气候变化对中国东北地区日本松干蚧传播扩散的影响
2014-08-10袁福香胡艳全张玉书
袁福香, 刘 实, 胡艳全, 张玉书, 戴 勇, 曹 蕊
(1. 吉林省气象科学研究所, 长春 130062; 2. 中国气象局沈阳大气环境研究所,沈阳 110016;3. 吉林省梅河口市森林病虫防疫监测站,梅河口 135000)
气候变化对中国东北地区日本松干蚧传播扩散的影响
袁福香1,*, 刘 实1, 胡艳全1, 张玉书2, 戴 勇3, 曹 蕊3
(1. 吉林省气象科学研究所, 长春 130062; 2. 中国气象局沈阳大气环境研究所,沈阳 110016;3. 吉林省梅河口市森林病虫防疫监测站,梅河口 135000)
基于气候资料和日本松干蚧传播资料,根据传播扩散范围及入侵地的气候特征,分析了日本松干蚧主要影响因子的年代变化对日本松干蚧在东北地区扩散的影响。结果表明:东北地区最冷月各旬及月平均最低气温总的呈升高趋势(r=0.86,P<0.05),冬季极端最低气温也有缓慢上升趋势(r=0.93,P<0.01),其年代间的冷暖变化与日本松干蚧在东北地区扩散有明显的相关性。1月平均最低气温和冬季极端最低气温明显升高的20世纪70年代和90年代,日本松干蚧快速扩散、危害地虫口密度大、危害程度重。日本松干蚧大范围扩散和爆发都发生在1月份最低气温较高的年份。1月最低气温和冬季极端最低气温升高是日本松干蚧在东北地区传播扩散的重要因素。复苏后的降水量、卵孵化期的空气相对湿度和夏季最高气温的年代变化对日本松干蚧扩散的影响不显著。
日本松干蚧; 传播扩散; 气候变化;东北地区
日本松干蚧是危害松树的重要害虫,该虫1942年在辽宁大连旅顺口老铁山首次被发现[1],在东北地区逐步向北扩散,Mark.S.McClure于1982年通过对中国的日本松干蚧调查,认为中国冬季的低温能抑制该蚧虫向北蔓延,但自20世纪90年代以来,日本松干蚧的扩散蔓延速度加快,1994年入侵到吉林境内[2],到2012年,吉林省的通化市、辽源市、四平市、长春市、吉林市都有该虫危害。30a的时间日本松干蚧又向北扩散了3个纬度的距离,并且没有止步的迹象[3- 6]。
日本松干蚧传播蔓延主要靠风、雨水以及人为活动,该虫近距离自然传播主要传播虫态是初孵若虫,其次是雄成虫、卵囊,扩散最盛期是卵孵化高峰期。樊尚仁[7]等对日本松干蚧传播的途径进行了初步研究,低虫口密度下在林缘外水平扩散距离最远达400m,卵囊(卵粒)和初孵若虫在林间可在半径为1—1.5m范围内的植株间传播扩散。
以往研究表明[8- 17],制约日本松干蚧生存及生长发育的气象因子主要有:冬季极端最低气温,越冬复苏后的降水量、卵孵化期的空气相对湿度和夏季的最高气温。东北地区由南向北,冬季极端最低气温逐渐降低。极端最低气温对日本松干蚧越冬死亡率影响极大,尤其是出现创极值的最低气温。连续8d极端最低气温在-30℃以下,最低为-37.7℃,裸露树干上的日本松干蚧全被冻死[12]。所以冬季极端低温是阻止日本松干蚧向北扩散的重要气象因子。相对湿度在70%—80%时,卵的孵化率高达95%以上,相对湿度小于36%时,大部分卵不能孵化。日本松干蚧的初孵若虫在5℃时能正常活动,1龄若虫在1.5—5.6℃的条件下仍可生长[6- 7]。