兴安落叶松林土壤的无机态氮及氮矿化速率1)
2014-09-18刘星岑魏江生周梅包翔赵鹏武
刘星岑 魏江生 周梅 包翔 赵鹏武
(内蒙古农业大学,呼和浩特,010018)
牛 香
(中国林业科学研究院森林生态环境与保护研究所)
土壤氮矿化被认为是土壤氮影响植物生长和陆地植被生产力的关键[1]。研究土壤中氮素的转化过程有助于改进提高氮素可利用性的技术措施,对于了解森林生态系统生产力、营养循环以及森林的经营管理有重要的实践意义[2]。自19世纪初,土壤氮矿化已被认为是土壤氮循环核心和控制植物有效氮的主要过程。随着目前气候的急剧变化、大气氮沉降的增加以及对陆地生态系统碳循环深入的研究,土壤氮矿化及其影响因素的研究也倍受关注[3]。一方面考虑气候变化后,土壤氮矿化如何影响植被生产力和土壤有机碳动态[4];另一方面,考虑气候变化和CO2体积分数增加后,土壤氮矿化是否改变,对植被生产力和土壤有机碳又将产生什么影响,进而对陆地生态系统碳汇功能将产生什么影响[5-7]。基于此,土壤氮矿化及其影响因素又成为目前陆地生态系统碳氮循环研究的热点。欧、美国家在森林氮矿化方面的研究较为系统,在探讨不同森林生态系统净氮矿化过程的同时,加强了氮总矿化潜力、碳氮耦合效应及氮矿化过程对干扰响应等方面的研究[8-11]。由于气候、土壤类型、森林植被等众多因素均不同程度地影响着氮矿化过程,因而不同类型的森林生态系统的氮矿化有其各自的特点,尚未形成统一的矿化模式。
大兴安岭地处东北亚环境敏感的寒温带地区,森林资源丰富,由于其屏障作用使南下的寒潮流速减慢,对东北和我国乃至东亚的气候都有着一定的调节作用。兴安落叶松是大兴安岭森林的优势树种,也是我国北方林的代表树种,其森林面积、蓄积量及对气候的调节在我国生态系统中占有重要地位。大兴安岭独特的低温高湿的环境使土壤有机质分解速度缓慢,林下土壤具有深厚的腐殖质层,随着气候的变化,有机质的分解速度及土壤氮矿化都将受到极大的影响。因此,深入研究兴安落叶松林下不同植被类型土壤无机态氮及氮矿化速率变化规律,对于提高兴安落叶松森林生产力及固碳能力具有重要意义。
1 试验地概况
研究区设在内蒙古大兴安岭森林生态系统国家野外科学观测研究站(以下简称大兴安岭森林生态站)试验区内,试验区为大兴安岭北部冷湿缘地貌强烈发育区,属寒温带湿润气候区,≥10℃年积温为1403℃,年平均气温为-5.4℃,最低气温-50℃,最高为40℃,年降水量为450~550 mm,60%集中在7、8月份。土壤属棕色针叶林土,腐殖质质量分数10%~30%,土壤 pH 值为 4.5~6.5,滞水性强,使土壤在生长季处于湿润状态,灰化现象不明显。河谷分布有草甸土和沼泽土,土层母质多残积物,基岩以花岗岩和玄武岩为主。盐基饱和度一般在50%~70%,交换性酸 0.01~0.6 mmol·g-1。从根河以南到大兴岭南部是岛状冻土区。