炎症因子与肝癌干细胞标志在肝癌诱导过程中的相关性研究
2014-08-30宦宏波吴黎雳
吴 林, 宦宏波, 吴黎雳, 别 平,夏 锋
(第三军医大学西南医院 全军肝胆外科研究所, 重庆, 400038)
Kupffer细胞是人体内数量最多的巨噬细胞,约占巨噬细胞总数的80%~90%[1]。它构成了肝脏先天免疫系统的重要组成部分,具有吞噬和杀灭细菌、凋亡细胞、微生物、内毒素、免疫复合物等功能[2-3]; 另一方面, Kupffer细胞在发挥免疫功能的同时释放白细胞介素6(IL-6)、肿瘤坏死因子α(TNF-α)、γ干扰素(IFN-γ)等炎症因子参与反应[3-6], 而先前研究证实炎症反应在肿瘤发生发展过程中起决定性因素[7-9],而且炎症因子进一步参与了肿瘤干细胞的自我更新和上皮细胞间质转化(EMT), 促进肿瘤的发生[10-11]。近年来有研究[12-13]报道Kupffer细胞通过释放炎症因子参与肝癌的发生发展,但关于Kupffer细胞分泌的炎症因子和肝癌干细胞标志在肝癌发生发展中的动态表达及其相关性尚未见报道。本研究通过建立大鼠肝癌诱导模型,采用定量PCR方法检测肝癌发展过程中不同时间点Kupffer细胞分泌的炎症因子及肝癌干细胞标志的动态变化,并对2者相关性进行统计分析。
1 材料与方法
1.1 材料与试剂
SD雄性大鼠(第三军医大学动物中心)、二乙基亚硝胺(DEN, 0.95 g/mL, 美国Sigma公司)、ED2抗体(英国Serotec AbD公司)、磷酸盐缓冲液(PBS,中杉金桥)、免疫组织化学检测试剂盒(中杉金桥)、DAB显色试剂盒(中杉金桥)、Trizol(日本Takara公司)、PrimeScript RT reagent Kit反转录试剂盒(日本Takara公司)、SYBR Green(日本Takara公司)、引物合成(北京华大基因公司)。
