大草蛉Chrysopa pallens(Rambur)触角cDNA 文库的构建
2014-12-16张礼生张海平刘晨曦王孟卿陈红印
王 娟,张礼生,张海平,刘晨曦,王孟卿,陈红印
(中国农业科学院植物保护研究所农业部作物有害生物综合治理重点实验室,中国-美国生物防治实验室,北京 100081)
大草蛉Chrysopa pallens(Rambur)属于脉翅目Neuroptera 草蛉科Chrysopidae,是叶螨、蚜虫、鳞翅目卵及低龄幼虫等多种农林害虫的重要天敌,在害虫生物防治中极具应用价值,广泛分布在我国大部分省市(林美珍等,2007)。天敌昆虫的行为受多种因素的影响,比如环境信息化合物、温度、湿度等。大草蛉成虫释放于田间或温室大棚后由于诸多因素的影响,通常会发生逃逸或者死亡,定殖性较差,严重降低了其利用价值(武鸿鹄,2014)。触角是昆虫感觉系统的重要组成部分,行使嗅觉和味觉功能,昆虫感受外界气味物质主要是通过触角上的感受器来完成的(Steinbrecht,1997;Steinhbrecht & Stankiewicz,1999),在长期的进化过程中,昆虫嗅觉系统已经发展成为一个高度专一、极其灵敏的化学监测系统。昆虫通过这种复杂的嗅觉系统来鉴定和识别环境中的特异性化学气味分子,并将这些化学信号转化为电信号,从而启动了昆虫的一系列专一行为反应,如寻找食物和配偶、搜寻产卵及生殖场所、避免不利环境的伤害等(Kaissling et al.,1987;Breer et al.,1994;Field et al.,2000),而这些功能主要是通过昆虫触角中几种嗅觉相关蛋白与外界气味分子的结合来实现的。对昆虫嗅觉相关基因深入研究,有利于从分子水平明确昆虫的嗅觉识别机制,并从生理生化水平了解昆虫触角与昆虫觅食、产卵、交配等昆虫行为的关系(Benton et al.,2009;Zhou et al.,2010)。从利用捕食性天敌昆虫进行害虫生物防治角度,掌握了天敌昆虫嗅觉相关蛋白与外界气味化合物之间的作用机理,有利于人们对天敌昆虫行为的控制,通过释放对天敌昆虫有较强吸引作用的气味化合物于田间或温室大棚,从而提高其在某一特定空间的定殖成功率。随着分子生物学的发展,全长cDNA 文库的构建、测序和功能注释,为深入开展昆虫触角嗅觉相关基因的克隆及信息识别机制的研究,提供了成本低廉且信息量丰富的技术平台。Li 等(2014)运用SMART 技术构建了云斑天牛Batocera horsfieldi(Hope)(Coleoptera:Cerambycidae)触角全长cDNA 文库,并成功筛选出了4个气味结合蛋白基因,2个性信息素结合蛋白基因及3个化学感受蛋白基因,为云斑天牛嗅觉相关基因的功能研究奠定了基础。张帅等(2009a,2009b)通过对棉铃虫Helicoverpa armigera(Hübner)cDNA文库基因筛选和生物信息学分析,鉴定获得11个气味受体基因,并克隆表达了信息素结合蛋白HarmPBP2。Gao & Wang(2014)从松褐天牛Monochamus alternatus Hope 触角cDNA 文库中克隆并表达了4种气味结合蛋白,并研究了其与17种植物挥发性化合物的荧光竞争结合特性,结果表明MaltOBP3 和MaltOBP5 与某几种特定植物挥发物有较强的结合作用。本研究应用Infusion SMART技术构建了大草蛉雌雄触角cDNA 文库,填补了脉翅目昆虫cDNA 文库数据信息的稀缺,丰富了大草蛉基因序列信息,为克隆发掘大草蛉嗅觉相关基因及探索其嗅觉识别机制奠定基础,为合理利用大草蛉进行生物防治提供理论依据。
1 材料与方法
1.1 材料
1.1.1 供试虫源
大草蛉采自中国农业科学院廊坊科研中试基地棉田和玉米田,成虫于室内人工气候箱中饲养,寄主为大豆蚜Aphis glycines,建立稳定种群,饲养条件:温度(26±2)℃,相对湿度(50±10)%,光周期L∶D=14 h∶10 h。羽化三天之内剪下触角,用离心管放液氮中收集50 对以后放-70℃冰箱备用。