主要侵蚀力为寒冻分化、冻胀侵蚀和雪蚀,主要冰缘地貌有石海、石河、泥炭丘、冰丘、雪蚀凹地等。该地区的植被主要为兴安落叶松林,其中草类—兴安落叶松林(Grass-Larix gmelini forest)、赤杨—兴安落叶松林(Alnus-Larix gmelini forest)、杜香—兴安落叶松林(Ledum Sphagnum-Larix gmelini forest)、柴桦—兴安落叶松林(Fruitcosa-Larix gmelini forest)是该地区典型的4种林型,占地面积较广。具体分布情况如下:①草类—兴安落叶松林(Grass-Larix gmelini forest)(简称草类林),分布在坡度20°以下的缓坡、山麓等地,而且多生长在阳坡,此林型面积较小,生产力较高,土壤为棕色针叶林土,湿度较低。②赤杨—兴安落叶松林(Alnus-Larix gmelini forest)(简称赤杨林),分布在大兴安岭北坡,土层浅薄,土壤为棕色针叶林土,赤杨为固氮植物,是大兴安岭兴安落叶松林寒温性针叶林亚带的代表。③杜香—兴安落叶松林(Ledum Sphagnum-Larix gmelini forest)(简称杜香林),分布在阴坡、半阴坡的坡中、坡上,是大兴安岭兴安落叶松林的主要林型之一,生境属于冷湿型,枯枝落叶层分解不良,土层较浅薄,土壤多为潜育泥炭化暗棕壤,永冻层的融解层较浅。④柴桦—兴安落叶松林(Fruitcosa-Larix gmelini forest)(简称柴桦林),分布于河谷一带,生境属水湿型,土壤水分经常饱和,下有多年冻土,土层中等厚度,具有泥炭层,潜育化程度较高。4种林型土壤的基本状况见表1。
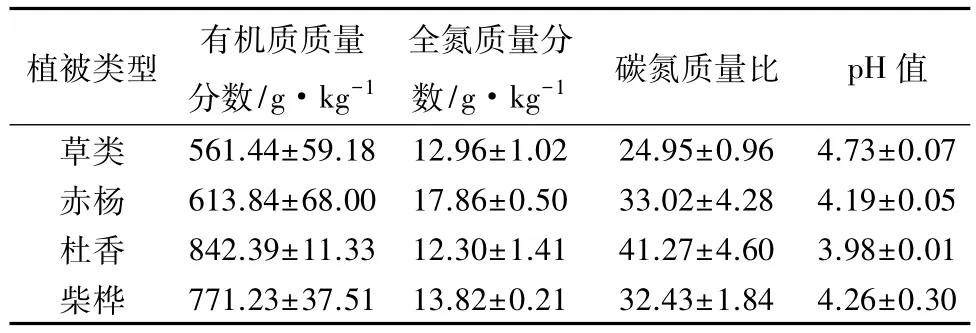
表1 4种林型土壤基本状况
2 研究方法
2.1 取样方法
本研究采用封顶埋管法来估测氮素矿化速率,在上述4种最典型的兴安落叶松林中分别随机布点,共布设60个采样点。采用野外封顶埋管法连续取样,在每一培养点将两根内径6 cm、长15 cm的PVC管打入土壤。小心取出两管后,一管土壤用于土壤分析,另一管管顶加盖,管底用纱布封口,埋回原位进行原位培养。培养30 d后取出培养管。同时将下一批管按上述方法布置于前一次培养点附近。每个取样点的土样均匀混合后过2 mm筛,装入自封袋中,低温带回实验室做室内分析。实验从2007年5月18日开始,至2007年10月15日结束,每个培养时期约为30 d,共进行5个时段的培养。
2.2 测定项目与方法
新鲜土样的NH4—N质量分数用靛酚蓝比色法测定,NO3—N质量分数用紫外分光光度法测定;全氮质量分数用浓H2SO4—2HClO4消化,开氏定氮测定分析;pH值用1 mol·L-1的KCL浸提,m(土) ∶m(水)=1∶2.5,复合电极法;土壤有机质采用重铬酸钾—外加热法测定。
2.3 数据分析