进入4月份,温度陆续稳定通过0℃,日本松干蚧的寄主松树逐渐恢复生长发育,在松树上越冬的日本松干蚧若虫也随着温度的回升开始复苏,此时处于乍暖还寒时,出现降水(多为雪)会造成气温明显下降,甚至达到0℃或0℃以下。使已经复苏的日本松干蚧的体表温度迅速减低,从而将日本松干蚧冻死。因此4月份降水越多对复苏后日本松干蚧的生存越不利。当夏季温度大于30℃,随着温度的升高,松干蚧寄生若虫死亡率越高。通过2012年对越夏代固定若虫室外采集虫源的温控试验,30℃恒温条件下,72h死亡率为9.67%;35℃恒温条件下,24h死亡率88.89%,48h为96.88%,72h全部死亡;40℃条件下,24h死亡率高达95.65%,48h全部死亡。
自20世纪70年代以来,相关森林病虫防治部门一直对日本松干蚧进行严密防控,但日本松干蚧在东北地区的扩散和蔓延一直没有停止,首先人类活动对该虫的传播起了重要作用[18],该虫个体小、监测难度大是另一影响因子,被传入地气象条件适宜其生存和传播更是日本松干蚧快速蔓延的重要因素。国外专家关于气候变化对一些昆虫等物种传播扩散的影响做过较为深入的研究[19- 21]。在中国东北地区,制约日本松干蚧生长发育的气象因子发生了怎样的变化,这种变化对日本松干蚧的扩散传播发挥了怎样的作用,至今未见相关研究报道。为了探究气候变化对日本松干蚧在东北地区扩散的影响,本文从影响日本松干蚧生长发育的生理气象指标入手,结合传播路径分析气象条件变化及其在传播中的作用,以期对日本松干蚧监测及防控提供参考。
1 资料及处理方法
气候资料来自辽宁省气候中心和吉林省气候中心,包括冬季日最低气温、夏季日最高气温、4月份日降水量、5—6月份和8—9月份的日空气相对湿度。范围为东北地区日本松干蚧危害区和吉林省境内可能危害和扩散、资料序列较长的区域,具体包括:大连市、丹东市、本溪市、鞍山市、沈阳市、营口市、抚顺市、盘锦市、铁岭市、通化市、辽源市、四平市、长春市、吉林市、白山市和延边州。资料年限从1961年至2011年。
各年代旬、月平均最低气温距平:利用各观测站旬、月最低平均气温计算得到各市(州)平均最低气温,利用1981年至2010年的气候平均计算出最低气温距平,分别计算1961—1970、1971—1980、1981—1990、1991—2000、2001—2010年的距平平均得到各年代旬、月平均最低气温距平。东北地区总平均指上述所指出的16个市(州)的平均。
同理计算各年代夏季6月、7月、8月最高气温距平值,各年代4月份降水距平百分率和各年代5月、6月空气相对湿度距平值。
2 气候变化对日本松干蚧扩散的影响
2.1 最冷月各旬最低气温和极端最低气温变化对日本松干蚧越冬及扩散的影响
2.1.1 最冷月各旬最低气温变化
1月份是东北地区最冷的月份,日本松干蚧能否安全越冬决定第2年的发生程度,文中分别分析了1月上中下旬和月平均最低气温的变化规律。各年代(1961—1970、1971—1980、1981—1990、1991—2000、2001—2010)1月上中下旬及月平均最低气温距平变化规律见图1—图4。
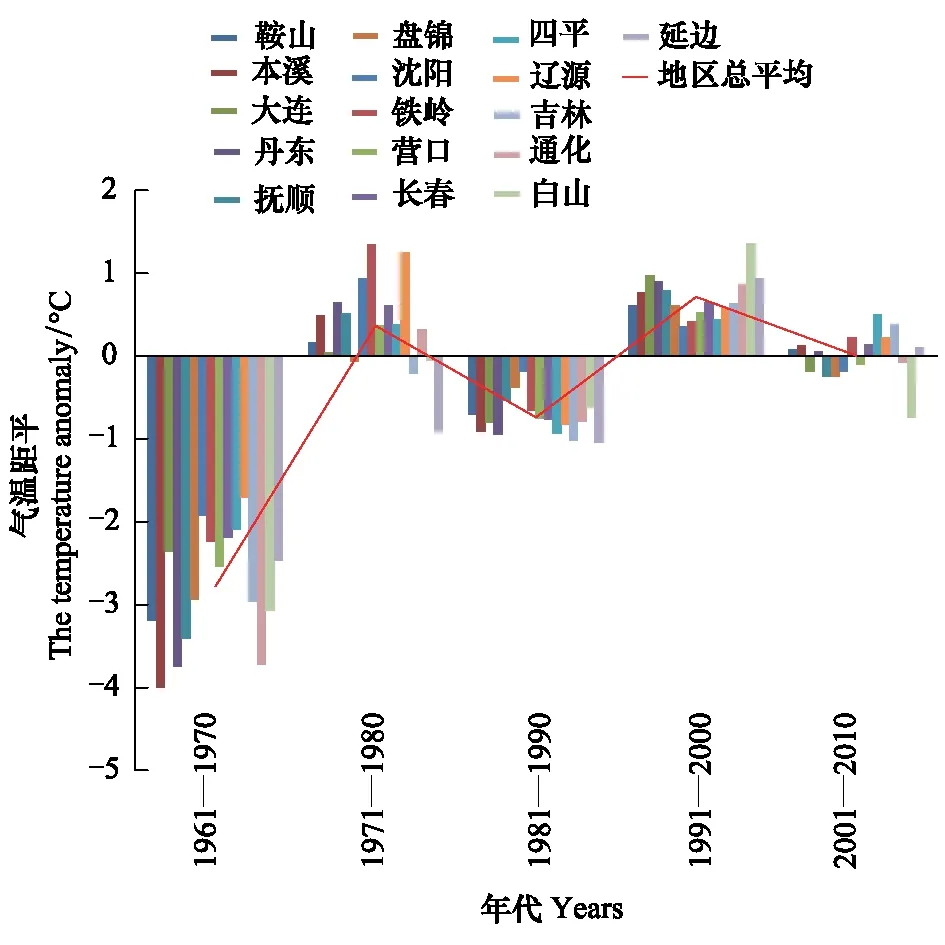
