光肩星天牛成虫4种活动行为日节律1)
2015-02-08吕飞海小霞王志刚阎爱华毕拥国刘炳响
吕飞 海小霞 王志刚 阎爱华 毕拥国 刘炳响
(河北省林木种质资源与森林保护重点实验室(河北农业大学),保定,071000)
光肩星天牛成虫4种活动行为日节律1)
吕飞 海小霞 王志刚 阎爱华 毕拥国 刘炳响
(河北省林木种质资源与森林保护重点实验室(河北农业大学),保定,071000)
采用室内雌雄配对隔离饲养的方法,对光肩星天牛(Anoplophoraglabripennis)取食、交尾、移动、静息4种行为进行观察。结果表明:①各活动行为的高峰期不同。取食行为发生在21:30(49%)、交尾行为在12:00(34%)、移动行为在10:30(36.5%)、静息行为在8:00—9:00(36%~41%)。②活动行为在发生时段上存在显著差异。取食和静息行为黑夜高于白天,交尾、移动行为白天高于黑夜。凌晨、上午、中午、下午、黄昏、晚上6个时间段,交尾、移动和静息行为存在极显著性差异(p<0.01),取食行为存在显著性差异(p<0.05)。③性别对各行为日节律有一定影响,雌雄虫发生各行为的高峰期存在差异。白天和黑夜,雌雄虫各行为事件的发生比例均无显著性差异(p>0.05);6个时间段,雌雄虫中午的取食行为、上午和中午的移动行为呈显著性差异、凌晨移动行为呈极显著性差异,其它时间段无显著性差异。综上可见,光肩星天牛成虫活动行为有明显的时间节律;性别对其活动行为节律有一定影响。
光肩星天牛;成虫;性别差异;昆虫行为;日节律
光肩星天牛(Anoplophoraglabripennis)属鞘翅目(Coleoptera)、天牛科(Cerambycidae)、沟胫天牛亚科(Lamiinae)、星天牛属(Anoplophora),是重要的林木蛀干害虫。广泛分布于我国黑龙江、宁夏、西藏、贵州等地,国外在亚洲、北美、欧洲均有分布[1]。成虫补充营养啃食幼嫩树枝、树叶主脉;幼虫蛀食树干为主[1]。主要寄主有槭属(Acer)、杨属(Populus)、柳属(Salicaceae)、榆属(Ulmus)等多种阔叶树[2-5]。20世纪末给我国“三北防护林”带来了毁灭性的危害[6-7]。在国外,至1996年首次报道在美国纽约发现光肩星天牛之后,在芝加哥、伊利诺斯、新泽西州、马萨诸塞州等均发现该虫踪迹[1,8];从1997到2008年,美国为了根除光肩星天牛的危害,8 543棵被侵染树、33 593棵高风险树被砍掉,共花费约3.73亿美元[1]。在欧洲,2001年报道奥地利发现该虫之后,法国、德国、意大利等地对该虫的入侵情况进行了相继报道[8-9];从2001年到2008年,欧洲为了根除该虫花费约55万欧元[1]。该虫给各国不仅造成了严重的经济损失,而且也给生态环境带来了严重的威胁[10]。
昆虫的行为是昆虫为了更好地适应环境而调节自己生活方式的活动,对昆虫行为活动规律的准确掌握,是人工饲养、生物防治、生理生态等相关内容研究的重要基础[11]。目前,有关光肩星天牛成虫行为学的研究,有配偶定位及交尾行为[12-15]、产卵选择行为[16-17]、主动扩散行为[18-19]、对不同寄主植物及其挥发物的趋向行为[20-22]等。而未见对该虫日活动行为节律的详细报道,仅在生物学特征[23]、行为学观察[24]研究中有部分零星描述。研究光肩星天牛成虫日活动节律,对深入了解其生物学特性,阐明其生活习性,研究该虫的行为学、田间抽样调查、预测预报及其综合治理等均有重要的意义。鉴于此,本试验在养虫室内对光肩星天牛成虫的日活动节律进行了详细观察。
