苏云金芽胞杆菌应用研究进展
2014-10-16刘明李海涛刘荣梅高继国
刘明,李海涛,刘荣梅,高继国
(1.黑龙江八一农垦大学测试中心,大庆163319;2.东北农业大学生命科学学院)
1901年日本生物学家石渡繁胤(Shigetane Ishiwatari)在研究蚕枯萎病的时候,分离出一株细菌,命名为碎倒细菌(Bacillus sotto),10年后,德国科学家贝尔林纳(Ernst Berliner)从患病的地中海粉斑螟(Ephestia kuehniella)幼虫中又分离出这种细菌,并用发现地德国图林根(Thuringia)正式命名为苏云金芽胞杆菌(Bacillus thuringiensis,Bt)[1]。Bt是一种内生芽胞的革兰氏阳性土壤细菌,在芽胞形成的同时,能产生不同形状的由cry或cyt编码的晶体蛋白,这种晶体蛋白对多种昆虫具有特异杀虫活性。
在农业害虫的防治中,最开始主要使用的是化学杀虫剂,然而,由于化学杀虫剂的使用会导致坏境污染、危害人类健康等诸多问题的产生。选择抗性品系,又会引起次级害虫的爆发[2]。经过一个世纪的发展,由于Bt杀虫剂对环境友好,对人畜无害等优势,已经成为用于害虫防治最成功的生物杀虫剂。主要介绍了苏云金芽胞杆菌cry基因资源的现状及应用情况。
1 Bt Cry毒素
Cry毒素的分类取决于其主要氨基酸的不同,1981年,Schnepf首次克隆到Bt cry基因,并证实了其杀虫活性。截至2014年1月,全球已克隆并被Bt国际命名委员会命名的杀虫晶体蛋白已经超过83类,共计771个,其中模式基因290个,包括Cry1~Cry73和Cyt1~Cyt3[3],这些Cry毒素被分为4个等级系统发育不相关的蛋白家族。从图1可以看出,近年来,Bt Cry毒素的分离鉴定克隆逐年增加,尤其是模式基因的发掘数量呈良好态势。
目前,许多公司、研究机构致力于Bt的研究以及相关的专利保护,1970年至2013年,国际上Bt的相关专利在整体呈现逐年增加趋势[4],但是,从数量与质量上,国内Bt基因专利处于劣势[5]。
Bt对不同的害虫幼虫具有特异活性,其中大部分的Bt库斯塔克(kurstaki)亚种对鳞翅目害虫具有特异活性,以色列(israelensis)亚种对双翅目害虫具有特异活性,莫氏(morrisoni)亚种对鞘翅目害虫具有特异活性。还有部分对害虫无活性,但对无脊椎动物具有活性,比如对线虫具有活性的Bt Cry5,Cry6类毒素。
目前8个不同的Cry蛋白的三维结构已经别解析 出 来 ,Cry1Aa、Cry1Ac、Cry2Aa、Cry3Aa、Cry3Ba、Cry4Aa、Cry4Ba和Cry8Ea[6-12]。通过比较发现,它们的三维结构都很相似。结构域I的7个α螺旋束与膜插入、毒素寡聚化和孔洞形成相关。结构域Ⅱ是一个由三个反向平行的β片围绕疏水内核的β棱柱,与受体识别相关。结构域Ⅲ是由2个反向平行的β片构成的β三明治结构。结构域Ⅱ、Ⅲ通过对不同害虫中肠蛋白的特异识别,与杀虫特异性相关[13]。