大豆蚜采自中国农业科学院廊坊科研中试基地大豆田,室内以大豆Glycine max L.,(品种为化诱5 号)为寄主饲养多代,建立稳定种群。饲养条件:温度(25±2)℃,相对湿度(50±10)%,光周期L∶D=14 h∶10 h。
1.1.2 主要试剂
Trizol 提取试剂购自Invitrogen 公司;氯仿、异丙醇、乙醇、琼脂糖、LB 培养基制备所用的胰蛋白胨、酵母提取液、AMP、X-gal、IPTG 琼脂粉、二甲苯蓝等试剂购自中保科农生物科技有限公司;In-Fusion SMART cDNA 文库构建试剂盒购自Clontech 公司(包括In-Fusion SMARTer PCR cDNA Synthesis Kit;In-Fusion HD Cloning Kit;In-Fusion SMARTer Directional Cloning & Screening Component;Advantage?倕2 PCR Kit);大肠杆菌Escherichia coli 感受态细胞DH5α 购自北京全式金生物技术有限公司。其他均为进口或国产分析纯试剂。
1.2 引物序列
第一链cDNA 合成引物In-Fusion SMART CDs Primer(12μM):
5'-CGGGGTACGATGAGACACCATTTTTTTTT TTTTTTTTTTTVN-3'
(N=A,C,G,or T;V=A,G,or C)
双链cDNA 合成引物:
3'In-Fusion SMARTer PCR Primer(12μM):5'-CGGGGTACGATGAGACACCA-3'
菌液PCR 检测采用引物:
F:5'-TCACACAGGAAACAGCTATGA-3',R:5'-CCTCTTCGCTATTACGCCAGC-3'
1.3 大草蛉触角总RNA 提取及cDNA 合成
采用Trizol 法提取大草蛉成虫触角总RNA。用准备好的剪刀剪下已被乙醚处理的大草蛉触角约50-100 mg 放入预冷的干净研钵中,加入液氮后,迅速研磨成粉末状,加入1 mL Trizol,充分混匀后,移入1.5 mL 离心管中,氯仿抽提,加入异丙醇沉淀RNA,75% 乙醇洗涤2 次,溶于30μL Rnase-free water。用琼脂糖凝胶电泳和Nanodrop ND-2000 定量总RNA 的纯度和浓度。
cDNA 第一链合成按照In-Fusion ® SMARTeTMDirectional cDNA Library Construction Kit 说明书进行。在无菌1.5 mL 离心管中依次加入:提取总RNA 1μg(约2μL),In-Fusion SMART CDs Primer(12μM)1μL,去离子水加至4.5μL,混匀,72℃孵育3 min 后,42℃放置2 min。再向反应管中依次加入以下试剂:5×First-Strand Buffer 2μL,DTT(100 mM)0.25μL,dNTP Mix(10 mM)1μL,SMART V oligonucleotide(12μM)1μL,Rnase Inhibitor 0.25μL,SMARTscribeTMReverse Transcriptase(100 U)1μL。简单离心后,42℃温浴90 min,68℃孵育10 min 终止第一链cDNA 的合成。将cDNA 放于-20℃保存备用。
1.4 LD-PCR 合成双链cDNA
于100μL PCR 反应体系中加入第一链cDNA 2μL,去离子水80μL,10×Advantage 2 PCR Buffer 10μL,50×dNTP Mix(10μM)2μL,5'PCR Primer ⅡA(12μM)2μL,3'In-Fusion SMARTer PCR Primer(12μM)2μL,50×Advantage 2 Polymerase Mix 2μL。简单离心混匀。反应条件为条件体系:oliacoli 95 ℃预变性1 min;95℃变性15 s,65℃退火30 s,68℃延伸6 min,先设置15个循环,然后暂停程序,转移70μL 至第二个反应管中,于4℃保存。余下的30μL 继续扩增,每加3个循环,取出5μL 于4 ℃保存,直至达到27个循环数。将不同循环数的产物进行电泳检测,确定最适循环数。复苏保存的产物,加至最适循环数后,加入10μL 1%的二甲苯蓝染料,混匀后,于-20℃保存备用。将循环合适的PCR 产物进行CHROMA SPIN+TE-1000 过柱后,选取0.4-2.0 kb 之间的cDNA 片段与载体连接。
