尖叶拟船叶藓光系统Ⅱ光合荧光特性、活性氧代谢与耐脱水生理生态适应的关系
2014-09-19杜晓濛田向荣李朝阳李鹄鸣
杜晓濛,李 菁, 2,田向荣, 2,*,李朝阳,李鹄鸣,2
(1. 湖南吉首大学生物资源与环境科学学院, 吉首 416000; 2. 植物资源保护与利用湖南省高校重点实验室, 吉首 416000)
自20世纪70年代以来,以山墙藓(Tortularuralis)等为材料的研究证实藓类植物具有极强的脱水耐性[1- 3], 其典型的变水(poikilohydric)特性使它们能够在特殊生境(如极度干旱地区)和特殊基质(如维管植物叶面)生长,其特殊的生理机制使其具有极强的适应干旱环境的能力[4]。而藓类植物体内的水分关系及耐受能力决定了其种群的分布范围和生存能力,即湿生藓类耐旱性较弱,干旱环境下生长的藓类具有较强的耐旱性。此种生理生态适应性一方面与其栖息地水势变化息息相关,另一方面则与生理代谢途径的不同密不可分[5]。
然而即使长期生活在同样干旱或湿润环境下的不同藓属,其脱水耐受能力和生理生化响应也存在一定差别。国内对沙漠结皮藓类,如刺叶赤藓(SyntrichiacaninervisMitt) 和土生对齿藓(Didymodonvinealis)的系列研究业已证明同为干旱区藓类其耐受性存在明显差距[6- 11],其中对土生对齿藓(Didymodonvinealis)与结皮共生的真藓(Bryumargenteum)进行光合特性分析发现两种藓类光合特性在脱水过程中差异明显[11]。而Minibayeva和Beckett的研究也指出湿润环境下生长的不同藓类在产生氧迸发(oxidative burst)和抗氧化能力方面仍存在明显差异[12]。
尖叶拟船叶藓(Dolichomitriopsisdiversiformis)系船叶藓科(Lembophyllaceae)拟船叶藓属植物,为东亚特有种,数量少,且分布狭窄。以往研究发现水分条件是限制其分布的主要因子之一[13]。由于环境日益干旱,在自然和人为因素的双重影响下,其天然种群不断衰减,分布范围越来越窄,濒临灭绝[14]。为保护这一濒危物种,本研究试图以广布湿生藓类—湿地匐灯藓(Plagiomniumacutum)为比较材料,从生理生态学角度探讨变水下尖叶拟船叶藓叶绿素光合荧光作用、氧代谢及其保护系统酶活性的变化与生态适应的关系。
1 材料和方法
1.1 实验材料
供实验的尖叶拟船叶藓采自贵州省梵净山海拔1680米的常叶阔叶林内[15],湿地匐灯藓采自湖南省吉首市西郊德夯风景区小溪畔,材料采集后在室温下以自然光源作保湿培养。
1.2 实验方法
选择硅胶快速脱水法作为本实验脱水方式和方法[16]。以未经处理为对比,脱水时间梯度分别为30 min(D30)、60 min(D60)、120 min(D120)、240 min(D240);脱水不同时间后,将干燥的藓类放置用湿滤纸覆盖的培养皿中进行复水处理,复水时间梯度为30 min(R30)、60 min(R60)、120 min(R120)、240 min(R240)。
1.2.1 含水量的测定
以干重为基础计算含水量,单位为g H2O/g 干重。每个处理时间点进行3次重复[17]。
1.2.2 PSⅡ叶绿素荧光参数测定
用PAM- 2500便携式调制叶绿素荧光仪和IMAGING-PAM 叶绿素荧光成像仪(WALZ,德国)分别绘制不同脱水与复水处理下的荧光光响应曲线(ETR-PAR)和荧光成像图,并测定最大光化学效率(Fv/Fm)、光化学量子效率Y(Ⅱ)、光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)等叶绿素荧光参数[18]。
1.2.3 活性氧的测定
1.2.4 抗氧化系统酶和非酶测定