1 材料与方法
1.1 供试虫源和饲养条件
2014年6月30日—7月10日,在河北农业大学西校区馒头柳(Salixmatsudana‘Umbraculifera’)和垂柳(Salixbabylonica)树上采集光肩星天牛成虫,约150头。随机选择性成熟、可正常交尾、健康活泼的雌雄成虫(10~15日龄)[25-27]接入自制养虫笼(直径为24 cm,高22 cm)中;为了保证养虫笼内部和养虫室的温湿度一致,选用带网眼的垃圾篓(材料为:聚乙烯)改装制成养虫笼;上方用纱布覆盖以防止试虫飞出,纱布上放置湿棉球供试虫补充水源,每日08:00和20:00进行补水,棉球的水分含量以达到潮湿但不滴水为宜;给予垂柳枝条进行饲喂,在养虫笼底部放置大约2 cm厚的湿度较大的细沙,以尽量保证柳条在48 h内含水量一致。每笼接入雌雄成虫1对,分别设置20个重复。饲养条件:温度(27±1)℃,相对湿度(65±10)%,光周期(日出时间,05:03—05:07;日落时间,19:43—19:44)。为了避免影响试虫活动规律,在试验开始第1天和第3天给试虫补充食物。
1.2 行为观察
试虫在上述条件下适应5日后,从7月10日06:30开始至7月15日06:00结束,每隔30 min记录一次,连续观察5 d;晚上,在红光灯下观察。期间,试虫死亡及时补接。共观察了240次,获得9 600个行为数据。
1.3 行为特征划分
根据预试验中光肩星天牛成虫在养虫笼中所表现的行为活动,将其划分为4类。①取食:观察时成虫头部口器接触柳条并且有取食动作。②交尾:观察时雌雄成虫处于配对状态。③移动:观察时成虫正在养虫笼中爬行。④静息:观察时成虫处于静止不动状态。
1.4 行为时间划分
日时间划分:白天06:30—19:30;黑夜20:00—06:00。根据所表现的日活动规律又进一步划分为:凌晨(04:00—06:00),上午(06:30—10:30),中午(11:00—13:30),下午(14:00—16:00),黄昏(16:30—19:30),晚上(20:00—03:30)。
1.5 数据统计与分析
采用Kruskal-WallisH检验分析光肩星天牛成虫24 h及6个时间段内的活动行为差异,采用Mann-WhitneyU检验各行为在白天和黑夜及雌雄虫间的差异[11]。数据在IBM SPSS 21.00和Micro office 2003进行分析,作图在Origin 9.0上完成。
2 结果与分析
2.1 光肩星天牛成虫日活动行为规律
光肩星天牛成虫4种活动行为日节律如图1所示。取食行为高峰期出现在21:30(49%),12:00最低(15%)。在一天中均有交尾行为发生,高峰期出现在12:00(34%),22:00、03:00、04:30交尾行为发生较少(12%~13%)。移动行为高峰期出现在10:30(36.5%),00:30—01:30发生该行为最少(6.5%~7.0%)。静息行为高峰期出现在08:00—09:00(36%~41%),13:30和15:30发生该行为最少(15.5%)。在一天中各时刻之间,取食行为(χ2=138.744,df=47,p<0.001)、交尾行为(χ2=159.616,df=47,p<0.001)、移动行为(χ2=180.665,df=47,p<0.001)和静息行为(χ2=81.346,df=47,p<0.001)均存在极显著性差异。