1.5 ds cDNA 的纯化与分离
按照试剂盒说明书,将CHROMA SPIN+TE-1000 柱子取出,颠倒数次,以便混匀悬浮胶体。将分离柱固定好,移去底盖,使贮存Buffer 通过重力作用留下,调整流速为1 d/40-60 s,待贮存Buffer 流尽后,沿着柱管壁缓慢加入700μL 柱Buffer,直到流尽,(约15~20 min),缓慢的加入含二甲苯蓝的ds cDNA 于分离柱内,静置2 min,使样品完全吸附于基质中,加入100μL 的柱Buffer 冲洗含有ds cDNA 的离心管,并缓慢加入到分离柱内。准备16个收集管至于柱子下面,第一个离心管对着柱子下面的出口。加入600μL 柱Buffer 后,立即收集,每管一滴(约35μL),直到收集完全部的离心管。每管取3μL 于1.1%的琼脂糖凝胶电泳上检测,染料迁移1-2 cm 时停止电泳(若时间太长,很难看到条带)。根据观察结果,收集片段最长的3-4 管ds cDNA 重新混合到一个新的离心管中。向离心管中依次加入1/10 体积的 Sodium Acetate(3 M;pH4.8),Glycogen(20 mg/mL)1.3μL,2.5 倍体积的95% 乙醇。前后晃动,充分混匀。于-20 ℃过夜保存,以便沉淀ds cDNA。次日,室温,14 000 rpm,离心20 min。用枪小心的移去上清,不要触及底部沉淀,充分干燥。向试管中加入600μL 80%的乙醇,室温下14 000 rpm 离心5 min。小心移去上清,不要碰及沉淀,干燥10 min。用10μL 去离子水重新悬浮沉淀,轻微混匀,于-20 ℃保存备用。
1.6 ds cDNA 与pSMART2IFD 载体连接
cDNA 片段和载体的连接比率是决定文库转化效率和文库库容的关键因素。因此,为了得到最佳的连接效率,在此建立了3个连接反应体系(表1)。

表1 In-fusion 克隆连接体系Table 1 In-fusion cloning reactions
加完上述试剂后轻轻混匀,短暂离心,50℃孵育15 min,然后转移至冰上。向连接体系A,B,C 中分别加入TE 90μL,Quick Clean Resin 10μL,涡旋1 min,将上清液转移至新的离心管中,向上清液中加入1.2μL glycogen,混匀后,加入280μL 无水乙醇,前后震荡混匀。将离心管与-70℃放置过夜。次日,15 000 rpm 离心20 min,小心吸走乙醇,不要触及底部沉淀。空气中干燥后,用10μL 无菌水重新悬浮。至此得到3种不同连接反应的重组质粒。
1.7 重组质粒电转入E.coli 感受态细胞
将上述含A,B,C 三管中的重组质粒分别加入解冻后的DH5α 电转感受态细胞,每管25μL,轻轻混匀。电转条件:电击杯内径0.1 cm,电转参数为2 000 V,200 Ω。电转完成后转移到加有965μL 的LB 培养基上,于37℃225 rpm 条件下孵育1 h。每管分别取出2μL 电转物涂布在含有氨苄霉素的培养基上,37℃倒置培养过夜。次日,检查菌落生长状况,从而确定最佳的连接比例。
1.8 文库质量鉴定
挑取文库中单克隆若干个,在液体LB 培养基中培养过夜,次日取1μL 作为菌液PCR 的模板,加入PCR Mix(2×)10μL,正向和反向引物各1μL(10μM),加去离子水至20μL,混匀,离心。PCR 反应程序如下:95℃变性30 s,65℃退火30 s,68℃延伸3 min,3个循环;95℃变性30 s,68℃延伸3 min,25个循环。PCR 完成后在1.5%的琼脂糖凝胶电泳上检测插入片段的大小。