超氧化物歧化酶(SOD)活性以抑制0.1个吸收值为1U, 过氧化氢酶(CAT)活性以每min光吸收值减少0.1为1U[21]。抗坏血酸过氧化物酶(APX)以每min光吸收值减少0.1为1U[22]。抗坏血酸(AsA)含量以mg/g 干重来表示[23]。蛋白质含量以牛血清蛋白为标准[24]。
1.2.5 数据统计分析
采用Excel2003和SPSS 13. 0 进行均值、标准差和差异显著性分析。
2 结果与分析
2.1 脱水与复水过程中尖叶拟船叶藓和湿地匐灯藓含水量的变化
脱水与复水过程中两种藓类植物含水量变化剧烈(表1)。正常情况下,两种藓类的含水量均保持高水平,而D240时其含水量均迅速下降至最低值,均降至低于0.05 g H2O/g 干重以下,但D60以后含水量再无显著差别(P>0.05)。脱水后迅速复水过程中,两种藓类植物的含水量均能迅速恢复,R120时基本接近对照水平,无论绝对值和变化幅度变化并无明显差别(P>0.05)。
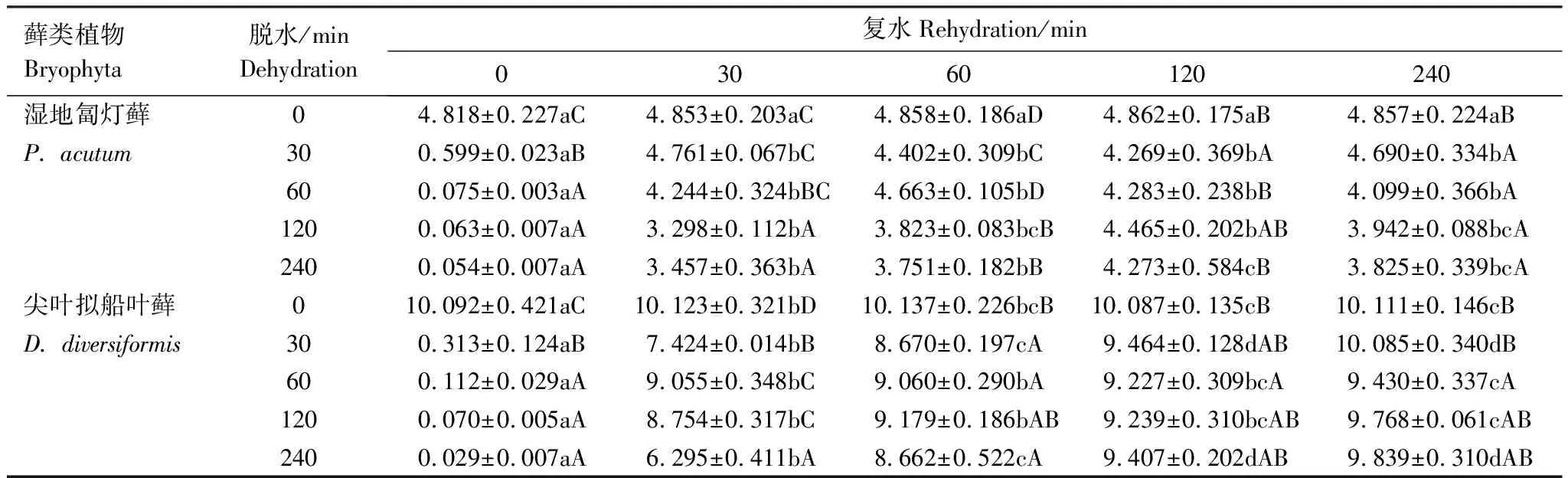
表1 脱水与复水过程中尖叶拟船叶藓与湿地匐灯藓的含水量(g H2O/g 干重)变化
同时,两种藓类植物含水量的差异明显(表1),尖叶拟船叶藓和湿地匐灯藓两种藓类的含水量分别为10.092 g H2O/g 干重和4.818 g H2O/g 干重。虽然同为湿生藓类,但前者的含水量为后者的2.09倍。由表1还可知两种藓类植物的控水能力差异也较明显。尖叶拟船叶藓的失水和复水幅度都比湿地匐灯藓大。脱水过程中,D120时尖叶拟船叶藓的含水量为对照的0.70%,而湿地匐灯藓则为其对照的1.3%,是湿地匐灯藓的1.7倍。复水过程中,无论脱水程度如何,尖叶拟船叶藓在R60时,含水量可恢复到其对照含水量的90%上下,而湿地匐灯藓均须在R120后才达到其对照的90%上下,后者明显慢于前者。
2.2 PSⅡ荧光光响应曲线(ETR-PAR)
脱水胁迫加剧过程中,尖叶拟船叶藓的抑制光强迅速降低,从对照的1375 μmol m-2s-1,下降到D30时105 μmol m-2s-1,D60时极微弱光也能造成抑制;相反,湿地匐灯藓在D30时的抑制光强仍达249 μmol m-2s-1(图1)。两种藓类在正常条件下所能达到的光合电子传递速率(ETR)都接近50μmol m-2s-1,D30时尖叶拟船叶藓ETR仅为5 μmol m-2s-1,D60时,ETR已无法检测;而湿地匐灯藓D60时仍可以达到10 μmol m-2s-1(图1)。复水后,两种藓类的抑制光强和ETR均逐渐恢复,抑制光强恢复迅速,R120时均可都恢复到正常水平。说明含水量较低时PSⅡ的光电转换能力会下降,而复水后可逐渐恢复。
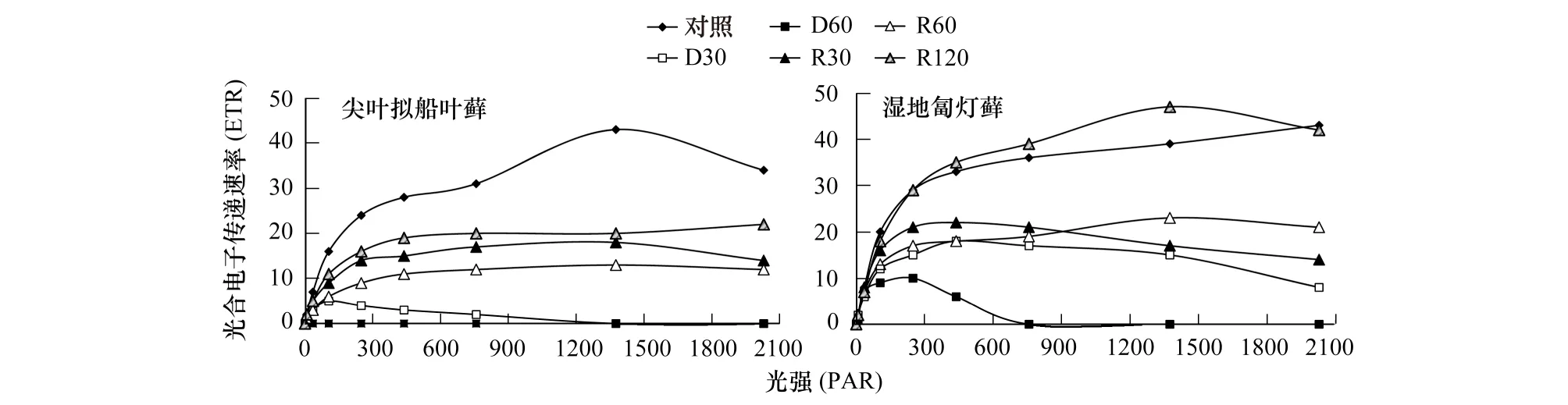
图1 脱水与复水过程中尖叶拟船叶藓与湿地匐灯藓的叶绿素荧光光响应曲线
2.3 脱水与复水过程中尖叶拟船叶藓和湿地匐灯藓PSⅡ叶绿素荧光参数变化
2.3.1 Fv/Fm荧光成像整体变化
尖叶拟船叶藓(图2 A—F)和湿地匐灯藓(图2 a—f)在脱水过程中荧光逐渐暗淡,株体收缩、卷曲,从蓝变绿至黄,表示Fv/Fm逐渐减小;复水过程中,叶片张开,荧光逐渐恢复亮度,蓝色面积逐渐增大,表示Fv/Fm逐渐增大。脱水过程中,尖叶拟船叶藓Fv/Fm低于湿地匐灯藓,R30时湿地匐灯藓已大面积恢复蓝色,而尖叶拟船叶藓仅有局部恢复绿色,至R120时,湿地匐灯藓已完全恢复蓝色,而尖叶拟船叶藓仅有部分区域恢复蓝色,表示从整体上看尖叶拟船叶藓在复水过程中Fv/Fm的恢复进程明显慢于湿地匐灯藓。
2.3.2最大光化学效率(Fv/Fm)和光化学量子效率Y(Ⅱ)