本实验所有数据采用SAS9.0软件进行分析,数据统计采用单因子方差分析,进行不同培养时间净氮矿化量和净氮矿化速率的比较,用Duncan多重比较及双因素方差分析对研究对象之间的差异进一步检验。
按以下公式计算有关参数:
矿化速率=[(培养后NO3—N质量分数+NH4—N质量分数)-(土壤初始NO3—N质量分数+NH4—N质量分数)]/培养时间;
硝化速率=[(培养后NO3—N质量分数)-(土壤初始NO3—N质量分数)]/培养时间;
铵化速率=[(培养后NH4—N质量分数)-(土壤初始NH4—N质量分数)]/培养时间。
3 结果与分析
3.1 土壤NH4—N和NO3—N质量分数
整个生长季,4种林型土壤的NH4—N质量分数的月动态变化总体上呈单峰式的季节变化特征,最低值均出现在5月份,但最高值出现的月份却因林型而异(见表3)。草类林土壤的NH4—N质量分数最高值出现在6月份,而赤杨林和柴桦林土壤的NH4—N质量分数在7月份达到最高值,杜香林土壤的NH4—N质量分数则在8月份达到最高;土壤NH4—N 质量分数的变化范围是 4.14~56.85 μg·g-1。方差分析结果表明:就不同林型而言,草类林、赤杨林和杜香林三者之间土壤NH4—N质量分数差异显著(P<0.0001);赤杨林和柴桦林土壤NH4—N质量分数差异显著(P<0.0001)。5、6、7 月份彼此之间土壤NH4—N质量分数差异显著(P<0.0001);7、9月份差异显著(P<0.0001)。同时,双因素方差分析结果表明:不同月份和林型的土壤NH4—N质量分数差异显著(P<0.0001),月份和林型之间有显著的交互作用。
4种林型土壤的NO3—N质量分数季节变化格局与NH4—N季节格局相似,同样表现为单峰曲线,但质量分数在林型间及月份间存在差异(表3)。草类林的土壤NO3—N质量分数在6月份达到最高,为(14.64±0.72)μg·g-1;赤杨林和柴桦林的土壤NO3—N质量分数的最高值出现在7月份,分别为(47.52±3.59)、(26.92±5.60)μg·g-1;而杜香林的最高值则出现在 9 月份,为(18.03±3.67)μg·g-1。方差分析结果表明:4种林型的土壤NO3—N质量分数差异不显著;整个生长季中,7月份与其他4个月份之间土壤NO3—N质量分数差异显著(P<0.0001)。双因素方差分析结果表明:不同月份和林型的土壤NO3—N质量分数差异显著(P<0.01),月份和林型之间有显著的交互作用。
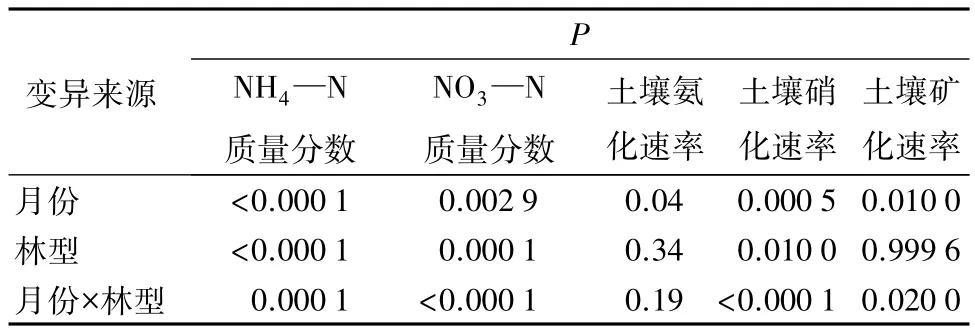
表2 月份、林型及其交互作用对土壤氮矿化的双因素方差分析
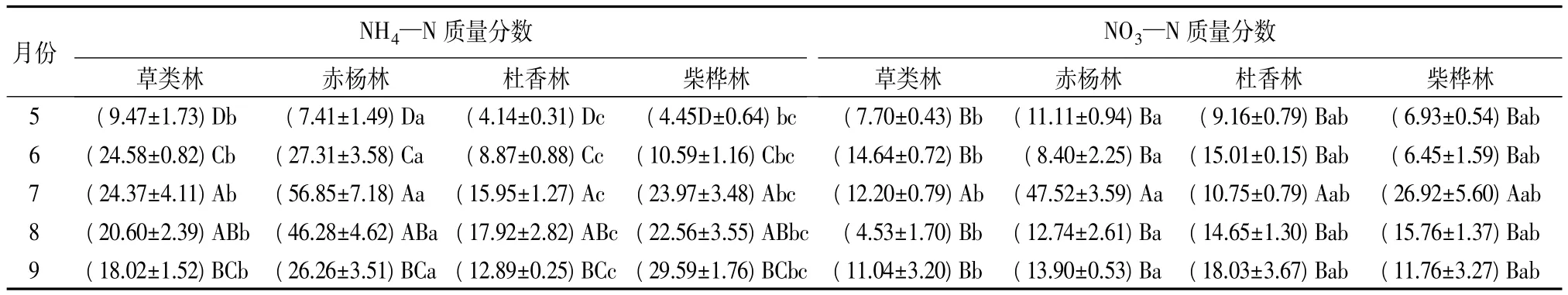
表3 4种林型土壤NH4—N和NO3—N质量分数 μg·g-1
3.2 土壤氮矿化速率