早期对Bt毒素杀虫机理研究发现,Bt毒素发挥杀虫作用的杀虫蛋白存在于伴胞晶体中,当害虫取食后,在害虫中肠的碱性条件下晶体溶解产生原毒素,由中肠内酶系统的作用,释放出活性毒素,由毒素与昆虫中肠内特异的受体结合而产生毒杀作用。这一杀虫机理使得某一特定的Bt毒素只对某一类昆虫有特异的毒杀作用。而哺乳动物的胃液为酸性,肠胃中也不存在与Bt毒素相结合的受体,当Bt毒素进入到哺乳动物肠胃中后,在胃液的作用下几秒钟之内全部降解。这也解释了为什么Bt对哺乳动物(酸性环境、缺少相应受体)无害、具有较窄的杀虫谱。
2 Bt杀虫剂
1927年,玛特斯(Mattes)在进行田间试验时发现Bt菌株对欧洲玉米螟(Ostrinia nubilalis)具有很好的防治效果[14],这个实验结果,最终导致了第一个Bt杀虫剂Sporeine于1938年在法国研究成功并应用。直到1961年,由美国作为商品注册的杀虫剂投放市场,我国第一个商业化的Bt杀虫剂产品“青虫菌”于1965年投入使用,用于防治大菜粉底、小菜蛾等蔬菜害虫,取得了良好的效果。
在过去50年里,世界范围内的农民都把Bt作为一种杀虫剂使用,尤其是那些有机耕田的农民从中获益不浅,因为Bt是为数不多的符合有机标准的杀虫剂。随着国际上对Bt相关的理论基础和应用基础的研究不断深入,为Bt杀虫剂的实用化发展奠定了基础。20世纪60年代,菌株的改良导致了大量新的Bt菌株替换早期的产品,杀虫效果提升达到10倍,而如今人们继续在寻找新的,更好的菌株。大多数产品都是基于芽胞-晶体制剂,来源于一些野生型的Bt菌株,比如表达Cry1Aa、Cry1Ab、Cry1Ac、Cry2Aa蛋白的Bt var.kurstaki(Btk)HD1菌株,主要用于控制很多重要作物食叶性鳞翅目害虫及森林食叶虫[14];或者含Cry1Ac蛋白的HD73菌株;含有Cry1Aa、Cry1Ba、Cry1Ca、Cry1Da的Bt var.aizawai HD137菌株,特异性的对食存储粮食的鳞翅目幼虫具有杀虫活性;含有Cry3Aa的Bt var.san diego和Bt var.tenebrionis,主要用于防治农业上的甲虫;含有Cry4A、Cry4B、Cry11A、Cyt1Aa的Bti,主要用来控制传播登革热和疟疾等人类疾病的蚊子[15]。部分Bt制剂产品在表1中列出[16]。
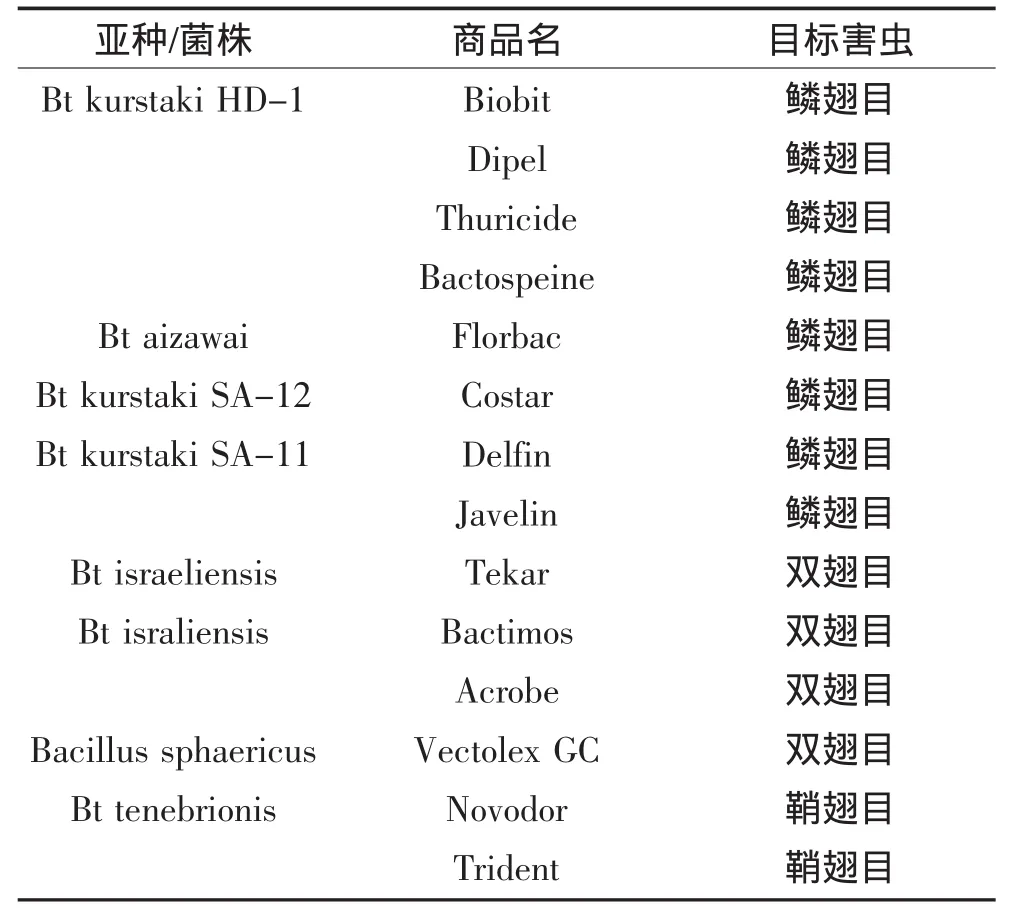
表1 基于野生型菌株的部分Bt制剂产品Table1 The topical products of Bt based on natural strains
随着Bt杀虫剂应用范围的不断扩大,基于单一野生型Bt菌株的杀虫剂已经不能满足人们的需要,这就要求继续开发杀虫谱广的Bt杀虫剂。目前,部分产品通过连接、定向转化等对已知毒素基因进行重组,形成新的菌株[17-19](Table 2),用以作为研发广谱杀虫剂的材料。

表2 基于转化结合和重组菌株的部分Bt产品Table2 The topical products of Bt based on transconjugantand recombinant strains
3 转Bt抗虫基因作物