脱水与复水过程中两种藓类的Fv/Fm值与含水量变化的趋势一致,呈先降后升的趋势(图3)。尖叶拟船叶藓D60达到最低值0.202,较正常水平下降了68%;而复水R120时Fv/Fm值仅为0.51,仍未恢复到正常水平。湿地匐灯藓Fv/Fm同比明显高于尖叶拟船叶藓。Y(Ⅱ)值的变化与Fv/Fm值的走势相似(图3),其中尖叶拟船叶藓D60时值达到最低,比对照下降了92.4%,与对照组相比差异显著(P<0.05),复水至R120仍未恢复到原有水平。说明尖叶拟船叶藓由于PSⅡ受损,光合效能恢复较慢。
2.3.3光化学淬灭(qP)和非光化学淬灭(NPQ)
随着脱水时间的延长,尖叶拟船叶藓的qP值迅速下降,由对照的0.658降到D60时的0.282(图3)。其中D30与D60并无显著差异(P>0.05),可认为D30时即已基本降至稳定的低值。复水后qP迅速恢复,R30时已恢复到原有水平。湿地匐灯藓变化幅度明显小于尖叶拟船叶藓。NPQ值的变化与qP值变化相反(图3),其中尖叶拟船叶藓D60时NPQ最大值才0.486,为对照的2倍多。复水过程中,NPQ迅速下降,D30后与对照的水平已基本无差别(P>0.05)。而湿地匐灯藓D60最大值达到为0.835,整个脱水和复水过程NPQ都要高于尖叶拟船叶藓。说明脱水胁迫下湿地匐灯藓可以更多地依靠热耗散能力来保护其光合机构,而尖叶拟船叶藓的此种光合机构保护能力则相对较弱。

图2 脱水与复水过程中尖叶拟船叶藓(A—F)与湿地匐灯藓(a—f)Fv/Fm荧光成像整体变化
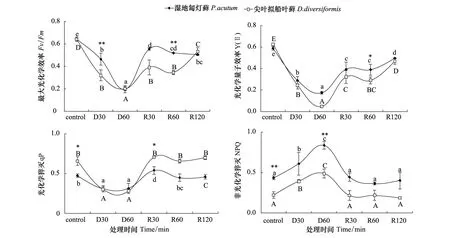
图3 快速脱水与复水过程中尖叶拟船叶藓与湿地匐灯藓最大光化学效率、光化学量子效率、 光化学淬灭和 非光化学淬灭的变化
2.4 脱水与复水过程中尖叶拟船叶藓和湿地匐灯藓活性氧代谢的变化
2.4.1 活性氧自由基
2.4.2 抗氧化酶活性的变化
活性氧的变化诱导两种藓类的SOD、CAT、APX活性均表现相同走势。其中尖叶拟船叶藓SOD活性在D60达到最大值202.1 U/mg protein,较最初的71.2 U/mg protein上升了2.8倍;R30时活性则下降到20 U/mg protein,复水后随时间延长,SOD酶活性差异并不显著,但低于对照原有水平(图5)。CAT和APX活性分别在D60 min时出现最大值2.7 U/mg protein和3.0 U/mg protein,而复水过程中,CAT和APX虽然在R30 min时恢复到与对照无显著差别水平(P>0.05),但随着复水时间延长,其活性明显下降,且尖叶拟船叶藓始终低于湿地匐灯藓(图5)。说明相对水分充沛小生境的尖叶拟船叶藓的抗氧化酶系统反应更为敏感。
2.4.3 抗坏血酸(AsA)含量
正常情况下,湿地匐灯藓的AsA含量(2.694 mg/g 干重)比尖叶拟船叶藓(1.189 mg/g 干重)高出50%(图5)。快速脱水与复水过程中,虽然两种藓类AsA含量呈现一致变化趋势,但尖叶拟船叶藓的AsA含量大都低于湿地匐灯藓近1倍。在D30时湿地匐灯藓比对照下降了71%,而尖叶拟船叶藓的下降幅度则不足48%;复水过程中,尖叶拟船叶藓在R30时即已恢复到原有对照水平,而湿地匐灯藓则到R120时才恢复到与对照一致的水平。而尖叶拟船叶藓的AsA含量始终低于湿地匐灯藓,说明其由SOD歧化产生的多余H2O2的清除可能更多依赖于酶促系统。
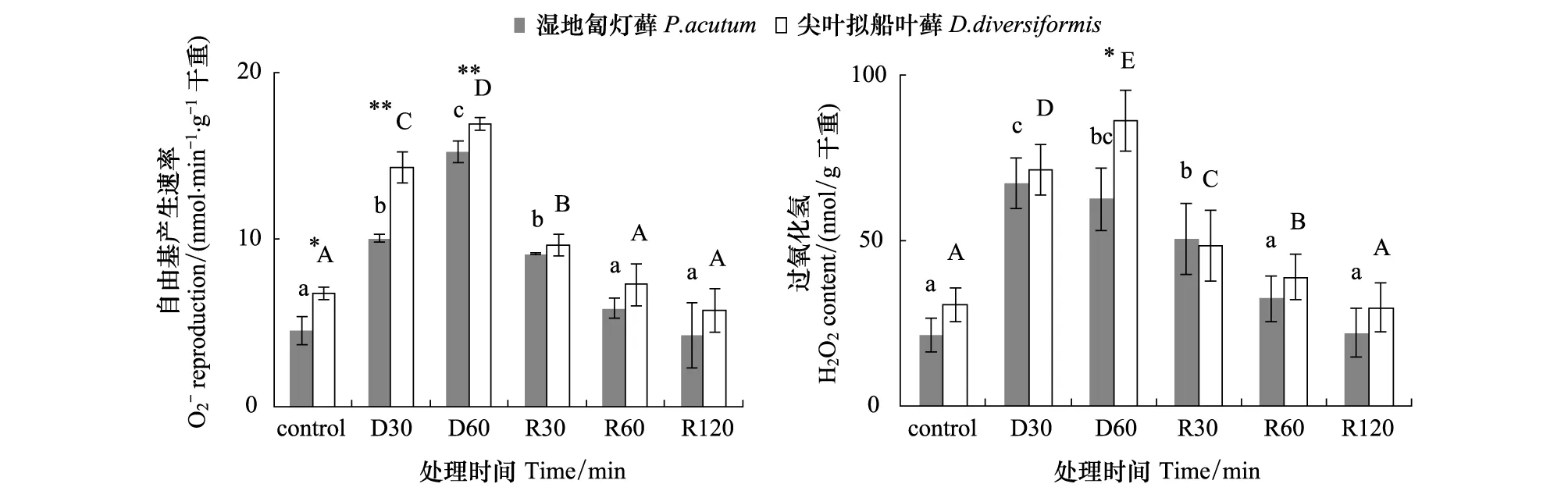
图4 快速脱水与复水过程中尖叶拟船叶藓与湿地匐灯藓超氧阴离子产生速率和H2O2含量的变化

图5 快速脱水与复水过程中尖叶拟船叶藓与湿地匐灯藓超氧化物歧化酶、过氧化氢酶、抗坏血酸过氧化物酶活性和抗坏血酸含量的变化
3 讨论与结论
3.1 含水量