4种林型的氨化速率存在明显的季节变化(见表4),草类林和杜香林7月份最高,9月份最低;赤杨林和柴桦林5月份最低,7月份最高。总体上,4种林型土壤氨化速率的变化范围是 0.28 ~1.59 μg·g-1·d-1,土壤平均净氨化率从大到小排序为:赤杨林(0.92±0.13)μg·g-1·d-1、柴桦林(0.85±0.18)μg·g-1·d-1、杜香林(0.83±0.22)μg·g-1·d-1、草类林(0.70±0.13)μg·g-1·d-1。方差分析表明:4 种林型土壤氨化速率差异不显著;整个生长季中,6、7、8月份彼此之间土壤氨化速率差异显著(P<0.05)。另外,双因素方差分析结果表明:月份和林型之间没有显著的交互作用。
4种林型硝化速率的季节变化和氨化速率相似,7月份硝化速率最高,草类林、赤杨林、杜香林和柴桦林土壤的最大硝化速率分别为(0.75±0.07)、(0.30±0.08)、(0.66±0.10)、(1.18±0.28) μg·g-1·d-1(表4)。总体上,4种林型土壤硝化速率的变化范围是-0.28~1.18 μg·g-1·d-1。由于森林土壤硝化作用的NH4—N来源于氨化作用,因此硝化速率往往低于氨化速率[7]。方差分析表明:就林型而言,草类林和赤杨林土壤硝化速率的差异显著(P<0.01,);整个生长季中,6、7、9 月份彼此之间差异显著(P<0.0005);月份和林型之间有显著的交互作用(P<0.0001)。
4种林型的氮矿化月动态变化趋势与氨化速率变化趋势相似,这种相似在赤杨林和柴桦林土壤中表现尤为明显(见表4),草类林和杜香林7月份最高,9月份最低;赤杨林和柴桦林5月份最低,7月份最高。总体上,4种林型土壤矿化速率变化范围是0.31~2.74 μg·g-1·d-1;土壤平均净矿化率从大到小排序为:赤杨林(1.04±0.15)μg·g-1·d-1、杜香林(1.03±0.31)μg·g-1·d-1、柴桦林(1.03±0.40)μg·g-1·d-1、草类林(1.02±0.22)μg·g-1·d-1。方差分析表明:不同林型间土壤氮矿化速率差异不显著;整个生长季中,6、7、8 月份彼此间差异显著(P<0.01);林型和月份间有显著交互作用(P<0.02)(表 2、表 4)。
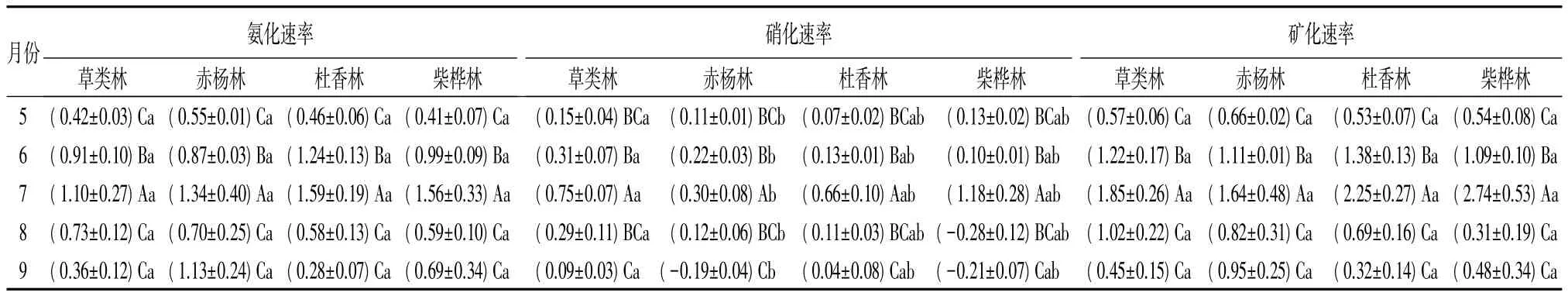
表4 4种林型土壤氨化速率、硝化速率、矿化速率 μg·g-1·d-1
4 讨论
4.1 NH4—N是森林土壤无机氮的主体形态