1.9 文库滴度的测定
在将原始cDNA 文库冷冻保存之前,应当确定检测其滴度(以1μL 文库菌液涂板产生的单克隆数表示)。美国Clontech 公司一个良好的基因文库标准为:原始文库的重组子数据应达到5×105-5×108。插入的cDNA 片段不小于300 bp。
文库滴度测定方法如下:
分别取0.5μL,0.25μL,0.05μL,0.025μL,0.005μL 涂布LB 平板,37℃培养12 h 左右,计算长出的单克隆数目。
文库滴度(pfu/mL)=(单克隆数×稀释倍数)/受体菌体积
2 结果与分析
2.1 大草蛉触角RNA 的提取
琼脂糖甲醛变性凝胶电泳检测总RNA,结果如图1(示雄性触角总RNA,雌同)。由图可见28s rRNA,18s rRNA 条带清晰可见,5s rRNA 及其他小核糖体RNA 条带微弱可见,表明提取的RNA完整性较好,也没有明显的DNA 污染。经Nanodrop ND-2000 测得总RNA 浓度为408 ng/μL,A260/A280值为1.91,表明提取RNA 纯度较高,完全符合构建文库的质量要求。

图1 大草蛉雄虫触角总RNA 电泳Fig.1 Total RNA of Chrysopa pallens male antennae
2.2 最佳循环数的确定
PCR 产物经过1.1%的琼脂糖凝胶电泳检测(图2),结果显示,设置18个循环数条带较亮,表明在LD-PCR 合成体系中循环数为18 较为合适。
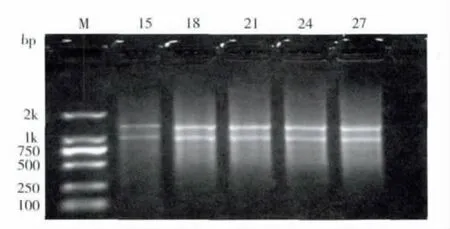
图2 不同循环数下的LD-PCR 扩增产物琼脂糖凝胶电泳Fig.2 Agarose gel electrophorests of ds cDNA based on different cycles
2.3 ds cDNA 分析
第一链cDNA 经LD-PCR 扩增后,其双链片段成弥散带,大小主要集中于0.4-2.0 kb 之间(图2),且均一性良好,并且可见几条明显的亮带,代表触角中含量比较高的mRNA。
2.4 ds cDNA 的纯化与分离
由于小片段比较容易连接到载体上,因此需要经过过柱分级分离,除去小片段的cDNA 片段和引物片段。利用CHROMA SPIN+TE-1000 柱除去小片段的cDNA 片段,分离ds cDNA 片段,共收集16 管洗脱液,电泳结果如图3,泳道1-16 代表按不同长度分级分离得到的ds cDNA,由图可以看出,第8-11 管ds cDNA 电泳产物清晰可见,并且长度适中,故收集此4 管的ds cDNA 合并成一管,和pSMART2IFD 载体连接并且电转化DH5α感受态细胞。

图3 按片段大小分离ds cDNAFig.3 Separation of ds cDNA by different length fraction
2.5 菌液PCR 鉴定文库插入片段的大小
随机挑取单克隆(>1000个)进行菌液PCR和酶切鉴定插入cDNA 片段的大小和文库重组率,部分结果如图4 示,由图可知,插入片段长度基本都在500 bp 以上,重组率在93%以上,说明构建的文库质量较高。
2.6 cDNA 文库滴度的测定
参照In-Fusion®SMARTer®Directional cDNA Library Construction Kit 说明书计算构建的大草蛉触角cDNA 文库的滴度,最后测得雄性触角cDNA 文库滴度为1.8×106pfu/mL,雌性触角cDNA 文库的滴度为2.1×106pfu/mL。
3 结论与讨论
高质量的RNA 是成功构建全长cDNA 文库的关键,28S 和18S 是真核生物RNA 的两个主要亚基,电泳时一般28S 的亮度约为18S 的两倍。本试验在反复提取总RNA 后,几次电泳得到的结果均显示18S 条带远比28S 亮,这与Gambacurta 等(1993)在提取昆虫触角RNA 的试验结果吻合。Fujiwara & Ishikawa(1986)认为这种现象的产生是由于28S RNA 较18S RNA 容易断裂,且断裂产物大小与18S 相当,导致28S 含量的减少或缺失而18S 相对偏多的情况。本试验得到的总RNA 具有较好的完整性(A260/A280=1.91),纯度及完整性均达到文库构建要求,完全可以用来合成双链cDNA。