藓类植物剧烈的变温和变水效应说明了其具有超强的脱水耐性[25]。本文的含水量实验结果表明尖叶拟船叶藓和湿地匐灯藓均具备一定的耐旱能力,而胁迫下保水能力的不同是造成两种藓类含水量变化的主要原因[2]。根据本文结果,不难看出尖叶拟船叶藓的正常含水量要明显高于湿地匐灯藓,换言之,其对环境水分的要求要高于湿地匐灯藓。同时,在脱水与复水的变水过程中,尖叶拟船叶藓更易失水也更易得水,说明保水能力要明显弱于湿地匐灯藓。
3.2 光合荧光特性
对不同大气温度、藓体含水量及光照条件下尖叶拟船叶藓光合速率研究中,我们已发现光合速率与光照强度、大气温度及藓体含水量之间关系密切,其在230 ETR反映实际光强条件下的表观电子传递速率,以其与有效辐照(PAR)建立的荧光光响应曲线可以很好的说明植物光合速率及光系统光电转换效率[26]。本文ETR-PAR的变化很好地说明了尖叶拟船叶藓PSⅡ对脱水胁迫较为敏感,表现在脱水过程中抑制光强和电子传递速率的快速下降[27],即使复水至R120时尖叶拟船叶藓ETR的绝对值仍仅有对照水平50%,暗示其PSⅡ的捕光复合体修复要快于PSⅡ中心色素,即尖叶拟船叶藓PSⅡ的反应中心色素(P680)可能是其受脱水损伤的主要部位。 Fv/Fm和Y(Ⅱ)反映 PSⅡ反应中心的光能转换效率,其变化可直接体现植物受胁迫的情况,qP则表示光合速率快慢与光合碳同化等光合化学反应密切相关[28]。本文中Fv/Fm,Y(Ⅱ)和qP均呈现在脱水时迅速下降,而在复水时渐次上升的趋势,说明两种藓类的耐脱水能力主要体现在复水后的修复能力[29]。与湿地匐灯藓对比,尖叶拟船叶藓Fv/Fm和Y(Ⅱ)在复水过程中恢复明显较慢,相反,其qP恢复迅速和复水后的高水平则体现了对于光能分配中优先光合碳同化[30]。NPQ反应热耗散的变化,是保护PSⅡ的重要机制,一般认为PSⅡ热耗散可能是通过叶黄素循环来实现[31]。脱水过程中NPQ的迅速升高,说明尖叶拟船叶藓和湿地匐灯藓可以通过启动热耗散来消耗掉过剩的光激发能,有利于其在脱水伤害中为稳定PSⅡ[32]。而结合尖叶拟船叶藓复水后Fv/Fm,Y(Ⅱ)的缓慢上升与qP/NPQ的比例迅速由2∶8转换为8∶2来看,能够说明尖叶拟船叶藓复水过程中NPQ值低于湿地匐灯藓可能与其叶黄素循环效能恢复较慢有关。 通过将尖叶拟船叶藓与同为湿生藓类的湿地匐灯藓进行光合荧光特性和抗氧化系统的比较,不难看出尖叶拟船叶藓在光合机构稳定性和抗氧化能力上均明显弱于湿地匐灯藓。基于此,尖叶拟船叶藓受到环境水分子限制的原因主要有两个 PSⅡ的反应中心色素(P680)对脱水伤害较为敏感;复水修复过程中抗氧化保护系统的保护能力偏弱。换言之,尖叶拟船叶藓耐脱水光保护机制的匮乏及其抗氧化系统的不完善可能是其面临濒危的主要生理因素。基于广布湿生藓类湿地匐灯藓和尖叶拟船叶藓两种湿生藓类的比较,以及与对湿度条件要求较低的金发藓的比较[16],还可以认为,藓类植物对水分环境的适应可能直接取决于光合机构的耐受性和抗氧化系统能力的强弱。 : [1] Dhindsa R S. Glutathione status and protein synthesis during drought and subsequent rehydration inTortularuralis. Plant Physiology, 1987, 83(4): 816- 819. [2] Oliver M J, Velten J, Wood A J. Bryophytes as experimental models for the study of environmental stress tolerance:Tortularuralisand desiccation-tolerance in mosses. Plant Ecology, 2000, 151(1): 73- 84. [3] Zsolt C, Proctor M C F, Tuba Z. Chlorophyll