经过生长季的培养,在4种林型土壤中,NH4—N质量分数在无机氮中所占比例变化范围,草类林为55%~82%,赤杨林为40%~78%,杜香林为31%~60%,柴桦林为39%~72%。草类林、赤杨林、杜香林和柴桦林的土壤NO3—N在无机氮中的比例变化范围分别为18%~45%、22%~60%、40%~69%和28%~61%。NH4—N是土壤无机氮的主要存在形式。这与众多研究结果相一致,例如莫江明等[12]研究得出鼎湖山马尾松阔叶混交林土壤的NH4—N占无机氮的90%;李贵才等[13]得出云南哀牢山木果柯(Lithocarpus xylocarpus)林的NH4—N占95%以上;沙丽清等[14]得出在云南西双版纳不同热带森林中NH4—N 占 60.14% ~80.74%;孟盈等[2]发现云南西双版纳森林土壤的NH4—N质量分数也较高,龙山林、季节雨林和橡胶林年均铵态氮分别约占67%、78%和82%。
4.2 不同森林类型对土壤氮矿化速率的影响
土壤矿化作用是指有机氮在微生物作用下转化为无机态氮(主要是铵态氮和硝态氮)的过程,为正值。不同植被类型的氮素矿化速率有很大差异[15]。李贵才等[13]对哀牢山森林生态系统的研究认为,不同生态系统的植被类型、微生物种类差异导致了矿化量和矿化速率的变化。苏波等[16]在东灵山油松(Pinus tabulaeformis)纯林和油松—辽东栎(Quercuswutaishanica)针阔混交林中进行的土壤氮素矿化/硝化作用研究发现,物种构成及在其影响下所产生的林下微生境和人为干扰活动是造成两种生态系统的土壤中氮素有效性和维持植物可利用氮素的能力有显著差异的主要原因。Knoepp[17]对阿巴拉契亚南部不同植被和不同海拔高度的氮矿化速率长达6a的研究发现,植被类型是氮矿化速率的主要控制因素。生态系统构成的差异是导致4种林型间矿化差异的主要原因。本研究表明,4种林型土壤净氮矿化量和矿化速率均为正值,说明这4种林型土壤的有机氮正向无机氮转化,表现为无机氮的净积累。本研究中赤杨是固氮植物,自身会固定一部分氮,加上有机质的分解,会促进氮的矿化进程。大量研究表明,外加氮源的输入将会在一定程度上增加土壤氮素矿化速率,因而赤杨林的净矿化速率稍高于其它3种林型。
4.3 土壤净氮矿化速率与环境因子的关系
赤杨林矿化速率最高,但是柴桦林和杜香林的矿化速率和赤杨林彼此之间相差并不大,这就说明外加氮源并不是氮矿化速率的唯一决定因素。土壤氮矿化的影响因素很多,原因很复杂,而土壤的氮矿化速率受土壤肥力、基质、水热条件和微生物等因素综合影响[17]。有研究表明,不同气候条件下氮循环也有很大差异,说明氮矿化受气象因素的影响也很大[17]。对于本研究来说,4种兴安落叶松林的样地均同时进行土壤氮矿化的培养,在同一气候条件下进行试验,可以不考虑影响因素;但是氮矿化速率随坡度、湿度的不同而变化,同时由于植被的阴蔽度不同而造成的温度变幅和极值差异对氮矿化速率也有一定影响[18]。陈印平等[19]对中亚热带栲木荷(Castanopsisplatyacantha-Schina sinensis)土壤氮矿化研究发现,土壤的碳氮质量比和全碳质量分数高,则硝化速率和矿化速率较高。