图4 菌液PCR(A)和酶切鉴定(B)cDNA 文库插入片段大小和重组率Fig.4 The inserts length and recombinant efficiency of the antennal cDNA library evaluated by PCR or enzyme digestion
过柱分离过程中,分离片段的大小会影响到文库的质量,是决定cDNA 文库插入片段平均长度和初始文库滴度的关键因素。如果为保证文库的插入片段平均长度而筛除过多的cDNA 片段,在构建文库过程中,势必会降低文库的初始滴度,因此可能丢掉一些基因,降低文库的质量。若不能有效筛除短片段,则其将与载体优先连接,这样就会使文库平均插入片段长度缩短,从而大大降低文库的应用价值(张丽芳等,2012)。本试验在cDNA 片段与载体连接之前,将合成的cDNA 通过CHROMA SPIN+TE-1000 分离柱进行分级分离,可以有效地将小片段和引物片段去除。此外,由于嗅觉相关基因的长度大多分布在0.4-2.0 kb 之间(张康,2011;Gao & Wang,2014),我们构建该文库的目的是为后续克隆大草蛉嗅觉相关基因奠定基础,所以本研究中仅切取长度在500 bp 以上的片段用于后续文库的构建。
全长cDNA 文库构建是筛选全长基因的重要方法,也是进行功能基因组研究的一种经济、快速、有效的途径。它克服了传统cDNA 文库的缺点,极大促进了功能基因组的研究进程(Wiemann et al.,2003)。为此,构建全长cDNA 文库已经越来越受到研究者的重视,许多种重要生物全长cDNA 文库相继建成。然而,对于较小组织而言,因获得足够的组织材料非常困难,其文库构建相应的也十分困难。昆虫触角即属于该种情况。谷少华(2010)运用传统 SMART 技术以苜蓿盲蝽Adelphocoris lineolatus(Goeze)雌雄触角各300 根以上为提取总RNA 的原始材料构建了其触角cDNA 文 库。Zhang 等(2011)以棉铃Helicoverpa armigera(Hübner)雌雄触角各200 根以上为材料构建了其触角cDNA 文库,所用组织材料数量较大且较难获得。本试验以大草蛉Chrysopa pallens(Rambur)雌雄触角为材料,继传统SMART 技术之后运用优化的Infusion SMART 技术以雌雄触角各60 根提取总RNA,成功构建了其触角cDNA 文库,且雌雄触角cDNA 初级文库滴度达到2.1×106pfu/mL 和1.8×106pfu/mL,构建获得的文库质量较高。与其他文库构建结果相比,Infusion SMART 技术是一种比较好的全长文库构建技术,其优点体现在:一是建库效率高。与其他较常用的OLIGO-CAPPING(Maruyam et al.,1994)、CAP TRAPPER(Suzuki et al.,1997)方法及传统SMART 技术相比,Infusion SMART 技术所使用的原始材料mRNA 用量更少,省去了双链cDNA 的酶切步骤,实验快速、简单,可以从poly+RNA 开始,也可以从总RNA 开始,大约需要50 ng 就可以构建一个结果较好的文库(106pfu左右),弥补了因材料不足所带来的困扰。二是文库重组率较高。Infusion SMART 试剂盒中载体与cDNA 之间虽然是平端连接,但试验结果显示自连可能性很小,本试验中文库重组率高达93%以上。