1.2 仪器与设备
Olympus显微镜(日本Olympus公司)、ND1000微量分光光度计(美国Nano Drop公司)、CFX96实时定量PCR仪(美国Bio-Rad公司)。
1.3 试验方法
1.3.1 肝癌诱导大鼠模型建立: SD雄性大鼠35只(180 g~200 g, 第三军医大学动物中心),随机分为正常对照组(5只),诱导模型组按诱癌时间点分为: 4周、8周、12周、16周、20周、24周(每组5只)。24周组每只大鼠肝脏组织又分为癌和癌旁2个亚组: 24-周-NT(癌旁)、24-周-T(癌)。诱导模型组大鼠按每天10 mL/100 mg body weight饮水中给予DEN溶液(0.01% v/v), 每周仅给予6天DEN溶液,剩余1 d给予普通饮水。正常对照组给予普通饮水。在诱癌过程中,每隔4周取材诱癌模型组5只大鼠的肝脏, -80 ℃冷藏备用,直至诱癌第24周肝癌形成。
1.3.2 免疫组织化学检测诱癌过程中Kupffer细胞ED2表达:大鼠肝脏组织常规甲醛固定、石蜡包埋、连续切片、烤片后,按免疫组化染色试剂盒说明书进行。脱蜡后, 3% H2O2封闭,抗原修复,山羊血清封闭,一抗ED2(1∶200)4 ℃冰箱过夜,滴加酶标抗体,通用二抗结合, DAB显色。
各试验组ED2阳性细胞分数计算方法:每只大鼠计数平均5个高倍镜视野下每100个细胞中ED2阳性细胞表达分数,取均值和标准差后计算每一组大鼠ED2阳性细胞分数。