fluorescence during drying and rehydration in the mossesRhytidiadelphusloreus(Hedw.) warnst.,Anomodonviticulosus(Hedw.) Hook. & Tayl. andGrimmmiapulvinata(Hedw.)sm. Annals of Botany, 1999, 84(2): 235- 244. [4] Proctor M C F, Oliver M J, Wood A J, Alpert P, Stark L R, Cleavitt N L, Mishler B D. Desiccation-tolerance in bryophytes: a review. The Bryologist, 2007, 110(4): 595- 621. [5] Oliver M J, Velten J, Mishler B D. Desiccation tolerance in bryophytes: a reflection of the primitive strategy for plant survival in dehydrating habitats? Integrative and Comparative Biology, 2005, 45(5): 788- 799. [6] Wei M L, Zhang Y M, Zhang J. Microscopic and Submicroscopic Structure of Leaf Cells of Syntrichia caninervis Mitt. In Biological Soil Crusts. Journal of Desert Research, 2009, 29 (3): 493- 498. [7] Li X R, Zhang Y M, Zhao Y G. A Study of Biological Soil Crusts Recent Development Trend and Prospect. Advances in Earth Science, 2009, 24(1): 11- 24. [8] Shi Y, Yang X J, Zhao X, Li X R. Responses of signal transduction substances ofDidymodonvinealisandBryumargenteumin biological soil crust to drought stress. Chinese Journal of Ecology, 2012, 31(5): 1136- 1142. [9] Wei M L, Zhang Y M. Effects of Dehydration on Photosynthetic Pigment Content and Chlooplast Ultrastructure of Syntrichia caninervis in Biological Soil Crusts. Journal of Desert Research, 2010, 30(6): 1311- 1318. [10] Zheng Y P, Zhao J C, Zhang B C, Li L, Zhang Y M. Advances on Ecological Studies of Algae and mosses in Biological Soil Crust. Chinese Bulletin of Botany, 2009, 44(3): 371- 378. [11] Chen C Y, Li X R, Zhao X. Photosynthetic