本研究中,杜香林的土壤氮矿化速率虽不是最高的,但与赤杨林相差不大,且差异不显著,这可能是杜香林土壤有机质质量分数高,碳氮质量比高,有利于微生物的活性,导致硝化速率和矿化速率较高。这与Vitousek和Sanford[20]发现肥力高的热带土壤的氮矿化速率高于肥力低的热带土壤结果相似,说明不仅是热带,在寒温带肥力高的土壤其氮矿化速率也高于肥力低的土壤。一般的研究表明高温、低湿环境有利于提高土壤的氮素矿化速率。周长平等[21]对长白山两种主要林型研究发现,温度对土壤微生物活性的影响非常显著,两种类型森林土壤的氮矿化速率都与温度呈正相关。本研究中柴桦林处于湿地,水分较饱和,土壤湿度的变化幅度小,使土壤透气性差,导致某些厌氧微生物和反硝化细菌生长活跃,因而其土壤矿化速率较低,但也由于其下有深厚的泥炭层,有机质质量分数较高,因而柴桦林土壤氮矿化速率高于草类林。
森林生态系统的氮矿化过程比较复杂,影响因素也很多,不同植物的遮阴状况、生物固氮能力、凋落物产量和质量以及土壤动物、微生物种类和活性的不同都会对土壤的氮矿化速率造成影响。本研究中兴安落叶松林下4种林型的氮矿化速率相差并不是很大,很难说明究竟是哪种因素占主导地位,所以要较全面地认识大兴安岭森林生态系统的氮循环特征及其影响因素,还需进行长期的野外定位研究以及室内定量研究。
5 结论
兴安落叶松林4种林型中,NH4—N在无机氮中所占比例约为31%~82%,NO3—N在无机氮中所占比例约为18%~69%,NH4—N是土壤有效氮的主要存在形式。不同月份和林型的土壤NH4—N和NO3—N质量分数差异显著(P<0.01),月份和林型之间有显著的交互作用。
兴安落叶松林土壤氨化速率与硝化速率存在明显的季节特征。7月份的氨化作用和硝化作用活动最强烈,该时期草类林、赤杨林、杜香林和柴桦林4种林型土壤的最大净氨化率分别为(1.10±0.27)、(1.34±0.40)、(1.59±0.19)、(1.56±0.33)μg·g-1·d-1;最大净硝化率分别为(0.75±0.07)、(0.30±0.08)、(0.66±0.10)、(1.18±0.28)μg·g-1·d-1。林型间差异不显著,氨化速率月份和林型之间没有显著的交互作用,硝化速率月份和林型之间有显著的交互作用(P<0.0001)。
兴安落叶松林4种林型的矿化作用存在明显的季节变化。土壤平均净矿化率从大到小依次为:赤杨林(1.04±0.15)μg·g-1·d-1、杜香林(1.03±0.31)μg·g-1·d-1、柴桦林(1.03±0.40)μg·g-1·d-1、草类林(1.02±0.22)μg·g-1·d-1。不同林型之间土壤氮矿化速率差异不显著,林型和月份之间有显著的交互作用(P<0.02)。
[1]Aber J D,Melillo J M.Terrestrial ecosystem[M].2nd.San Diego:Harcourt Academic Press,2001.