图1 各年代1月上旬最低气温距平变化趋势 Fig.1 The change trend of the mean-minimum-temperature anomaly the first ten-day of January
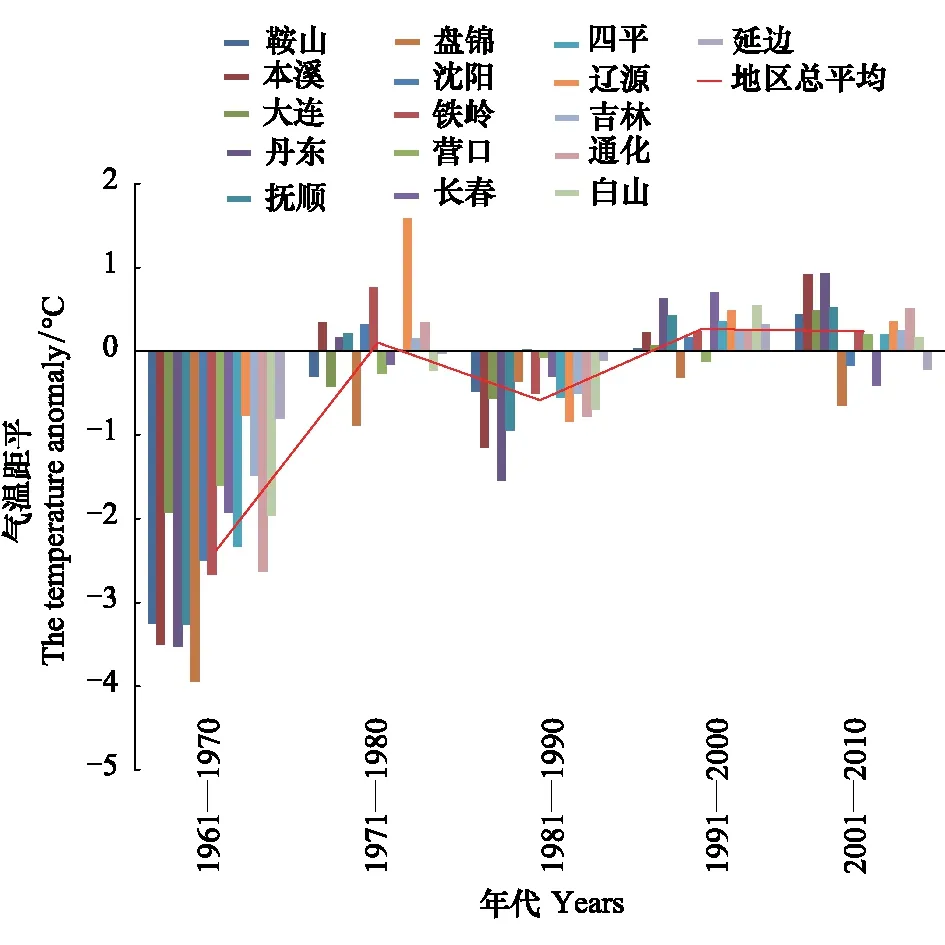
图2 各年代1月中旬最低气温距平变化趋势Fig.2 The change trend of the mean-minimum-temperature anomaly of the second ten-day of January
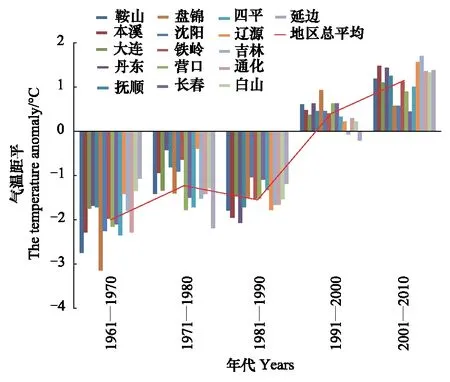
图3 各年代1月下旬最低气温距平变化趋势Fig.3 The change trend of the mean-minimum-temperature anomaly of the last ten-day January
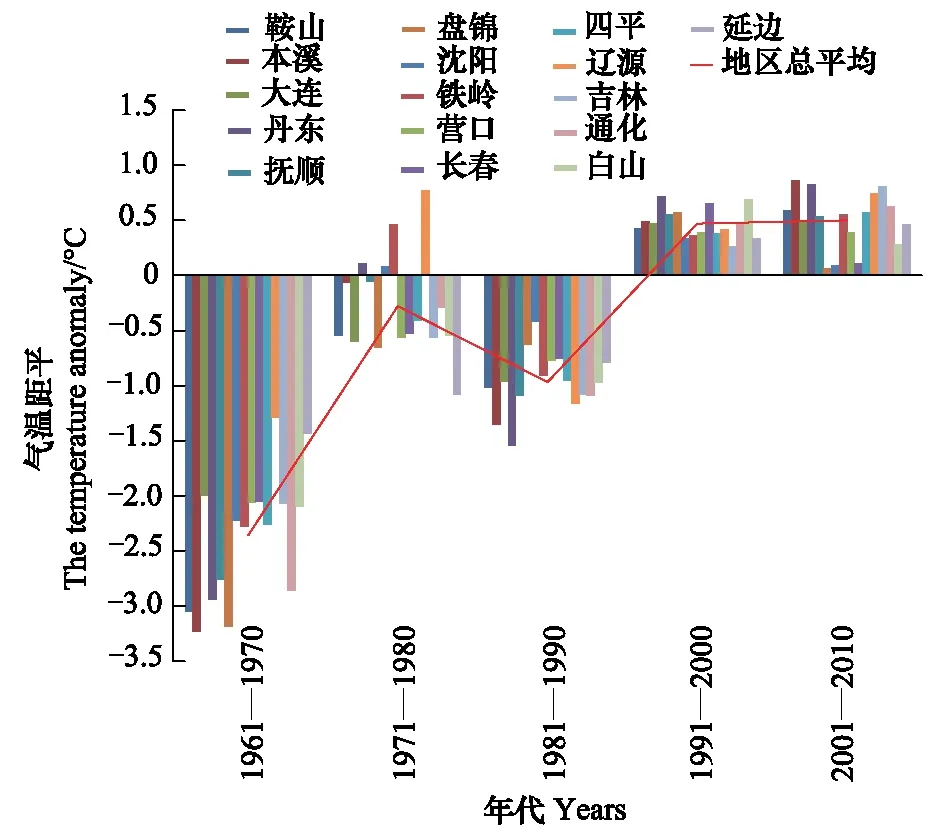
图4 各年代1月份最低气温距平变化趋势Fig.4 The change trend of the mean-minimum-temperature anomaly of January
由图1—图4可知,1月上旬和中旬最低气温升温不显著(P上旬>0.05,P中旬>0.05),下旬和月最低气温升温趋势明显(r下旬=0.93,P下旬<0.01,r1月=0.86,P1月<0.05)。20世纪60年代和80年代,1月各旬和1月份最低气温距平都为负距平,最低气温较低;20世纪70年代1月上旬多为正距平,中旬正距平的市略多于负距平,下旬都为负距平,1月份最低气温距平负距平多、正距平少;到90年代和21世纪的第1个10a多为正距平,最低气温较高。从分析结果可以看出,60年代,气候比较寒冷,70年代最低气温升高,尤其是1月上中旬,80年代最低气温又转低,但较60年代距平绝对值明显减小,从90年代开始,1月下旬和1月最低气温都明显上升。
2.1.2 冬季极端最低气温变化
东北地区极端最低气温距平年代变化规律如图5。
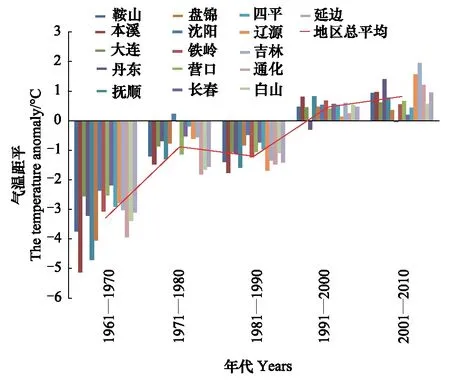
图5 东北地区各年代冬季极端最低气温距平变化趋势Fig.5 Change trend of the extreme-minimum-temperature anomaly in winter every ten-year in northeast China
由图5可见,20世纪90年代以前极端最低气温大都为负距平,年极端最低气温较低,尤其是60年代负距平值较大,70年代和80年代距平绝对值较60年代小,到90年代多转为正距平,本世纪前10年正距平值较大。由此可以看出,东北地区极端最低气温20世纪60年代以来升温显著(r=0.93,P<0.01)。
2.1.3 1月各旬、月最低气温和极端最低气温变化对日本松干蚧扩散的影响