properties of two crust mosses during dehydration. Chinese Journal of Ecology, 2012, 31(12): 3064- 3069. [12] Minibayeva F, Beckett R P. High rates of extracellular superoxide production in bryophytes and lichens, and an oxidative burst in response to rehydration following desiccation. New Phytologist, 2001, 152(2): 333- 341. [13] Li Y J, Li J, Chen J, Huang G W. Response ofDolichomitriopsisdiversiformisphotosynthesis to light, air temperature and plant water content. Chinese Journal of Applied Ecology, 2004, 15(3): 391- 395. [14] Chen G X, LI J, Chen J, Zhu J Y, Chen J R, Liu Y D. A preliminary study on the community environment ofDolichmitriopsisdiversiformis(Mitt) Nog. In Mt. Fanjing, Guizhou, China. Guizhou Science, 2001, 19(4): 81- 84. [15] Chen J, Liu B, Li J, Jiang D S. Relationship between the distribution and the environmental factors in epiphytic mossDolichomitriopsisdiversiformis. Journal of Jishou University (Natural Sciences Edition), 2006, 27(5): 85- 88. [16] Du X M, Li J, Tian X R, Li Z Y. Reactive Oxygen Species Metabolism and Lipid peroxidation ofPolytrichumcommuneandPlagiomniumacutumduring Fast Drying and rehydration. Acta Botanica Boreali -occidentalia Sinica, 2012, 32(11): 2279- 2285. [17] Vertucci C W, Roos E. Theoretical basis of protocols for seed storage II. The influence of temperature on optimal moisture levels. Seed Science Research, 1993, 3(3): 201- 203. [18] Du X M, Tian X R, Li J, Shi J X. Comparison on Alternation of Chlorophyll Fluoroscence BetweenPplytrichumcommuneandPlagiomniumacutumDuring Dehydration and Rehydration. Life Science Research, 2012, 16(6): 521- 