[2]孟盈薛,敬意,沙丽清,等.西双版纳不同热带森林下土壤氨态氮和硝态氮动态研究[J].植物生态学报,2001,25(1):99-104.
[3]Schimel J P,Bennett J.Nitrogen mineralization:challenges off a changing paradigm[J].Ecology,2004,85(3):591-602.
[4]Ineson P,Taylor K,Harrison A F,et al.Effects of climate change on nitrogen dynamics in upland soils(1):A transplant approach[J].Global Change Biology,1998,4(2):143-152.
[5]Finzi A C,Delucia E H,Hamilton J G,et al.The nitrogen budget of a pine forest under free air CO2enrichment[J].Oecologia,2002,132(4):567-578.
[6]McMurtrie R E,Medlyn B E,Dewar R C.Increased understanding of nutrient immobilization in soil organic matter is critical for predicting the carbon sink strength of forest ecosystem over the next 100 years[J].Tree Physiology,2001,21(12/13):831-839.
[7]Swift M J,Andren O,Brussaard L,et al.Global change,soil biodiversity,and nitrogen cycling in terrestrial ecosystem:three case studies[J].Global Change Biology,1998,4(7):729-743.
[8]Bruun S,Stenberg B,Breland T A,et al.Empirical predictions of plant material C and N mineralization patterns from near infrared spectroscopy,stepwise chemical digestion and C/N ratios[J].Soil Biology and Biochemistry,2005,37(12):2283-2296.
[9]Hart S C,Nason G E,Myrold D D,et al.Dynamics of gross nitrogen transformations in an old-growth forest:the carbon connection[J].Ecology,1994,75(4):880-891.
[10]Chen Y,Hogberg P.Gross nitrogen mineralization rates still high 14 years after suspension of N input to a N-saturated forest[J].Soil Biology and Biochemistry,2006,38(7):2001-2003.
[11]Corre M D,Lamersdorf N P.Reversal of nitrogen saturation after long-term deposition reduction:impact on soil nitrogen cycling[J].Ecology,2004,85(11):3090-3104.
[12]莫江明,方运霆,林而达,等.鼎湖山主要森林土壤N2O排放及其对模拟N沉降的响应[J].植物生态学报,2006,30(6):901-910.
[13]李贵才,韩兴国,黄建辉.哀牢山木果柯林及其退化植被下土壤无机氮库的干季动态特征[J].植物生态学报,2001,25(2):210-217.
[14]沙丽清,孟盈,冯志立,等.西双版纳不同热带森林土壤氮矿化和硝化作用研究[J].植物生态学报,2000,24(2):152-156.
[15]Clemensson-Lindell A,Persson H.The effects of nitrogen addition and removal on Norway spruce fine-root vitality and distribution in three catchment areas at Gardsjon[J].Forest Ecology and Management,1995,71(1/2):123-131.
[16]苏波,韩兴国,渠春梅,等.东灵山油松纯林和油松—辽东栎针阔混交林土壤氮素矿化硝化作用研究[J].植物生态学报,2001,25(2):195-203.
[17]Knoepp J D,Swank W T.Using soil temperature and moisture to predict forest soil nitrogen mineralization[J].Biology and Fertility of Soils,2002,36(3):177-182.
[18]Fernandez I J,Simmons J A,Briggs R D.Indices of forest floor nitrogen status along a climate gradient in Maine,USA[J].Forest Ecology and Management,2000,134(1/3):177-187.
[19]陈印平,潘开文,吴宁,等.凋落物质量和分解对中亚热带栲木荷林土壤氮矿化的研究[J].应用与环境生物学报,2005,11(2):146-151.
[20]Vitousek P M,Sanford R L.Nutrient cycling in moist Tropical forests[J].Annual Review of Ecology and Systematics,1986,17:137-167.
[21]周才平,欧阳华.长白山两种主要林型下土壤氮矿化速率与温度的关系[J].生态学报,2001,21(9):1469-1473.