日本松干蚧1942年传入大连的旅顺,1958年蔓延至盖县、庄河、东沟一线[22],1966年进入鞍山以南,辽阳的吉洞、隆昌、八会,以及岫岩、凤城的宝山、东方红公社一带,1970年传到辽阳太子河以南各公社、凤城县、凤山以南区域。从受害区域冬季的气象条件分析(表1),在1970年以前,1月各旬的平均最低气温、旬最低气温和冬季极端最低气温都出现在宽甸。1970年日本松干蚧刚刚侵入宽甸南部的虎山镇、辽阳的太子河以南和凤城的南部,尚未到达宽甸县城,受日本松干蚧为害的旅顺、大连、庄河、瓦房店、丹东等地气候比较温暖,最冷月平均最低气温都在-16℃以上,1月上旬平均最低气温的最低值为-19.3℃,于1968年出现在丹东,1月中旬为-21.4℃,于2001年出现在庄河,1月下旬为-20.6℃,于1960年出现在瓦房店。冬季极端最低气温为-28.1℃,2000年出现在庄河。这样的低温天气对日本松干蚧的生存构不成威胁,因此,尽管20世纪60年代气候寒冷,该区冬季气象条件都适宜日本松干蚧生存,在食物丰富的区域不管以何种方式传入,日本松干蚧都能在当地生存并扩散。
进入70年代1月各旬最低气温和冬季极端最低气温明显回升,日本松干蚧大举北侵,据宽甸县森林病虫防疫监测站资料记载,日本松干蚧在宽甸的传播路径是:1970年在宽甸县虎山镇发现,1973年在杨木川发现,1975年在毛甸子、长甸、永甸、宽甸镇发现,1978年在灌水、双山子、大川头、硼海、红石等乡镇发现。全国四省松干蚧科研协作会议于1974年12月10—15日在辽宁省辽阳市召开[23],根据会议纪要,1974年辽阳市柳河公社、栗子园林场和华子林场已经遭受日本松干蚧为害。1975年传入本溪,抚顺地区于1975年在抚顺县海浪乡首次发现,1976年最北蔓延到沈阳市东陵区王滨公社,东北发展到抚顺大伙房水库、温道林场的台沟、以及本溪高官寨、牛心台的兴隆山、草河城、草河掌和宽甸县的大川头、双山子至长甸的拉古哨一线[22]。20世纪70年代日本松干蚧扩散范围广。
以宽甸为代表站分析20世纪70年代1月各旬最低气温、冬季极端最低气温变化(表2,图6)对日本松干蚧扩散的影响。
由表2可知,宽甸20世纪70年代,上旬最低气温1971年和1977年较低外,其它年份温度都较常年偏高;中旬最低气温1971—1974、1976、1978都不到-20℃,其他几年低于-20℃,但都高于-22℃,下旬也都高于-23℃,明显高于其前的多数年份。60年代宽甸1月最低气温平均为-21.6℃,70年代仅为-17.4℃,明显高于60年代,冬季极端最低气温1972—1979年都高于-30℃,明显高于60年代(图6)。
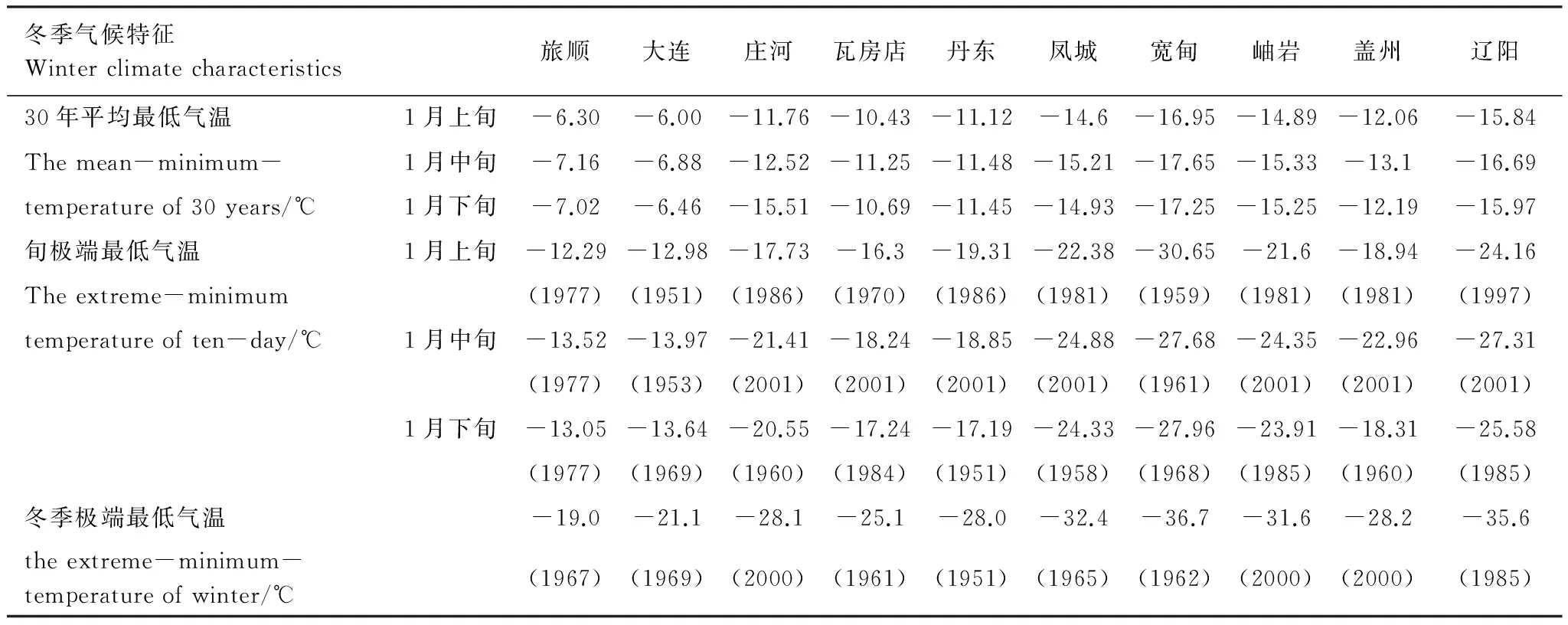
表1 日本松干蚧最早为害区冬季气候特征
括号内为最低气温出现的年份
表2 宽甸20世纪70年代1月各旬及1月份最低气温变化
Table 2 The mean-minimum-temperature change of every ten-day and January in 1970s in Kuandian
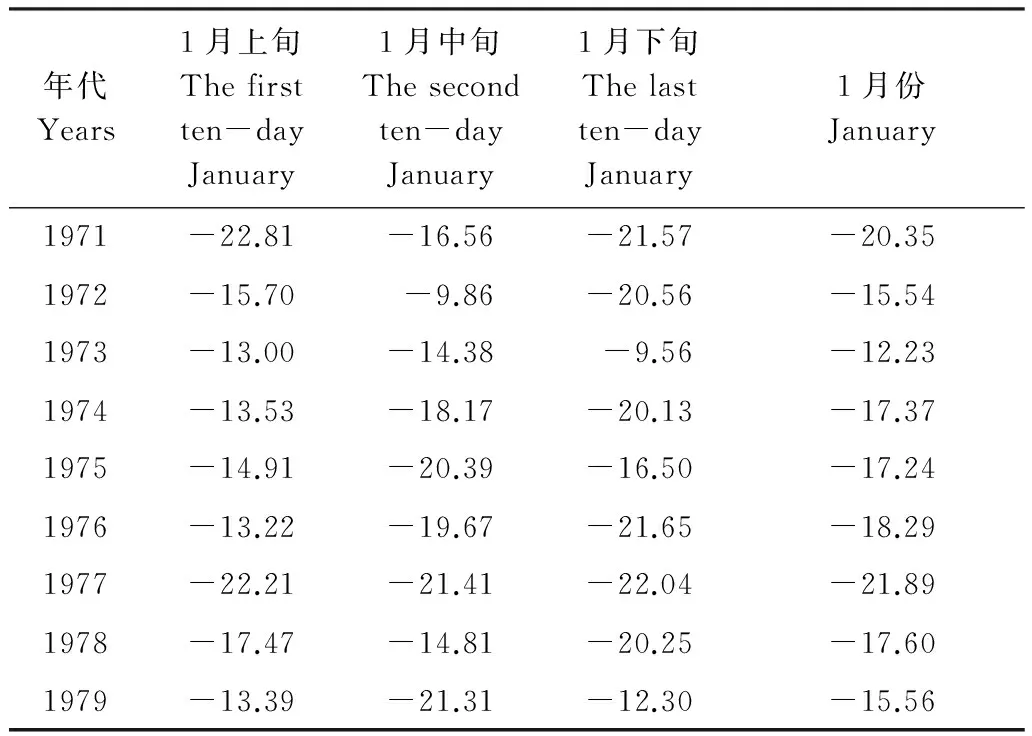