525. [19] Bors W, Lengfelder E, Saran M. Oxidation of hydroxylamine to nitrite as an assay for the combined presence of superoxide anions and hydroxyl radicals. Biochemical and Biophysical Research Communications, 1977, 75(4): 973- 979. [20] Patterson B D, Macrae E A, Ferguson I B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Analytical Biochemistry, 1984, 139(2): 487- 492. [21] Jiang M Y, Zhang J H. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiology, 2001, 42(11): 1265- 1273. [22] Donahue J L, Okpodu M C, Cramer C L, Grabau E A, Alscher R G.. Responses of antioxidants to paraquat in pea leaves (Relationships to resistance). Plant Physiology, 1997, 113(1): 249- 257. [23] Mishra N P, Mishra R K, Singhal G S. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiology, 1993, 102(3): 903- 910. [24] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 1976, 72(1/2): 248- 254. [25] Oldenhof H, Wolkers W F, Bowman J L, Tablin F, Crowe J H. Freezing and desiccation tolerance in the mossPhyscomitrellapatens: An in situ fourier transform infrared spectroscopic study. Biochimica et Biophysica Acta, 2006, 1760(8): 1226- 1234. [26] Genty B, Briantais J M, Baker N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta, 1989, 990(1): 87- 92. [27] Fu W G, Li P P, Wu Y Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Scientia Horticulturae, 2012, 135: 45- 51. [28] Lichtenthaler H K, Miehé J A. Fluorescence imaging as a diagnostic tool for plant stress. Trends in Plant Science, 1997, 2(8): 316- 320. [29] Oliver M J, Cushman J C, Koster K L. Dehydration tolerance in plants. Methods in