2014年是转基因作物商业化的第19年,据国际农业生物技术应用服务组织的统计数据显示,截至2013年,全球转基因作物的种植面积达到1.752 hm2,比2012年上升500万hm2,连续17年呈增长趋势。转基因作物涉及烟草、棉花、玉米、大米、大豆、油菜、甜菜、甜椒、番茄、木瓜、南瓜、苜蓿、白杨等,通过转基因技术,以达到抗虫、抗旱、耐除草剂等目的[20]。
由于Bt对不同的害虫幼虫具有特异活性,但是对阳光照射敏感以及对钻孔的害虫杀虫活性的局限性等因素限制了Bt制剂在农业上的应用。在转基因植物中,能够保护Cry毒蛋白杀虫活性避免在紫外线的照射下降解,并且特异性的针对钻孔咀嚼的害虫,所以转基因Bt作物的研究越来越广泛。
第一个成功的例子是由比利时的植物遗传系统公司在1987年6月报道的[21]。他们用农杆菌介导法成功地将完整的cryA(b)基因和3'、5'端缺失后只保留5'端编码毒蛋白核心区的不同长度的cryA(b)基因导入烟草获得了抗烟草天蛾的转基因烟草植株。
目前最重要的Bt作物是棉花,玉米,大豆。表达Cry1Ac蛋白的商业化Bt棉花用来控制棉铃虫等鳞翅目害虫。第二代的Bt棉花,除了Cry1Ac外,表达Cry2Ab作为抗性管理机制。表达Cry1Ac蛋白的Bt玉米能有效的控制例如烟蚜夜蛾和欧洲玉米螟这样的鳞翅目害虫。下一代的商业化Bt玉米表达Cry34Ab/Cry35Ab双毒素、Cry3Bb,控制鞘翅目害虫,例如玉米根叶甲;表达Cry1A,Cry2Ab and Cry1F控制鳞翅目害虫,例如草地夜蛾[22]。2013年孟加拉国首次批准种植转基因Bt茄子。
在2013年,全球超过2 500万hm2的Bt作物,直接导致化学杀虫剂的使用明显减少[20]。表3列出了目前商业化种植的转Bt植物。

表3 商业化种植的转Bt基因植物Table3 The commercial planting of Btgene plants

续表3商业化种植的转Bt基因植物Continued Table3 The commercial planting of Btgene plants
随着Bt应用的深入,抗性的问题也随之而来。80年代,是在实验室选择压力下,部分野生型害虫种群产生了抗性[23]。在野生条件下,三种鳞翅目害虫对Bt产品产生抗性,如:印度谷螟、小菜蛾、粉纹夜蛾[24-26]。近些年,最少4种抗Bt作物的害虫已经被证明,在美国,美洲棉铃虫对转Cry1Ac棉花产生抗性[27];在波多黎各,草地贪夜蛾对转Cry1F的Bt玉米产生抗性[28]。在南非,玉米楷夜蛾对转Cry1Ab的Bt玉米产生抗性[29]。在印度,棉红铃虫对转Cry1Ac的Bt棉花产生抗性[30]。
4 展望
Bt作为目前世界上应用最广泛的微生物杀虫剂,由于避免了化学农药所带来的环境污染和抗性问题,在害虫防治制剂以及转基因材料研究中一直备受关注。尽管目前国内外已经分离克隆到了700多个Bt杀虫蛋白基因,其杀虫谱基本覆盖了所有的鳞翅目、鞘翅目、双翅目、膜翅目等害虫,但是,在Bt的研究过程中还存在着许多问题,如:每一种的Bt杀虫蛋白基因的杀虫谱却十分有限、cry基因产物表达水平有限、转基因植物中出现基因沉默以及抗性的产生等问题。因此还需通过新的试验技术手段进行分离筛选出更广谱高效的菌株、构建新的工程菌、改良现有的Cry毒蛋白、研究确定适当的Cry毒素表达水平、在同一作物中共同表达多个Cry毒蛋白、研究明白这些毒素的作用模式以及害虫对于与Cry蛋白结合的反应等方法以使这些问题得到有效的解决。