年代Years1月上旬Thefirstten-dayJanuary1月中旬Thesecondten-dayJanuary1月下旬Thelastten-dayJanuary1月份January1971-22.81-16.56-21.57-20.351972-15.70-9.86-20.56-15.541973-13.00-14.38-9.56-12.231974-13.53-18.17-20.13-17.371975-14.91-20.39-16.50-17.241976-13.22-19.67-21.65-18.291977-22.21-21.41-22.04-21.891978-17.47-14.81-20.25-17.601979-13.39-21.31-12.30-15.56
20世纪70年代和冬季极端最低气温明显升高,不但导致日本松干蚧向北扩散蔓延速度加快,同时危害程度加重。1982年6月调查[24],岫岩县前堡油松混交林树冠上截留大量的松干蚧卵囊,状如挂幡柳絮。说明在此之前的70年代,日本松干蚧在此生存并发生量极大,危害极重。日本松干蚧的快速扩散及大量发生与70年代1月最低气温和冬季极端最低气温明显升高有很大关系。
进入80年代1月最低气温较70年代明显下降(图7),日本松干蚧扩散速度降低,发生程度减轻。首届日本松干蚧学术讨论会于1984年12月在浙江省富阳县召开[25],会议纪要中指出,日本松干蚧除个别地区外大多数发生地区危害程度减轻,死树现象明显减少。沈夫君等[24]于1982—1984年在丹东市的油松林内设置30多块标准地进行连续调查,发现宽甸县中部、中东北部的松干蚧边界区和保护区,松干蚧蔓延速度与1976年以前相比明显减缓,经在宽甸县边界区的几年调查计算,每年仅向北、东北方向推移1—2km。说明20世纪80年代以前日本松干蚧为害严重,进入80年代为害减轻。
但自20世纪90年代以来,日本松干蚧的扩散蔓延速度加快,1994年在与辽宁接壤的吉林境内的东丰县、东辽县、梅河口市7个乡镇[2]发现了侵入的日本松干蚧疫点,1996年爆发[3],为害面积突然增大。90年代以后1月最低气温和冬季极端最低气温升温明显。东丰和梅河口90年代的1月最低气温较60年代明显升高,从1990年至1999年梅河口的1月最低气温都在-24℃以上。日本松干蚧的扩散总伴随着最冷月的气温升高,其爆发蔓延的时间和最低气温的高低高度相关,最低气温低的年份,日本松干蚧在原地为害或扩散的速度较慢,而最低气温高的年份日本松干蚧迅速向北、向东扩散或大范围爆发,为害程度重。

图6 宽甸冬季极端最低气温变化Fig.6 The extreme-minimum-temperature in winter in Kuandian
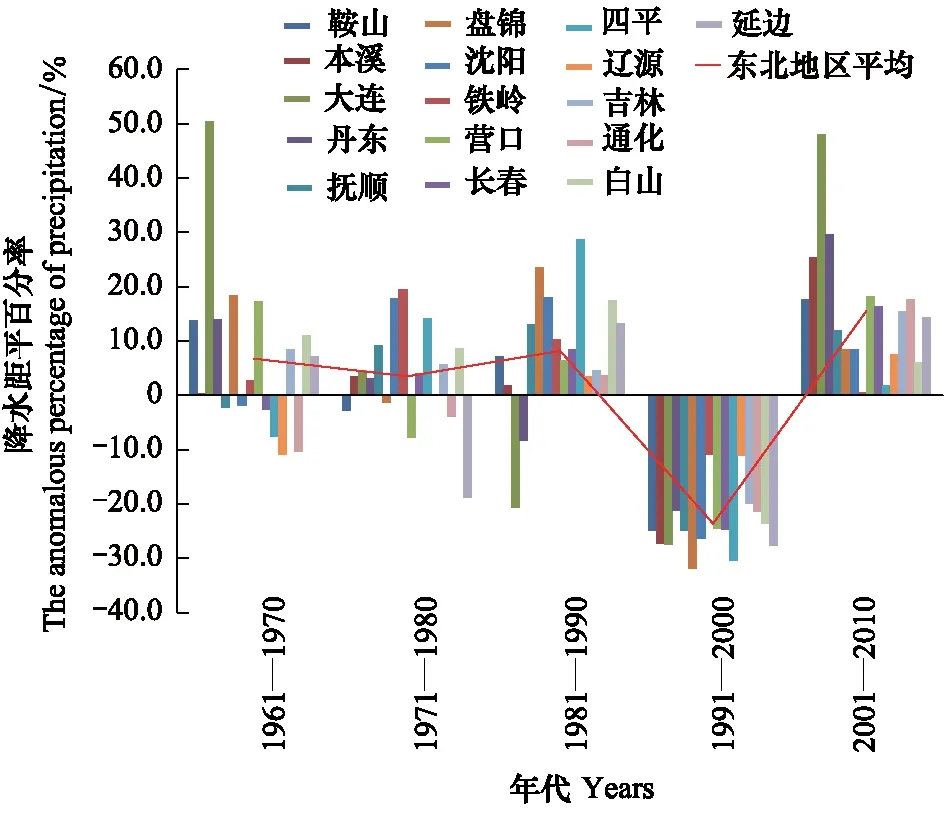
图8 东北地区4月份各年代降水距平百分率变化Fig.8 Anomalous percentage of precipitation in April in northeast China
2.2 降水变化对复苏后松干蚧若虫的影响
从历史4月降水数据(图8)来看,80年代以前,东北日本松干蚧为害区降水量多数市(州)偏多,到90年代各市(州)降水量全部偏少,到21世纪第1个10a,4月份降水量明显偏多。70年代和本世纪前10a降水偏多,日本松干蚧为害重,扩散速度快,90年代降水明显偏少,日本松干蚧扩散速度依旧很快,为害程度仍然很重,说明4月份降水量虽然对当年越冬代日本松干蚧的发生影响较大,但其年代变化对日本松干蚧在东北地区扩散却无明显影响。