Molecular Biology, 2010, 639: 3- 24. [30] Misra A N, Srivastava A, Strasser R J. Utilization of fast chlorophyll a fluorescence technique in assessing the salt/ion sensitivity of mung bean and Brassica seedlings. Journal of Plant Physiology, 2001, 158(9): 1173- 1181. [31] Dugald C D, Beadle C L. Xanthophyll-cycle dynamics and rapid induction of anthocyanin synthesis in Eucalyptus nitens seedlings transferred to photoinhibitory conditions. Journal of Plant Physiology, 2005, 162(1): 37- 46. [32] Zhao L Y, Deng X P, Shan L. The Response Mechanism of Active Oxygen Species System to Droght Stress. Acta Botanica Boreali - occidentalia Sinica, 2005, 25(2): 413- 418. [34] Fu S L, Zhou Y B, He X Y, Chen W. Effects of drought stress on photosythesis physiology ofPopuluspseudo-sinonii. Chinese Journal of Applied Ecology, 2006, 17(11): 2016- 2019. [33] Pompelli M F, Barata-Luís R, Vitorino H S, Gonçalves E R, Rolim E V, Santos M G, Almeida-Cortez J S, Ferreira V M, Lemos E E, Endres L. Photosynthesis, photoprotection and antioxidant activity of purging nut under drought deficit and recovery. Biomass and Bioenergy, 2010, 34(8): 1207- 1215. [35] Charron A J, Quatrano R S. Between a Rock and a dry place: the water-stressed moss. Molecular Plant, 2009, 2(3): 478- 486. [36] Xu Z F, Luo G H, Wang A G. Photoinhibition of Photosynthesis and the Metabolism of Active Oxygen in Photosynthetic Organs. Plant Physiology Journal, 1999, 35(4): 325- 331. [37] Pospíšil P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. Biochimica et Biophysica Acta, 2012, 1817(1): 218- 231.3.3 活性氧代谢与抗氧化系统酶活性
3.4 尖叶拟船叶藓受水分因子限制的生理生态原因