脉翅目昆虫基因数据信息较少,对其进行全长cDNA 文库的构建、测序和功能注释为研究该物种的基因功能提供了极为宝贵的资源,为深入开展昆虫触角嗅觉相关基因的克隆及信息识别机制的研究,提供了成本低廉且信息量丰富的技术平台。从利用捕食性(或寄生性)天敌昆虫进行害虫生物防治角度,掌握了天敌昆虫嗅觉相关蛋白与外界气味化合物之间的作用机理,有利于人们对天敌昆虫行为的控制,通过释放对天敌昆虫有较强吸引作用的气味化合物于田间或温室大棚,从而提高其在某一特定空间的成功定殖性。张康(2011)通过构建中红侧沟茧蜂Microplitis mediato(Haliday)触角cDNA 文库共鉴定获得10个气味结合蛋白基因和34个气味受体基因,并结合气味结合蛋白荧光结合功能与室内行为试验共验证7种气味化合物对中红侧沟茧蜂有较强吸引作用。本试验应用In-fusion SMART 技术构建了大草蛉雌雄触角全长cDNA 文库。希望通过对cDNA 文库的筛选、克隆,大量获得大草蛉嗅觉相关蛋白基因,为研究大草蛉嗅觉识别机制奠定基础,为进一步利用大草蛉防治农林害虫提供理论依据。
References)
Benton R,Varmiee KS,Gamez C,et al.Variant lonotropic glutamate receptors as chemosensory receptors in Drosophila[J].Cell,2009,136(1):149-162.
Breer H,Raming K,Krieger J.Signal recognition and transduction in olfactory neurons[J].Biochem.Biophys.Acta,1994,1224:277-287.
Field LM,Pickett JA,Wadhams LJ.Molecular studies in insect olfaction[J].Insect Molecular Biology,2000,9:545-551.
Fujiwara H,Ishikawa H.Molecular mechanism of introduction of the hidden break into the 28S rRNA of insects:Implication based on structural studies[J].Nucleic Acids Research,1986,14(16):6393-6401.
Gambacurta A,Piro MC,Asco1i F.Cooperative homodimeric hemoglobin from Scapharca inaequivalvis:cDNA cloning and expression of the fully functional protein in E.coli[J].FEBS Letters,1993,330(1):90-94.
Gao X,Wang MQ.A cDNA library from the antenna of Monochamus alternatus Hope and binding properties of odorant-binding proteins[J].Journal of Applied Entomology,2014,138(3):233-240.
Gu SH.Exploration and functional analysis of olfactory related genes in the lucerne plant bug,Adelphocoris lineolatus(Goeze)[D].Beijing:Chinese Academy of Agricultural Sciences,2014.[谷少华.苜蓿盲蝽嗅觉相关基因的发掘及功能分析[D].北京:中国农业科学院硕士学位论文,2010]
Kaissling KE,Zack SC,Rumbo ER.Adaptation processes in insect olfactory receptors:Mechanisms and behavioral significance[J].Annals of the New York Academy of Sciences,1987,510:104-112.