2.3 越冬代卵孵化期湿度变化及影响
5、6月份日本松干蚧在东北地区自南向北陆续进入成虫产卵及卵孵化期,由图9、图10可知,5月份相对湿度从60年代到80年代呈上升趋势,对日本松干蚧扩散有利;90年代,辽宁各地湿度减小,吉林各地湿度增加,有利于吉林境内为害的日本松干蚧进一步扩散;21世纪前10a,辽宁省各地湿度增加,吉林各地湿度降低,但扩散速度并没有减缓。6月相对湿度呈先增后减的趋势。由历史资料可以看出,5月份空气相对湿度多在50%以上,6月份多在60%以上。且松树皮表面的湿度要比空气中的湿度大些,基本上能满足卵孵化的湿度需求。因此,空气湿度的年代变化对日本松干蚧生存扩散有影响,但影响不是很大,尤其是6月份的湿度。越夏代卵孵化期处于8、9月份,此期正处雨季或雨季刚过,植物茂盛,林间空气湿度大,不是制约卵孵化的主要气象影响因子,相对湿度年代变化也不是抑制或促使日本松干蚧在东北传播的主要影响因子。

图9 5月份相对湿度距平变化规律Fig.9 The variation of the relative-humidity anomaly in May
2.4 夏季最高气温变化对日本松干蚧扩散的影响
由图11可知90年代以前,6月份平均最高气温距平值多为负距平,90年代转为正距平,到21世纪的第1个10a,正距平值增大。60、70年代最高气温呈下降趋势,有利于日本松干蚧固定若虫生存及扩散,70年代以后呈上升趋势,理论上对日本松干蚧生存及扩散产生不利影响,但日本松干蚧扩散前沿区域的吉林境内6月份最高气温多在30℃以下,升温最明显的近10a最高气温多不超过33℃,且昼夜温差大、持续时间较短,因此6月份的高温年代变化对日本松干蚧扩散基本没有影响。
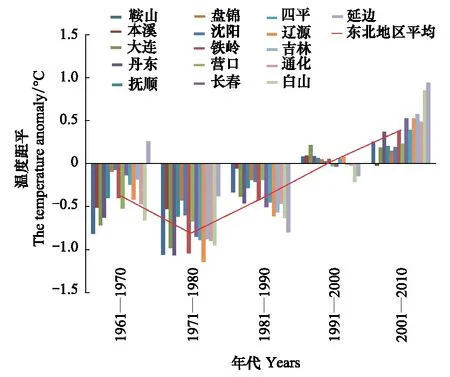
图11 6月份平均最高气温距平变化趋势Fig.11 The trend of the mean-maximum-temperature anomaly in June
7月份平均最高气温距平值70年代前和本世纪第1个10a规律不明显,有正距平有负距平,80年代出现明显一致负距平,90年代出现一致的正距平(图12)。80年代最高气温低,松干蚧扩散速度并未因此而加快,90年代气温高,扩散速度也并未因此而降低。说明7月份最高气温年代变化对日本松干蚧影响不大。由8月份平均最高气温变化(图13)可以看出,20世纪70年代下降剧烈,70年代至90年代明显升温,近20a最高气温较为平稳,理论上近20a持续的高温气候会对日本松干蚧的生存及扩散带来不利影响,实际情况通过受害区域内的四平观测站、长春观测站、吉林观测站和通化观测站极端最高气温变化趋势为例(图14)分析其变化对扩散的影响。