[1]Sanahuja G,Banakar R,Twyman R M,et al.Bacillus thuringiensis:a century of research,development and commercial applications[J].Plant Biotechnol J,2011,9(3):283-300.
[2]Devine G J,Furlong M J.Insecticide use:contexts and ecological conse-quences[J].Agr.Hum.Values,24(3):281-306.
[3]Crickmore N.Bacillus thuringiensis Toxin Nomenclature[EB/OL].(2013-03-07)[2014-03-31].http://www.lifesci.Sussex.ac.uk/home/Neil_Crickmore/Bt/.
[4]Swamy H M M,Asokan R.New Innovations in Bacillus Thuringiensis Research-A Review[J].International Journal of Agriculture,2013,6(4):545-550.
[5]张杰,束长龙,张春鸽.Bt杀虫基因专利保护现状与趋势[J].植物保护,2011,37(3):1-6.
[6]Li JD,Carrol J,Ellar D J.Crystal structure of insecticidal d-endotoxin from Bacillus thuringiensis at 2.5 A resolution[J].Nature,1991,353(6347):815-821.
[7]Grochulski P,Masson L,Borisova S,et al.Bacillus thuringiensis CryIA(a)insecticidal toxin:crystal structure and channel formation[J].JMol Biol,1995,254(3):447-464.
[8]Morse R J,Yamamoto T,Stroud R M.Structure of Cry2Aa suggests an unexpected receptor binding epitope[J].Structure,2001,9(5):409-417.
[9]Galitsky N,Cody V,Wojtczak A,et al.Structure of the insecticidal bacterial d-endotoxin Cry3Bb1 of Bacillus thuringiensis[J].Acta Crystallogr D,2001,57(8):1101-1109.