Kazuo M,Sumio S.Oligo-capping:a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides[J].Gene,1994,138(1-2):171-174.
Li H,Zhang AJ,Chen LZ,et al.Construction and analysis of cDNA libraries from the antennae of Batocera horsfieldi and expression pattern of putative odorant binding proteins[J].Journal of Insect Science,2014,14(57):1-15.
Lin MZ,Chen HY,Wang SY,et al.Development of artificial diet for Chrysopa pallens larva[J].Chinese Journal of Biological Control,2007,23(4):316-321.[林美珍,陈红印,王树英,等.大草蛉幼虫人工饲料的研究[J].中国生物防治,2007,23(4):316-321]
Steinbrecht RA.Pore structures in insect olfactory sensilla:a review of data and concepts[J].International Journal of Insect Morphology and Embryology,1997,26:229-245.
Steinbrecht RA,Stankiewicz BA.Molecular composition of the wall of insect olfactory sensilla-the chitin question[J].Journal of Insect Physiology,1999,45(8):785-790.
Suzuki Y,Yoshitomo NK,Maruyama K,et al.Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library[J].Gene,1997,200(1-2):149-156.
Wiemann S,Mehrle A,Bechtel S,et al.cDNAs for functional genomics and proteomics[J].Comptes Rendus Biologies,2003,326(10):1003-1009.
Wu HH.Research on the effect of greenhouse environmental factors on Chrysopa pallens(Rambur)and Chrysopa formosa Brauer dispersal behavior[D].Beijing:Chinese Academy of Agricultural Sciences,2014.[武鸿鹄.温室环境因子对大草蛉和丽草蛉扩散行为的影响研究[D].北京:中国农业科学院硕士学位论文,2014]
Zhang K.Binding characterization of odorant-binding protein MmedOBP8 in the Microplitis mediator and the taxis responseauthentication[D].Wuhan:Huazhong Agricultural University,2011.[张康.中红侧沟茧蜂气味结合蛋白MmedOBP8 的气味结合特征及行为反应验证[D].武汉:华中农业大学硕士毕业论文,2011]
Zhang LF,Zhu JY,Yang FZ,et al.Construction and analysis of full length cDNA library from the antenna of Spodoptera exigua[J].Biotechnology,2012,4:11-14.[张丽芳,朱家颖,杨发忠,等.甜菜夜蛾触角全长cDNA 文库的构建与分析[J].生物技术,2012,4:11-14]
Zhang TT,Gu SH,Wu KM,et al.Construction and analysis of cDNA libraries from the antennae of male and female cotton bollworms Helicoverpa armigera(Hübner)and expression analysis of putative odorant-binding protein genes[J].Biochemical and Biophysical Research Communications,2011,407:393-399.
Zhang S,Zhang YJ,Su HH,et al.Cloning and tissue-specific expression of olfactory receptors in Helicoverpa armigera(Hübner)[J].Acta Entomologica Sinica,2009,52(7):728-735.[张帅,张永军,苏宏华,等.棉铃虫气味受体的克隆与组织特异性表达[J].昆虫学报,2009,52(7):728-735]
Zhang S,Zhang YJ,Su HH,et al.Cloning,expression and tissue-specific expression of cDNA encoding pheromone binding protein PBP2 in Helicoverpa armigera(Hübner)[J].Scientia Agricultura Sinica,2009,42(7):2359-2365.[张帅,张永军,苏宏华,等.棉铃虫信息素结合蛋白PBP2 cDNA 的克隆、表达与组织特异性表达[J].中国农业科学,2009,42(7):2359-2365]
Zhou JJ,Field LM,He XL.Insect odorant—binding protein:Do they offer an alternative pest control strategy?[J].Outlooks on Pest Management,2010,21(1):31-34.