从长春、四平、吉林和通化4站历年极端最高气温变化情况看,极端最高气温变化趋势有所不同,有升温趋势,有降温趋势。自1951年以来,四平出现大于等于35℃的高温天气共13次(表3),1997年出现有资料记录以来的极端最高气温,为37.3℃,出现在6月28日,6月27—29日3d极端最高气温超过30℃,分别为33.6、37.3℃和33.6℃,而此3d的日平均气温分别为25.9、28.8℃和28.0℃,虽然出现了超过35℃的高温天气,但持续时间较短,未达到24h,且树林中的温度较百叶箱的温度低些,树皮下的温度更低些,对日本松干蚧固定若虫伤害较小。长春大于等于35℃的高温天气出现概率为23%(时间段1951—2012,下同),大于等于36℃的概率为8%;吉林大于等于35℃的高温天气的概率为20%,大于等于36℃出现的概率为7%;通化大于等于35℃的概率为7%,没有出现过大于36℃的高温天气。在东北地区极端高温天气持续时间短,最高气温超过36℃的很少,且昼夜温差较大,因此尽管东北地区极端最高气温总体有升温趋势,但目前夏季最高气温不是日本松干蚧在东北地区扩散的限制因素。
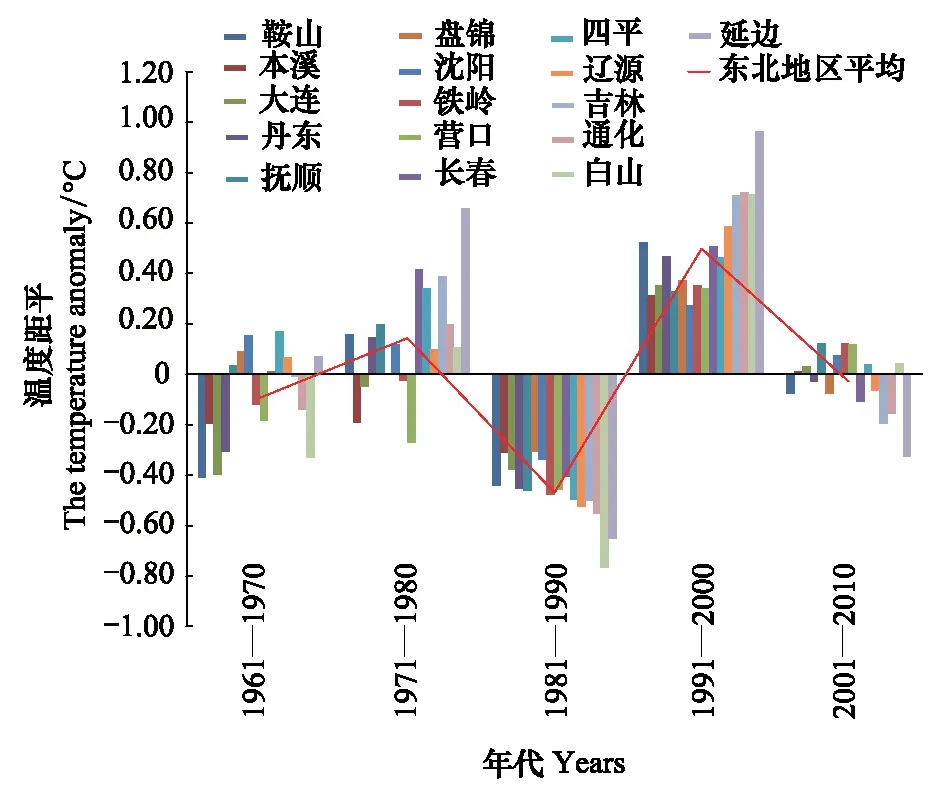
图12 7月份平均最高气温距平变化趋势Fig.12 The trend of the mean-maximum-temperature anomaly in July
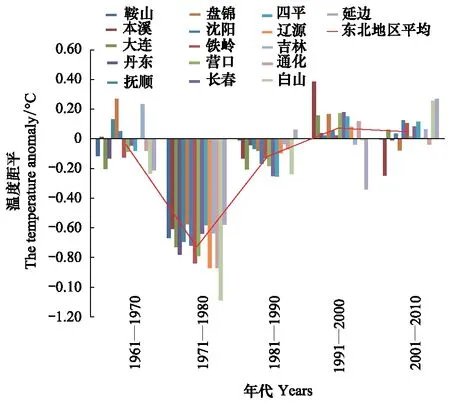
图13 各年代8月份平均最高气温距平变化Fig.13 The trend of the mean-maximum-temperature anomaly in August
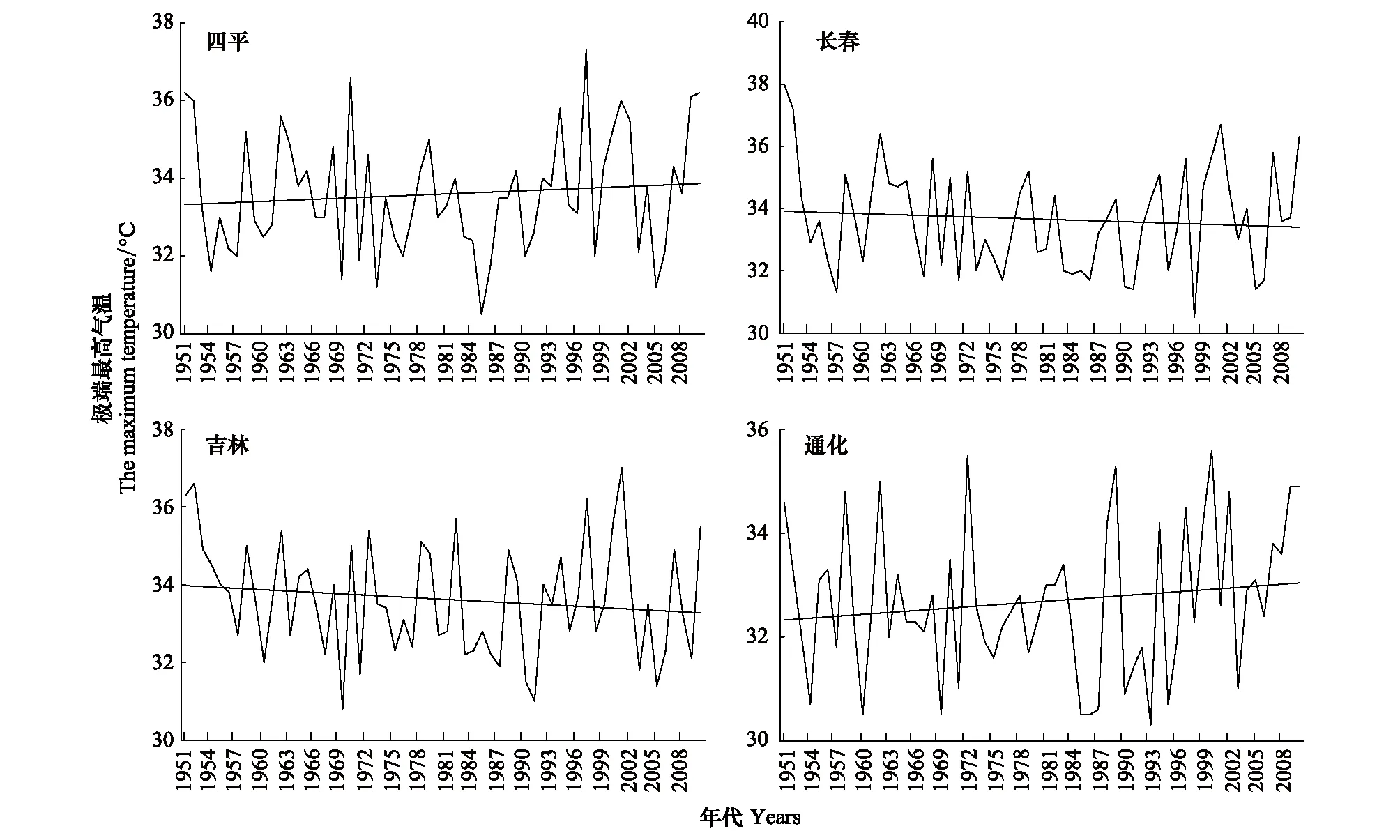
图14 夏季极端最高气温逐年变化Fig.14 Years change of the extreme-maximum-temperature in summer

表3 1951年以来极端最高气温分析
3 结论与讨论
3.1 结论
本文通过对日本松干蚧在东北地区传播路径与影响其生长发育及生存气象因子的年代变化分析得出:1)东北地区1月各旬及月最低气温的变化是影响日本松干蚧的在东北地区扩散的主要气象因子,1月最低气温低的年份日本松干蚧扩散较慢,1月最低气温高的年份扩散快。2)冬季极端最低气温的变化也是影响日本松干蚧扩散的主要影响因子,极端最低气温低的年份日本松干蚧扩散较慢,极端最低气温高的年份日本松干蚧扩散较快。1月最低气温和冬季极端最低气温升高是日本松干蚧在东北地区传播扩散的重要因素。3)日本松干蚧复苏后的降水量变化对日本松干蚧扩散影响不明显,卵孵化期的湿度变化和夏季最高气温的变化也不是日本松干蚧在东北地区扩散的主要影响因素。
3.2 讨论
日本松干蚧能传播蔓延首先它具有强大的繁殖能力。2000年冬季吉林省出现极其严寒天气,吉林省东丰县2001年1月10—17日最低气温在-36.0℃以下,其中11—16日最低气温为-40.7—-42.7℃,日本松干蚧在东丰的分布面积由2000年的3631.3 hm2剧减到2001年的310 hm2,到2002年春季日本松干蚧虫口又恢复到以往高密度状态,说明2001年夏秋季的气象条件适宜其越夏代的生长及繁殖,且其自身繁殖力十分惊人。其次,东北地区的风向有利于日本松干蚧继续向北传播。卵孵化期是日本松干蚧传播的主要时期之一,5、6月份为越冬代成虫产卵及卵孵化期,此期东北地区从春季的偏西风向转向偏西南风向;8、9月份为越夏代成虫产卵及卵孵化期,此期东北地区以西南风为主[26],这为日本松干蚧从辽宁向北传播到吉林境内和在吉林境内进一步扩散传播提供了可能。