[10]Boonserm P,Davis P,Ellar D J,et al.Crystal Structure of the Mosquito-larvicidal Toxin Cry4Ba and Its biological implications[J].JMol Biol,2005,348(2):363-382.
[11]Boonserm P,Mo M,Angsuthanasombat C,et al.Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thur-ingiensis at a 2.8-angstrom resolution[J].JBacteriol,2006,188(9):3391-3401.
[12]Guo S,Ye S,Liu Y,et al.Crystal structure of Bacillus thuringiensis Cry8Ea1:An insecticidal toxin toxic to underground pests,the larvae of Holotrichia parallela[J].JStruct Biol,2009,168(2):259-266.
[13]Bravo A,Gill SS,Soberón M.Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control[J].Toxicon,2007,49(4):423-435.
[14]Husz B.Field experiments on the application of Bacillus thuringiensis against the corn borer Int Corn Borer Invest[J].SciRep,1930,3:91-98.
[15]Soberón M,Gill SS,Bravo A.Signaling versus punching hole:how do Bacillus thuringiensis toxins kill insect midgut cells[J].Cell Mol Life Sci,2009,66(8):1337-1349.
[16]Kaur S.Molecular approaches towards development of novel Bacillus thuringiensis biopesticides[J].World J Microbiol Biotechnol,2000,16(13):781-793.
[17]González JM Jr,Brown B J,Carlton B C.Transfer of Bacillus thuringiensis plasmids coding for deltaendotoxin among strains of B.thuringiensis and B.cereus[J].Proc Natl Acad SciUSA,1982,79(22):6951-6955.
[18]Kronstad JW,Schnepf H E,Whiteley H R.Diversity of locations for Bacillus thuringiensis crystal protein genes[J].JBacteriol,1983,154(1):419-428.
[19]Baum J A,Johnson T B,Carlton B C.Bacillus thuringiensis:Natural and recombinant bioinsecticide products[J].Methods Biotechnol,1995,5:189-209.
[20]James C.Global Status of Commercialized Biotech/GM Crops:2013[R].Metro Manila:ISAAA Brief,2014.
[21]Vaeck M,Reynaerts A,Hofte H,et al.Transgenic plants protected from insect attack[J].Nature,1987,328(2):33-37.
[22]Christou P,Capell T,Kohli A,et al.Recent developments and future prospects in insect pest control in transgenic crops[J].Trends Plant Sci,2006,11(2):302-308.
[23]Veloorvalappil N J,Robinson B S,Sailas B.An Overview on the Crystal Toxins from Bacillus thuringiensis[J].Advances in Microbiology,2013,3(5),462-472.
[24]McGaughey W H.Insect resistance to the biological insecticide Bacillus thuringiensis[J].Science,1985,229(4709):193-195.
[25]Tabashnik B E.Evolution of resistance to Bacillus thuringiensis[J].Annu Rev Entomol,1994,39,47-49.
[26]Janmaat A F,Myers JH.Rapid evolution and the cost of resistance to Bacillus thuringiensis in greenhouse populations of cabbage loopers,Tricoplusia ni.[J].Proc Biol Sci,2003,270(1530),2263-2270.
[27]Tabashnik B E,Gassman A J,Crowdwer D W,et al.Insect resistance to Bt crops:evidence versus theory[J].Nat Biotechnol,2008,26(2):199-202.
[28]Storer N P,Babcock JM,Schlenz M,et al.Discovery and characterization of field resistance to Bt Maize:Spodoptera frugiperda(Lepidoptera:Noctuidae)in Puerto Rico[J].JEcon Entomol,2010,103(4):1031-1038.
[29]Van Rensburg J B J.First report of field resistance by stem borer Busseola fusca(Fuller)to Bt-transgenic maize[J].SAfr JPlant Soil,2007,24:147-151.
[30]Bagla P.Hardy cotton-munching pests are latest blow to GM crops[J].Science,2010,327:1439.