2.2 不同时间段内光肩星天牛成虫活动行为的规律
2.2.1 昼夜活动行为的节律
光肩星天牛成虫活动行为在白天和黑夜存在差异(表1)。白天,4种活动行为中以静息行为所占比例最多,其次为移动行为、交尾行为,取食行为最少;4种活动行为之间无显著性差异。黑夜,4种活动行为以取食行为所占比例最多,其次为静息、交尾行为,移动行为最少;各活动行为之间存在极显著性差异。
白天和黑夜,取食、静息行为黑夜高于白天,交尾和移动行为白天高于黑夜。交尾(U=0,p<0.001)和移动(U=7.000,p<0.001)行为具有极显著性差异,取食(U=17.000,p=0.013)行为存在显著性差异,静息行为(U=37.000,p=0.326)无显著性差异。
2.2.2 6个时间段内活动行为的节律
一天,取食行为从凌晨到上午呈下降趋势、之后逐渐上升;交尾行为从凌晨到中午呈上升趋势,直到中午升为最高,之后逐渐下降;移动行为从凌晨到上午呈上升趋势,上午、中午、下午变化不大,之后逐渐下降;静息行为从凌晨到下午呈下降趋势,之后呈上升趋势(表1)。交尾(χ2=43.046,df=5,p<0.001)、移动(χ2=26.591,df=5,p<0.001)和静息行为(χ2=17.116,df=5,p=0.004)具有极显著性差异,取食(χ2=14.059,df=5,p=0.015)行为存在显著性差异。在各个时间段,4种活动行为之间中午存在显著性差异;凌晨、上午和晚上存在极显著性差异;下午和黄昏无显著性差异。
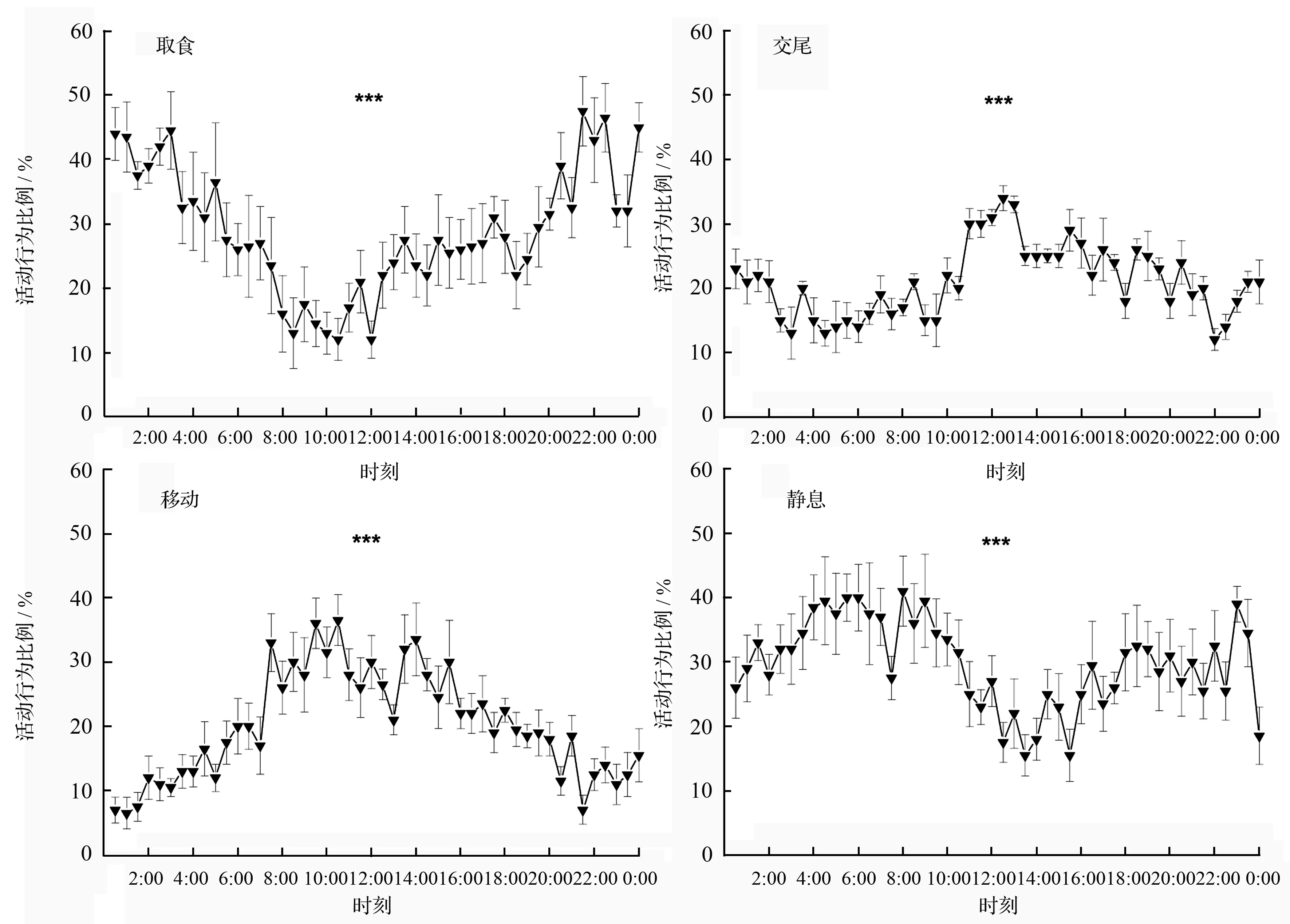
图中数值为平均值±标准误(n=5)。*** 表示经Kruskal-Wallis H测验4种活动行为之间在0.001水平上呈显著性差异。

表1 光肩星天牛成虫不同时间段内的活动行为
2.3 性别对光肩星天牛成虫活动行为的影响
2.3.1 对活动行为日节律的影响
性别对光肩星天牛成虫活动行为日节律有一定影响。在一天内,雌雄虫取食、移动、静息3种行为活动的变化情况较相似,仅仅是在数值和时间上具有一定差异(图2)。一天各个时刻,雌虫发生取食行为所占的比例低于雄虫;取食高峰期,雄虫出现在21:30(49%)、雌虫在21:30和22:30(46%);低谷,雄虫在12:00(15%)、雌虫在10:00—10:30(7%)。移动行为,雌虫所占的比例高于雄虫;高峰期雄虫出现在9:30(31%)、雌虫在10:30(43%);低谷,雄虫在01:00(3%)、雌虫00:30—01:30(7%)。静息行为,雌雄虫发生该行为所占的比例相似;高峰期雄虫在08:00(47%)、雌虫在09:00(44%);低谷,雄虫在00:00(10%)、雌虫在14:00和15:30(12%)。在一天各时刻之间,雄虫取食和静息行为均无显著性差异,移动行为存在极显著性差异;雌虫除静息行为无显著性差异外,取食和移动行为均存在极显著性差异。

图中数值为平均值±标准误(n=5)。ns表示p>0.05,*** 表示p<0.001。
2.3.2 对昼夜活动行为的影响
光肩星天牛雌雄成虫在白天和黑夜发生各行为情况如表2所示。白天,雄虫各行为发生比例由高到低依次为静息、取食、交尾、移动;雌虫为移动、静息、交尾、取食。黑夜,雌雄虫均为取食、静息、交尾、移动。各行为之间白天无显著性差异,黑夜存在极显著性差异。