1.3.3 实时定量PCR检测炎症因子和肝癌干细胞标志表达:所有操作均严格按照试剂说明书进行。常规Trizol提取各组大鼠肝脏组织RNA, 测定RNA浓度后,反转录试剂盒将mRNA反转为cDNA,加入特定引物(引物序列见表1),用SYBR Green试剂盒实时定量PCR检测Kupffer细胞释放的相关炎症因子IL-1α、IL-1β、IL-6、IL-8、IL-10、TNF-α、IFN-γ、单核细胞趋化蛋白-1(MCP-1)、转化生长因子β(TGF-β)和肝癌干细胞指标CD90、CD133、Sirt1、AFP各基因表达, GAPDH作为内参。
1.3 统计学分析
本试验所有数据均采用SPSS 17.0进行统计分析。诱癌过程中,各试验组Kupffer细胞ED2阳性分数、Kupffer细胞相关炎症因子表达和肝癌干细胞标志表达均采用单因素方差分析(ANOVA)和Tukey多重比较。炎症因子与肝癌干细胞指标之间的相关性采用Pearson相关性检验进行分析。P<0.05为差异有统计学意义。
2 结 果
2.1 DEN诱导大鼠肝癌模型的建立
通过对诱导模型组大鼠饮水中加入DEN溶液,大鼠肝癌演变过程表现为经典的炎症反应期(1~12周)、肝增生纤维化期(12~16周)、肝硬化形成期(16~20周)、肝癌形成期(20~24周)(见图1, A-N)。到诱癌24周时, 24周组大鼠均发现肝癌形成。

表1 引物合成序列
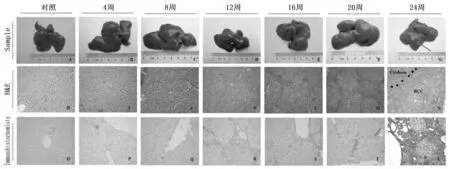
图1 大鼠肝癌诱导模型建立及Kupffer细胞在诱癌过程中的动态变化100倍
2.2 大鼠肝癌诱导过程中Kupffer细胞ED2表达
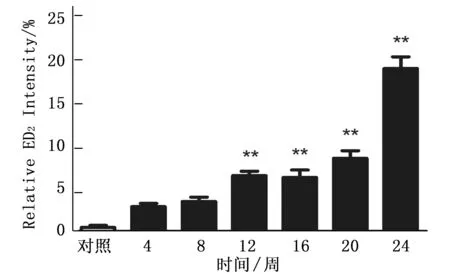
