年胶州湾入海河流污染状况和污染物入海通量分析 [J]. 环境科学与管理, 2009, 34(6): 23-28.
[11] 宋秀贤, 俞志明. 胶州湾东北部养殖海域夏季营养盐分布特征及其对浮游植物生长的影响 [J]. 海洋与湖沼, 2007, 38(5): 446-452.
[12] 任玲, 张曼平, 孙军, 等. 胶州湾内外水体部分化学生物因子调查分析 [J]. 青岛海洋大学学报: 自然科学版, 2003, 33(4): 557-564.
[13] Yang D F, Gao Z H, Sun P Y, et al. Mechanism of nutrient silicon and water temperature influences on phytoplankton growth [J]. Marine Science Bulletin, 2006, 8(2): 49-59.
[14] 李艳, 李瑞香, 王宗灵, 等. 胶州湾浮游植物群落结构及其变化的初步研究 [J]. 海洋科学进展, 2005, 23(3): 328-334.
[15] 李超伦, 张芳, 申欣, 等. 胶州湾叶绿素的浓度、分布特征及周年变化 [J]. 海洋与湖沼, 2005, 36(6): 499-506.
[16] 孙晓霞, 孙松, 赵增霞, 等. 胶州湾营养盐浓度与结构的长期变化 [J]. 海洋与湖沼, 2011, 42(5): 662-669.
[17] 孙晓霞, 孙松, 张永山, 等. 胶州湾叶绿素a及初级生产力的长期变化 [J]. 海洋与湖沼, 2011, 42(5): 654-661.
[18] Shen Z L. Historical changes in nutrient structure and its influences on phytoplantkon composition in Jiaozhou Bay [J]. Estuarine, Coastal and Shelf Science, 2001, 52(2): 211-224.
[19] 王勇, 赵澎, 单宝田. 胶州湾营养盐限制浮游植物生长的初步模拟现场实验研究 [J]. 海洋科学, 2002, 26(10): 55 -58.
[20] 姚云, 郑世清, 沈志良. 胶州湾营养盐及富营养化特征 [J]. 海洋通报, 2007, 26(4): 91-98.
[21] 李克强, 王修林, 石晓勇, 等. 胶州湾围隔浮游生态系统氮、磷营养盐迁移-转化模型研究 [J]. 海洋学报, 2007, 29(3): 76-83.
[22] Han H Y, Li K Q, Wang X L, et al. Environmental capacity of nitrogen and phosphorus pollutions in Jiaozhou Bay, China: Modeling and assessing [J]. Marine Pollution Bulletin, 2011, 63(5): 262-266.
[23] Ren L, Zhang M P, Brockmann, U. H, et al. Pelagic Nitrogen Cycling in Jiaozhou Bay, a model study Ⅰ: The conceptual model [J]. Chinese Journal of Oceanology and Limnology, 2003b, 21, 358-367.
[24] Liu Z, Wei H, Bai J, et al. Nutrients seasonal variation and budget in Jiaozhou Bay, China: a 3-dimensional physical-biological coupled model study [J]. Water Air & Soil Pollution: Focus, 2007, 7(6): 607-623.
[25] 王震勇. 胶州湾浮游生态系统四十年变化的模拟与分析 [D]. 青岛: 中国海洋大学, 2007.
[26] 张燕, 孙英兰, 袁道伟, 等. 胶州湾氮、磷浓度的三维数值模拟 [J]. 中国海洋大学学报: 自然科学版, 2007, 31(1): 021-026.
[27] 赵亮, 魏皓. 胶州湾水交换的数值研究 [J]. 海洋与湖沼, 2002, 33(1): 23-29.
[28] 浦祥. 大沽河流域非点源氮输出及其对胶州湾水质影响的数值研究 [D]. 青岛: 中国海洋大学, 2011.
[29] Liu S M, Zhang J, Chen H T, et al. Factors influencing nutrient dynamics in the eutrophic Jiaozhou Bay, North China [J]. Progress in Oceanography, 2005, 66(1): 66-85.
[30] 王刚. 胶州湾入海点源、海水养殖污染物通量研究 [D]. 青岛: 中国海洋大学, 2009.
[31] Liu S M, Li X N, Zhang J, et al. Nutrient dynamics in Jiaozhou Bay [J]. Water Air Soil Pollution: Focus, 2007, 7(6): 625-643.
[32] 田恬, 魏皓, 苏健, 等. 黄海氮磷营养盐的循环和收支研究 [J]. 海洋科学进展, 2003, 21(1): 1-11.
[33] 王保栋. 黄海和东海营养盐分布及其对浮游植物的限制 [J]. 青岛海洋大学学报: 自然科学版, 2003, 14(7): 1122-1126.
[34] 吴玉霖, 孙松, 张永山, 等. 胶州湾浮游植物数量长期动态变化的研究 [J]. 海洋与湖沼, 2004, 35(6): 518-523.
[35] 沈志良. 胶州湾营养盐结构的长期变化及其对生态环境的影响 [J]. 海洋与湖沼, 2002, 33(3): 322-331.
[36] 王保栋. 黄海和东海营养盐分布及其对浮游植物的限制 [J]. 青岛海洋大学学报: 自然科学版, 2003, 14(7): 1122-1126.
[37] Zhang J. Nutrient elements in large Chinese estuaries [J]. Continental Shelf Research, 1996, 16(8): 1023-1045.
责任编辑 庞 旻
Numerical Study on the Regional and Long-Term Variation of Nutrient Limitation in Jiaozhou Bay
JIA Shou-Wei, SHI Jie, GAO Hui-Wang
(The Key Laboratory of Marine Environment and Ecology,Ministry of Education,Ocean University of China,Qingdao 266100,China)
A box ecosystem model is developed to simulate the seasonal variations of nutrient limitation in the Jiaozhou Bay.After model validation, the regional and interannual variations of nutrient limitation are analyzed. The results show that the nutrient limitation exhibits obvious seasonal variation. In spring, the phytoplankton grows fast and exhausts the available nutrients. The nutrient limitation becomes the strongest in late spring and turns to be the weakest in summer when the river in flood season supply sufficient nutrients into the Jiaozhou Bay. Meanwhile, the regional variations of nutrient limitation are obvious. The limitation nutrient is SIL in aquaculture area, and is SIL and DIP alternatively in both the river mouth area and the mouth of the Bay. The nutrient limitation is the strongest in the mouth area of the Bay, while the weakest in the river mouth area. In 1960s, the main limiting nutrient is DIN. Under the influence of human activity and climate change, the limiting nutrient has altered from DIN to SIL until now. The DIP limitation occurs in the certain region and period. The nutrient limitation is continually enhanced as time goes on. As a result, the phytoplankton biomass at the beginning of this country decreases by 21% comparing with that in 1960s.
the Jiaozhou Bay; nutrient limitation; regional variation; interannual variation
国家自然科学基金青年基金项目(41106007);国家自然科学基金委员会—山东省人民政府联合资助海洋科学研究中心项目(N1406403)资助
2014-01-17;
2014-02-27
贾守伟(1987-),男,硕士生。
** 通讯作者: E-mail:shijie@ouc.edu.cn
P735
A
1672-5174(2015)05-001-10
10.16441/j.cnki.hdxb.20140024