雌雄成虫取食、静息行为表现为黑夜高于白天,交尾、移动行为白天高于黑夜。雄虫,除了交尾行为具有极显著性差异外(U=0,p=0.008);其他行为均无显著性差异:取食(U=5.000,p=0.151)、移动(U=4.000,p=0.095)、静息行为(U=12.000,p=1.000)。雌虫,交尾(U=0,p=0.008)和移动(U=0,p=0.008)行为均存在极显著性差异,取食行为(U=2.000,p=0.032)呈显著性差异,静息行为(U=8.000,p=0.421)无显著性差异。
在白天和黑夜,雄虫取食行为均高于雌虫、移动行为雌虫高于雄虫;静息行为,白天雄虫略高于雌虫,黑夜雌虫略高于雄虫。4种活动行为雌雄虫之间均无显著性差异。
2.3.3 对6个时间段内活动行为的影响
在凌晨、上午等6个时间段,雌雄虫取食和移动行为发生比例有一定差异,静息行为发生比例无显著性差异(表2)。雄虫上午发生移动行为最多、雌虫以中午最多;取食行为均是上午最低,晚上最多;交尾行为以凌晨最少,中午最多;静息行为中午最少,凌晨最多。一天各个时间段4种活动行为之间,雄虫除晚上存在极显著性差异外,其它时间段均无显著性差异;雌虫上午存在显著性差异,中午和晚上存在极显著性差异,其它时间段均无显著性差异(表2)。
一天6个时间段之间,雄虫,除交尾行为(χ2=21.158,df=5,p<0.001)存在极显著性差异外,其它行为均无显著性差异:取食(χ2=5.288,df=5,p=0.382)、移动(χ2=9.158,df=5,p=0.103)、静息(χ2=7.844,df=5,p=0.165);雌虫,静息行为呈显著性差异(χ2=11.751,df=5,p=0.038),交尾(χ2=21.158,df=5,p<0.001)和移动(χ2=21.112,df=5,p<0.001)行为呈极显著性差异,取食行为(χ2=9.914,df=5,p=0.078)无显著性差异。
光肩星天牛雌雄虫在凌晨、上午等6个时间段的活动行为存在一定差异。各个时间点,发生取食行为的比例雄虫均高于雌虫,移动行为雌虫高于雄虫;静息行为凌晨到下午雄虫高于雌虫、黄昏和晚上雌虫高于雄虫。一天内,雌雄虫中午发生取食行为的比例呈显著性差异(U=3.000,p=0.047);上午(U=3.000,p=0.047)和中午(U=1.000,p=0.016)发生移动行为的比例呈显著性差异、凌晨(U=0,p=0.009)呈极显著性差异;其它各个时间段各行为雌雄虫之间均无显著性差异。

表2 不同时间段内光肩星天牛雌雄成虫行为活动的差异
3 结论与讨论
本试验观察发现该成虫一天中,静息时间约占总时间的1/3,其余时间均进行取食、移动、交尾等行为活动;这与张波等[24]在3 m×3 m×3 m自制养虫笼中观察发现:该成虫白天活动、晚上静息的试验结果有一定出入。另外,本试验发现该虫在24 h内均有交尾现象发生,这与张玉凤等[28]研究认为成虫的交尾行为不受温度限制,在24 h内均有发生的结果相符;但是,由于试验所设置的温度为恒温(27±1)℃,因此,温度是否影响交尾行为的发生还需要进一步去论证。另外,本试验还观察到:雌、雄虫取食行为的高峰期发生在21:30—22:30(46%~49%),取食低值却出现在10:00—12:00(7%~15%);而且晚上发生取食行为的比例显著高于其它行为。以上通过取食和交尾行为的观察均可以说明该虫在晚上也处于活动状态。