通过免疫组化染色检测Kupffer细胞特异性抗体ED2,观察大鼠肝癌诱导过程中ED2动态表达,免疫组化结果提示ED2在肝癌诱导过程中表达逐渐增多(图1, O-U), 并通过统计分析计算出各组ED2阳性细胞分数,结果表明ED2在肝癌诱导过程中呈逐渐升高的趋势(P<0.001)。而且从诱癌第12周开始,12周、16周、20周和24周的ED2的表达与正常对照组有明显差异性(P<0.01)。该结果表明Kupffer细胞数量在诱癌过程中呈逐渐增多的趋势。(图2)。
2.3 大鼠肝癌诱导过程中Kupffer细胞相关炎症因子的动态表达
注:**P<0.05
通过定量PCR方法检测鼠肝癌诱导过程中Kupffer细胞相关炎症因子的动态表达,结果表明IL-6(P<0.001)、MCP-1(P<0.001)、TGF-β(P=0.003)和TNF-α(P=0.047)在诱癌过程中明显升高(图3 A,B,C,D), 而且与正常肝脏组织相比较,在肝癌组织中IL-6(P<0.001)、MCP-1(P<0.001)和TGF-β(P<0.001)表达更高,而其它炎症因子在诱癌过程中无明显变化(图3, E-I,P>0.05)。
2.4 大鼠肝癌诱导过程中肝癌干细胞标志的动态表达

注:**P<0.05
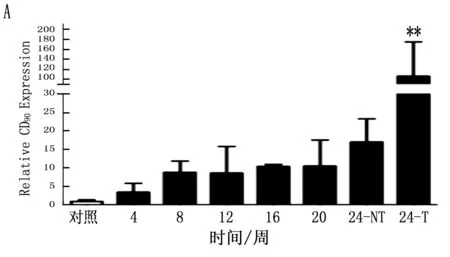
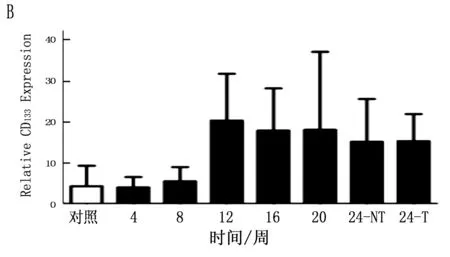
通过定量PCR方法检测鼠肝癌诱导过程中肝癌干细胞标志的动态表达,结果表明CD90在诱癌过程中呈逐渐升高的趋势(P<0.001),特别是与正常肝脏组织比较,肝癌组织中CD90升高更明显(图4 A;P<0.001)。CD133虽然呈一定升高趋势,但ANOVA分析其结果无明显变化(P=0.125), 同时其它肝癌干细胞标志物在诱癌过程中无明显变化(图4 B,C,D;P>0.05)
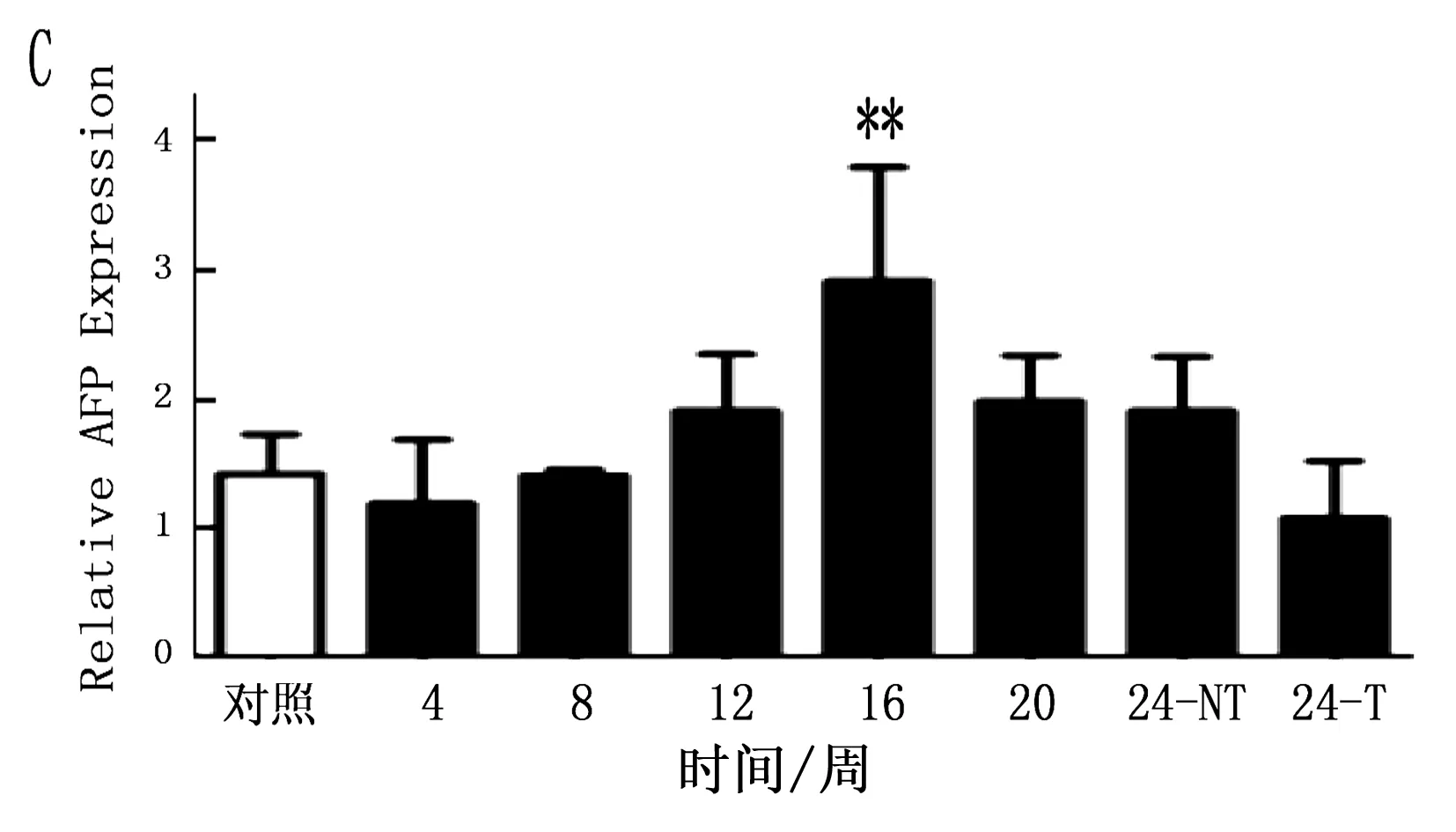
注:**P<0.05
2.5 Kupffer相关炎症因子与肝癌干细胞标志的相关性
通过以上结果统计分析筛选出在诱癌过程中差异性表达的Kupffer相关炎症因子: IL-6、MCP-1、TGF-β和TNF-α, 以及差异性表达肝癌干细胞标志CD90。将肝癌诱导过程中CD90与IL-6、MCP-1、TGF-β和TNF-α的动态表达进行Pearson相关性分析,结果表明IL-6、MCP-1、TGF-β和CD90的表达呈明显正相关性(图5 A,B,C;P<0.001), 而TNF-α与CD90表达无明显相关性(图5 D;P=0.288)。
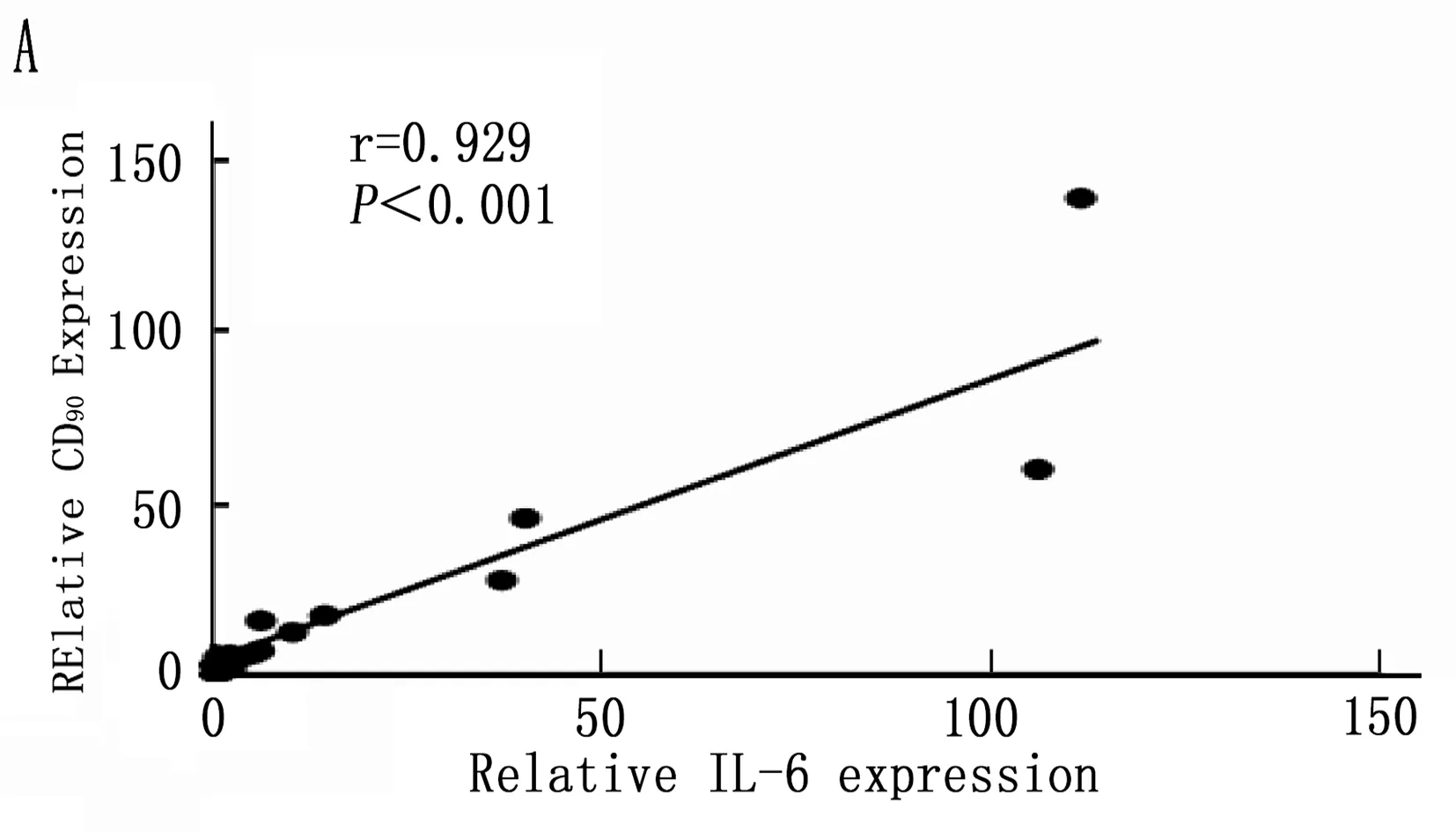
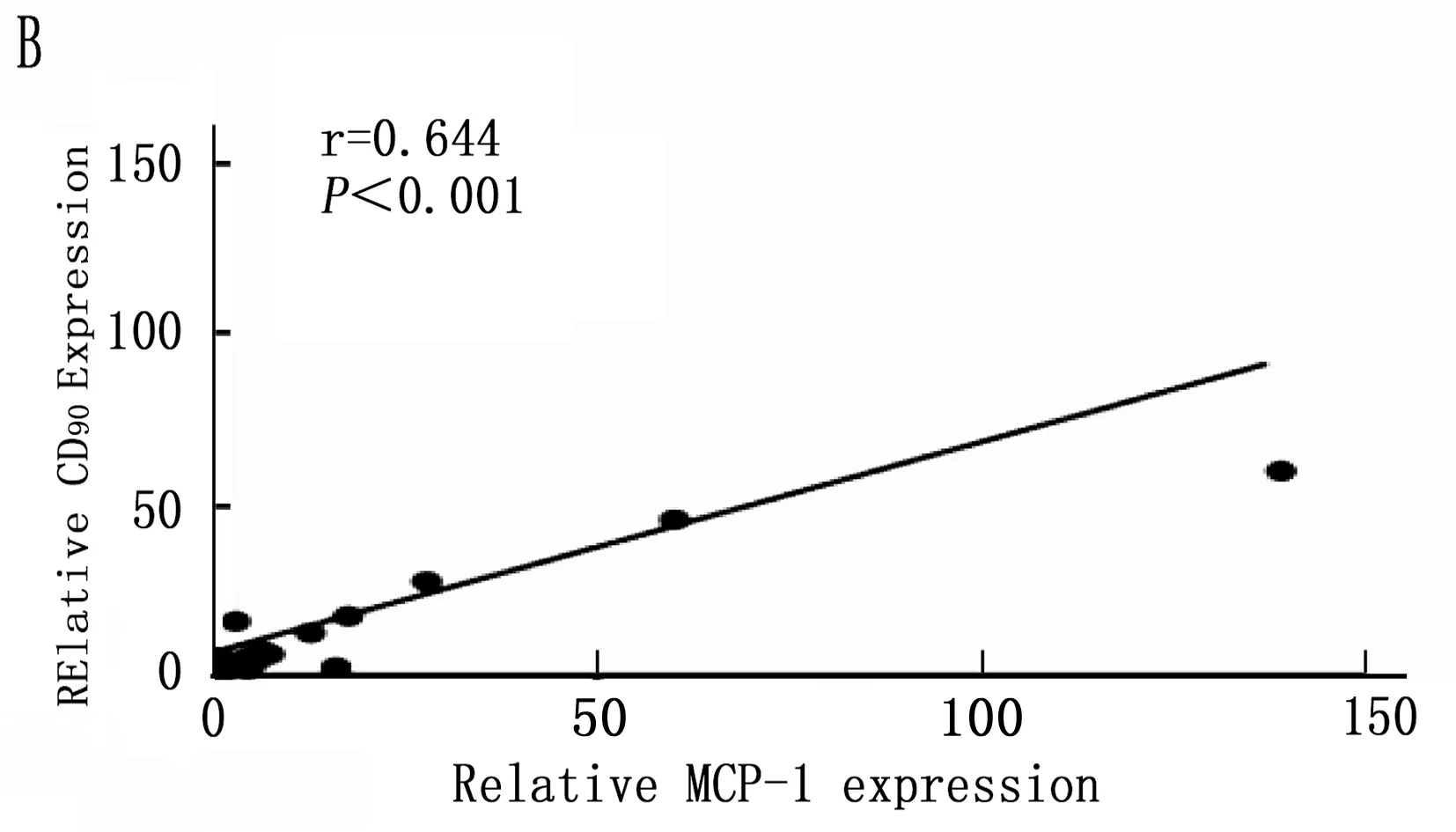
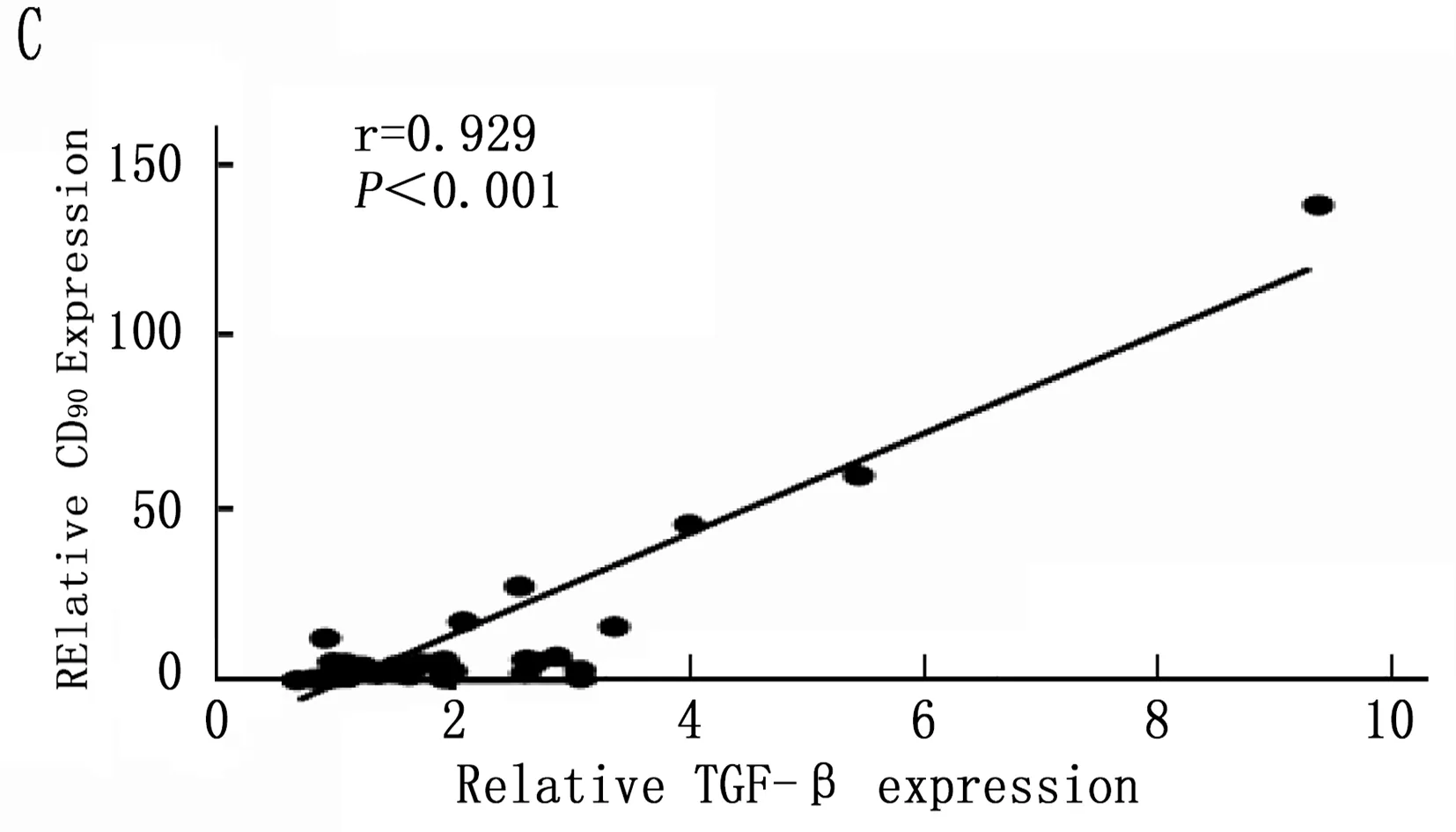
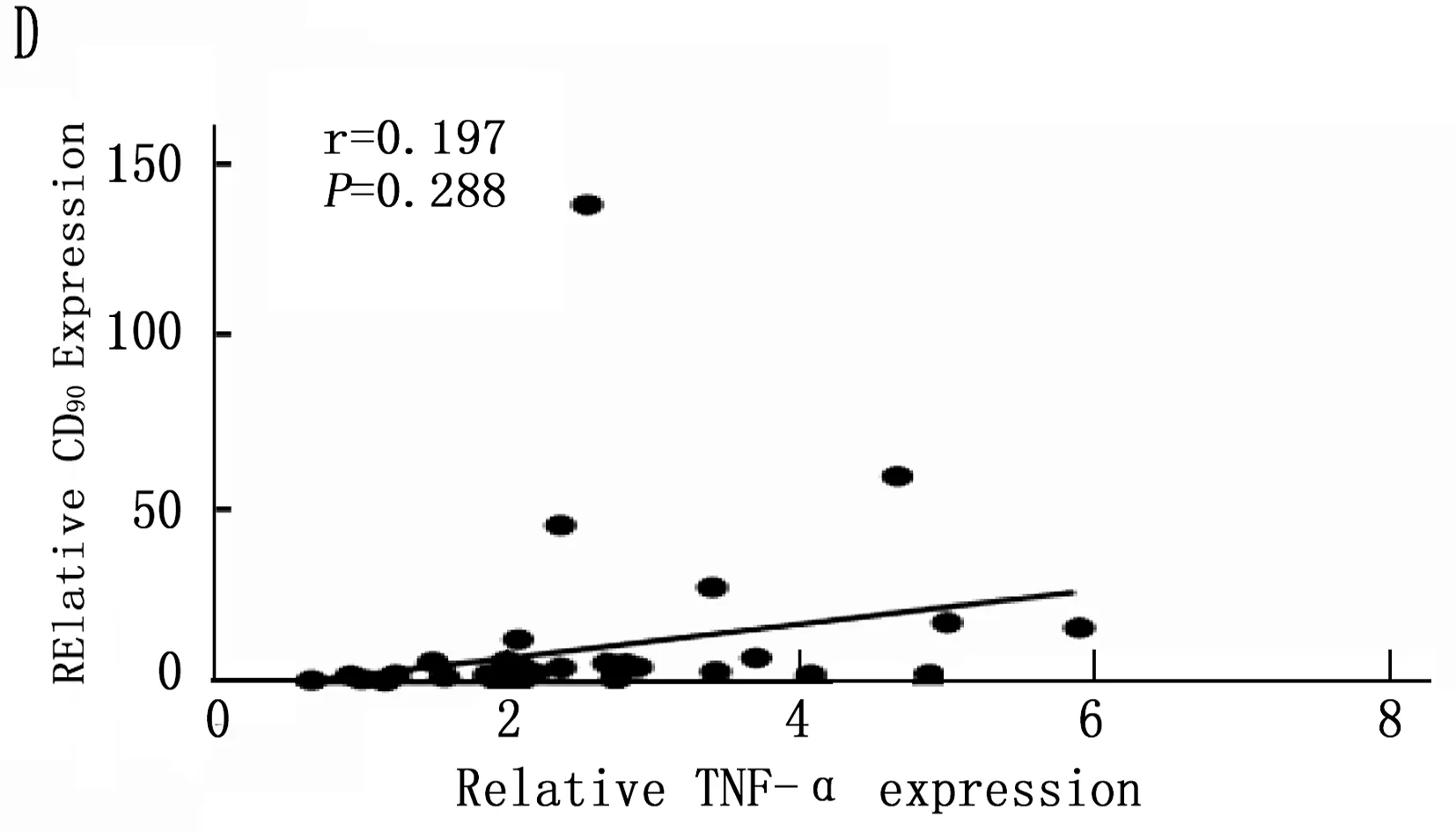
图5 肝癌诱导过程中CD90与Kupffer细胞相关炎症因子的相关性分析
3 讨 论
本研究首先通过DEN诱导成功建立经典大鼠肝硬化肝癌模型,在诱癌24周时发现肝脏肿瘤,然后对各组大鼠采用免疫组织化学方法检测了Kupffer细胞的特异性抗体ED2[14-15]在肝癌诱导过程中的动态变化,结果表明ED2阳性细胞在诱癌过程中逐渐升高(图2;P<0.001)。采用定量PCR方法检测了Kupffer细胞释放的炎症因子在诱癌过程中的表达变化,结果筛查出IL-6(P<0.001)、MCP-1(P<0.001)、TGF-β(P=0.003)和TNF-α(P=0.047)在诱癌过程中变化明显(图3 A,B,C,D); 同时肝癌干细胞标志物筛查出CD90指标在诱癌过程中呈明显升高趋势(图4 A;P<0.001)。