本试验观察到光肩星天牛的交尾行为存在明显的日节律(p=0.002),高峰时段出现在11:30—13:00(30%~34%)、15:30(29%)、23:00—01:30(21%~23%);该结果和周嘉熹等[29]观察黄斑星天牛(Anoplophoranobilis)交尾行为节律10点左右和18点左右存在2次高峰期的结果有一定的差异;与张波等[24]观察到光肩星天牛在11:00、18:00、21:00左右为交尾高峰期的结果较相似。尚玉昌[30]认为自然条件下,生物节律由于受到光暗周期的影响,呈现严格的24 h节律;因此,这种节律会随着光暗周期的改变而进行调节。本试验结果与张波等[24]观察结果的差异,也许是由于地理位置的不同造成光暗周期的差异,从而影响了该虫的节律行为。根据光肩星天牛所表现出的日活动规律,将一天划分为凌晨、上午、中午、下午、黄昏和晚上6个时间段,发现交尾行为在6个时间段之间存在极显著性差异(p<0.001);交尾行为发生高峰期出现在中午,随后开始下降。这与田间采虫时,发现上午采集到的试虫以未交尾单头成虫为主,而在中午和下午以交尾成对的成虫为主相一致。在田间,也许是由于中午温度升高,导致该虫向树体下方(温度较低区域)聚集,从而该时间段的交尾行为发生比例增加;但是,本试验是在室内恒温条件下进行,这说明交尾行为是受日节律的影响,而非温湿度等条件的影响。在对果蝇及仓鼠等哺乳动物的研究中已发现存在一个调节正常昼夜行为节律的基因(clock基因),并且证明clock基因明显受到光周期的影响[31]。也许光肩星天牛成虫也存在一个类似的clock基因,来感受光周期的变化从而调节各种活动行为的日节律。
一天各个时间段,光肩星天牛4种行为中取食行为雄虫均略高于雌虫,也许是由于雄虫个体小于雌虫导致耐饥饿能力较差;在做行为学试验中进行饥饿处理时,同样发现雌虫的抗饥饿能力明显高于雄虫。移动行为雌虫高于雄虫,这也许是由于雌虫不仅需要补充营养进行取食,而且还需要寻找适合的产卵场所导致的。对4种活动行为进行显著性差异分析,发现移动行为在凌晨到中午时间段雌虫显著高于雄虫(p凌晨=0.009,p上午=0.047,p中午=0.016),也许是雌虫在寻找合适的产卵场所导致移动行为在凌晨、上午和中午时间段与雄虫表现出差异,因此,推断产卵行为的高峰期应位于上午或者中午时间段,并且存在明显的日节律。由于试虫的食物为直径0.5~2.0 cm左右的柳树枝条[32]、产卵行为无法分辨等条件所限导致产卵行为的日节律数据没有获得。如何确定该虫产卵行为、产卵行为是否存在日节律以及该节律又是如何变化需要进一步深入研究。
本研究表明,光肩星天牛移动高峰期雄虫在09:30、雌虫在10:30。因此,建议在该时间段采集虫源,以保证采集到的虫源具有较高的活性;同时建议,在对该虫种群结构调查时,在移动高峰期进行,即09:30—10:30;另外,在进行不同环境立地条件中该虫的种群结构比较研究时,应尽量在相同的时间段内进行以减少因成虫活动行为的节律变化导致试验误差。另外,本研究仅在试验室内(27±1)℃、相对湿度(65±10)%条件下,观察了该虫的取食、交尾、移动、静息4种行为日活动节律,对于自然条件下,该虫的活动、交尾、产卵等行为的日活动节律还不清楚,这些均有待于进一步研究。
[1] Haack R A, Hérard F, Sun J, et al. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: a worldwide perspective[J]. Annual Review of Entomology,2010,55:521-546.