通过Pearson检验分析IL-6、MCP-1、TGF-β、TNF-α与CD90的相关性,提示IL-6、MCP-1、TGF-β与CD90的表达明显相关(图5 A,B,C;P<0.001)。从而证明Kupffer细胞产生的炎症因子与肝癌的发生发展密切相关。
许多研究证实炎症因子与肿瘤发生之间存在密切关系。Naugler WE等[12]通过研究DEN对不同性别大鼠肝癌形成的影响,证明雌性大鼠通过雌激素调控Kupffer细胞上Toll样受体衔接蛋白MyD88,抑制坏死肝细胞释放IL-6以及清除循环血液中的IL-6, 从而达到抑制化学性诱癌的发生。Wu K等[16]在一部分大鼠及人体肝癌组织中发现肝脏前体细胞共表达肿瘤初始细胞标志(T-IC), 同时TGF-β的表达与T-IC的表达呈正相关性,而在缺乏TGF-β的情况下却不能启动肝癌的发生,从而证明了TGF-β在肝癌发生发展中的重要作用。Wang WW等[17]通过蛋白抗体芯片检测了58例肝癌切除患者及11例乙肝病毒携带者的血清蛋白,结果发现113种血清标志物被调控,其中MCP-1的变化最为明显,而且MCP-1有希望作为AFP的一种补偿标志物,通过ELISA常规定量检测患者血清MCP-1浓度,从而帮助临床预测和诊断肝癌。Yang ZF等[18]对肝癌患者循环血液及肝癌组织进行研究发现, CD45-、CD90+细胞可以在90%的肝癌患者循环血液及肝癌组织中被检测,证明CD90可以作为检测和诊断肝癌的一个重要标志物。
部分研究也明确了炎症因子与肿瘤干细胞之间的相关性。Jin X等[10]证明在脑肿瘤发生过程中,干扰素调节因子7(IRF-7)可以通过IL-6和Notch信号通路诱导神经胶质瘤细胞的产生以及血管生成。此外, Li Y[11]等通过研究结肠癌细胞,发现IL-1β可以通过激活肿瘤干细胞的自我更新及EMT促进结肠癌的生长与侵袭。Korkaya H等[19]对曲妥珠单抗的耐药乳腺癌细胞系研究发现, IL-6炎症反馈通路导致了肿瘤干细胞的扩增,长时间通过曲妥珠单抗处理的细胞系筛选出大量的肿瘤干细胞,这些肿瘤干细胞表现出EMT现象及其分泌的IL-6相比普通细胞系表达高出近百倍,同时通过阻断IL-6受体可以明显降低肿瘤的生长和转移。以上试验均表明炎症因子对肿瘤干细胞的重要调控作用,而Kupffer细胞释放的相关炎症因子与肝癌干细胞标志CD90的相关性尚未见报道。
本研究尚存在一些不足之处: ① Kupffer细胞相关炎症因子促进肝癌干细胞标志表达的具体机制还需进一步功能学试验明确; ②本试验结果筛查出CD90与Kupffer细胞释放的炎症因子明显相关,但其它肝癌干细胞标志物与炎症因子的相关性还需进一步探讨。因此,作者设想下一步是否可以通过抑制Kupffer细胞释放的炎症因子,或将Kupffer细胞清除达到降低甚至抑制肝癌的发生。
综上所述,本研究结果表明Kupffer细胞释放的炎症因子IL-6、MCP-1、TGF-β的表达升高与CD90上调呈明显正相关,进一步明确了Kupffer细胞促进肝癌发生的重要作用。因此, Kupffer细胞可以作为潜在的肝癌治疗靶点为肝癌的临床诊治开辟新的领域。
[1]Liaskou E, Wilson D V, Oo YH. Innate immune cells in liver inflammation[J]. Mediators Inflamm, 2012, 2012: 949157.