[2] 王志刚.中国光肩星天牛发生动态及治理对策研究[D].哈尔滨:东北林业大学林学院,2004.
[3] Williams D W, Lee H P, Kim I K. Distribution and abundance ofAnoplophoraglabripennis(Coleoptera: Cerambycidae) in natural Acer stands in South Korea[J]. Environmental Entomology,2004,33(3):540-545.
[4] Smith M T, Bancroft J, Li G, et al. Dispersal ofAnoplophoraglabripennis(Cerambycidae)[J]. Environmental Entomology,2001,30(6):1036-1040.
[5] Gaag D J, Loomans A. Host plants ofAnoplophoraglabripennis, a review[J]. EPPO Bulletin,2014,44(3):518-528.
[6] 阎俊杰,阎晔辉.光肩星天牛生态控制模式的研究[J].河北农业大学学报,1999,22(4):83-87.
[7] 潘宏阳,郑华,方国飞,等.我国杨树天牛综合治理回顾与展望[J].南京林业大学学报:自然科学版,2005,29(4):1-5.
[8] Hérard F, Ciampitti M, Maspero M, et al.Anoplophoraspecies in Europe: infestations and management processes[J]. EPPO Bulletin,2006,36(3):470-474.
[9] Herard F, Maspero M, Ramualde N, et al.Anoplophoraglabripennisinfestation (col.: cerambycidae) in Italy[J]. EPPO Bulletin,2009,39(2):146-152.
[10] Hu J, Angeli S, Schuetz S, et al. Ecology and management of exotic and endemic Asian longhorned beetleAnoplophoraglabripennis[J]. Agricultural and Forest Entomology,2009,11(4):359-375.
[11] 杨茂发,杨大星,徐进,等.稻水象甲成虫活动行为的日节律[J].昆虫学报,2013,56(8):952-959.
[12] 贺萍,黄竞芳.光肩星天牛成虫的行为[J].昆虫学报,1993,36(1):51-55.
[13] 李德家,所雅彦,中岛忠一.光肩星天牛成虫交配行为机制研究[J].北京林业大学学报,1999,21(4):33-36.
[14] 陆群,张玉凤,张宏世.光肩星天牛求偶、交尾及产卵行为的研究[J].内蒙古林业科技,1998(3):8-9.
[15] Wickham J D, Xu Z, Teale S A. Evidence for a female-produced, long range pheromone ofAnoplophoraglabripennis(Coleoptera: Cerambycidae)[J]. Insect Science,2012,19(3):355-371.
[16] Morewood W D, Neiner P R, Mcneil J R, et al. Oviposition preference and larval performance ofAnoplophoraglabripennis(Coleoptera: Cerambycidae) in four eastern North American hardwood tree species[J]. Environmental Entomology,2003,32(5):1028-1034.
[17] 赵博光,李周直,葛庆杰.光肩星天牛在杨树上产卵部位的选择[J].北京林业大学学报,1997,(3):29-33.
[18] 黄竞芳,骆有庆,周章义.中国光肩星天牛研究的新进展[J].陕西林业科技,1992,(2):57-62.
[19] 温俊宝,李友常,夏乃斌,等.杨树光肩星天牛成虫扩散格局的研究[J].生态学报,1998,18(3):47-55.
[20] 李建光,金幼菊,骆有庆,等.光肩星天牛不同寄主树种挥发性物质的比较分析[J].北京林业大学学报,2002,24(Z1):169-173.
[21] 阎雄飞,李晓娟,骆有庆,等.光肩星天牛成虫对原寄主枝条挥发物趋向的测定[J].北京林业大学学报,2008,30(3):80-84.
[22] 张风娟,金幼菊,陈华君,等.光肩星天牛对4种不同槭树科寄主植物的选择机制[J].生态学报,2006,26(3):870-877.
[23] 赵瑞良,吕晓宏,武祥云.光肩星天牛生物学特性研究[J].山西林业科技,1991(4):1-6.
[24] 张波,徐庆林.光肩星天牛成虫行为的研究[J].森林病虫通讯,1991(4):14-17.
[25] 王涛,温俊宝,许志春.鉴别光肩星天牛雌雄成虫的一种简便方法[J].中国森林病虫,2004,23(5):42-44.
[26] Keena M A.Anoplophoraglabripennis(Coleoptera: Cerambycidae) fecundity and longevity under laboratory condition: comparison of populations from New York and Illinois onAcersaccharum[J]. Environmental Entomology,2002,31(3):490-498.
[27] Smith M T, Bancroft J, Tropp J M. Age-specific fecundity ofAnoplophoraglabripennis(Coleptera: Cerambycidae) on three tree species infested in the United States[J]. Environmental Entomology,2002,31(1):76-83.
[28] 张玉凤,陆群,张宏世,等.光肩星天牛成虫特性及其行为的研究[J].内蒙古林业科技,1997(4):7-9.
[29] 周嘉熹,张克斌,逯玉中.黄斑星天牛成虫行为及其对树种的选择性[J].西北林学院学报,1984(1):119-129.
[30] 尚玉昌.动物行为学[M].北京:北京大学出版社,2005:237-239.
[31] 蒋志刚.动物行为原理与物种保护方法[M].北京:科学出版社,2004:57-59.
[32] 沈荣武,章士美.光肩星天牛的研究[J].昆虫知识,1966(3):153-155.
Diurnal Rhythm of Four Types of Adult Behaviors ofAnoplophoraglabripennis//
Lü Fei, Hai Xiaoxia, Wang Zhigang, Yan Aihua, Bi Yongguo, Liu Bingxiang
(Key Laboratories for Germplasm Resources of Forest Trees and Forest Protection of Hebei Province, Agricultural University of Hebei, Baoding 071000, P. R. China)//Journal of Northeast Forestry University,2015,43(9):90-95.
AllAnoplophoraglabripennisbehaviors of foraging (FG), mating (MT), moving (MG), and resting (RG) was observed by pair (male and female) rearing individually in the laboratory. The happen percentages of various behaviors were significant difference in all day. The peak of FG was at 21:30 (49%), MT was at 12:00 (34%), MG was 10:30 (36.5%), and RG was 8:00-9:00 (36%-41%). The happen percentages of various behaviors had significant difference at different time period of all day. The happen percentages of FG and RG at night were higher than those at day, but the MT and MG at day were higher than those at night. The MT, MG and RG had extremely significant difference, and the FG had significant difference at six periods including early morning, morning, noon, afternoon, dusk, and night. The diurnal rhythm of various behaviors was affected by sexual difference to a certain degree, and the peak of various behaviors had difference between females and males. The happen percentages of various behaviors had no significance different between females and males at the day and night, the happen percentage of FG at noon, the happen percentage of MG at morning and noon had significant differences, and the MG of early morning had extremely significant difference at the different times period of all day by the Mann-WhitneyUtest. Therefore, the temporal variation and sexual difference could significantly influence the activity behaviors ofA.glabripennis.
Anoplophoraglabripennis; Adult; Sex difference; Insect behavior; Diurnal rhythm
1)国家自然科学基金项目(C30571501);河北省自然科学基金项目(C2012204098);河北省科技支撑计划项目(12222905)。
吕飞,男,1982年3月生,河北省林木种质资源与森林保护重点实验室(河北农业大学),博士研究生。E-mail:haimolv@foxmail.com。
王志刚,河北省林木种质资源与森林保护重点实验室(河北农业大学),教授。E-mail:wzhg@hebau.edu.cn。
2015年4月10日。
S763.38
责任编辑:程 红。