[2]Kolios G, Valatas V, Kouroumalis E. Role of kupffer cells in the pathogenesis of liver disease[J]. World J Gastroenterol, 2006, 12(46): 7413.
[3]Laskin D L. Macrophages and Inflammatory Mediators in Chemical Toxicity: A Battle of Forces[J]. Chem Res Toxicol, 2009, 22(8): 1376.
[4]Helk E, Bernin H, Ernst T, et al. TNFα-mediated liver destruction by Kupffer cells and Ly6Chi monocytes during Entamoeba histolytica infection[J]. PLoS Pathog, 2013, 9(1): e1003096.
[5]Lucey, M. R., P. Mathurin, et al. Alcoholic hepatitis[J]. N Engl J Med, 2009, 360(26): 2758.
[6]Xu L, Yin W, Sun R, et al. Kupffer cell-derived IL-10 plays a key role in maintaining humoral immune tolerance in hepatitis B virus-persistent mice[J]. Hepatology, 2013, doi: 10.1002/hep.26668.[Epub ahead of print]
[7]Grivennikov S I, Greten F R, Karin M. Immunity, Inflammation, and Cancer[J]. Cell, 2010, 140(6): 883.
[8]Park E J, Lee J H, Yu G Y, et al. Dietary and genetic obesity promote liver inflammation and tumorigenesis by enhancing IL-6 and TNF expression[J]. Cell, 2010, 140(2): 197.
[9]Ben-Neriah Y, Karin M. Inflammation meets cancer, with NF-κB as the matchmaker[J]. Nat Immunol, 2011, 12(8): 715.
[10]Jin X, Kim S H, Jeon H M, et al. Interferon regulatory factor 7 regulates glioma stem cells via interleukin-6 and Notch signalling[J]. Brain, 2012, 135(Pt 4): 1055.
[11]Li Y, Wang L, Pappan L, et al. IL-1β promotes stemness and invasiveness of colon cancer cells through Zeb1 activation[J]. Mol Cancer. 2012, 11: 87.
[12]Naugler W E, Sakurai T, Kim S, et al. Gender Disparity in Liver Cancer Due to Sex Differences in MyD88-Dependent IL-6 Production[J]. Science, 2007, 317(5834): 121.
[13]Maeda S, Kamata H, Luo J L, et al. IKKbeta couples hepatocyte death to cytokine-driven compensatory proliferation that promotes chemical hepatocarcinogenesis[J]. Cell, 2005, 121(7): 977.
[14]Tapia G, Santibá?ez C, Farías J, et al. Kupffer-cell activity is essential for thyroid hormone rat liver preconditioning[J]. Mol Cell Endocrinol, 2010, 323(2): 292.
[15]Golbar H M, Izawa T, Murai F, et al. Immunohistochemical analyses of the kinetics and distribution of macrophages, hepatic stellate cells and bile duct epithelia in the developing rat liver[J]. Exp Toxicol Pathol, 2012, 64(1/2): 1.
[16]Wu K, Ding J, Chen C, et al. Hepatic Transforming Growth Factor Beta Gives Rise to Tumor-Initiating Cells and Promotes Liver Cancer Development[J]. Hepatology, 2012, 56(6): 2255.
[17]Wang W W, Ang S F, Kumar R, et al. Identification of serum monocyte chemoattractant protein-1 and prolactin as potential tumor markers in hepatocellular carcinoma[J]. PLOS ONE, 2013, 8(7): e68904.
[18]Yang Z F, Ngai P, Ho D W, et al. Identification of Local and Circulating Cancer Stem Cells in Human Liver Cancer[J]. Hepatology, 2008, 47(3): 919.
[19]Korkaya H, Kim G I, Davis A, et al. Activation of an IL6 inflammatory loop mediates trastuzumab resistance in HER2+ breast cancer by expanding the cancer stem cell population[J]. Mol Cell. 2012, 47(4): 570.