在吉林与辽宁接壤首次发现日本松干蚧的地点可以看出,山体为西北东南走向,以山脊为界南坡为辽宁清原,北坡为吉林梅河口。松干蚧疫情从南坡传到山顶后,可以通过风、雨水或自然扩散传到吉林境内,然后在吉林蔓延扩散。通过对日本松干蚧传播路径的实地考察,即使是玉米田中孤立的黑松林也受到日本松干蚧的为害,分析风等自然传播的可能性较大。即使是人为传播,也要有从疫木向被害木转移的过程,即便它有强大的繁殖能力,数量众多,这一过程对日本松干蚧来说也是一个遥远而艰难的过程,因此日本松干蚧的生存能力和传播能力不可小觑。
根据王石立[27]研究,东北气候变暖主要表现于冬季,最低温度升高幅度大于最高温度的升高幅度。气候变化除了对日本松干蚧自身生存的影响,还对其寄主松树及天敌有影响,据王婷等[28]研究,生长季开始时最低温度升高有利于延长生长季、与树木年轮宽度成正比。气候变暖有利于寄主松树的生长发育,为日本松干蚧提供了丰富的食源。异色瓢虫是日本松干蚧的天敌,气候变暖有利于异色瓢虫的生长发育和发生代数,对日本松干蚧的种群数量有一定抑制作用。另外,冬季气候变暖有利于日本松干蚧的发生,同时经历过极端气候事件如低温或其它不良气候条件存活下来的日本松干蚧对不良环境有一定的抵抗能力,自身对不利的气象条件适应能力提高,有利于日本松干蚧的生存,这些方面综合影响需进一步探究。本文只分析了气候因子变化对日本松干蚧扩散的宏观影响,为将来从气候角度分析预测日本松干蚧能否进一步向北扩散奠定基础,同时为日本松干蚧监测和防控提供参考。
[1] McClure M S, Ji L Z. The effect on temperature and host to Matsucoccusmatsumurae distribution in Asia and America.Journal of Liaoning Forestry Science and Technology, 1988, (5): 72- 75.
[2] Shan G M, Gao J C, Gao L J, RenLW, YuY P, Pi Z Q, Cao L J, Li F, Cao Y K. Occurrence and development trend ofMatsucoccusmatsumuraeand its control in Jilin, China. Jilin Forestry Scienceand Technology, 2003, 32(3): 23- 28.
[3] Zhong XW, Wang Z M, Pi Z Q. An analysis ofMatsucoccusmatsumuraedistribution status and its spread probability in Jilin Province.ChinaPlantProtection, 2008, 28(3): 27- 29.
[4] LiHM, Han HX, Xue DY. Prediction of potential geographic distribution areas for the pine bark scale,Matsucoccusmatsumurae(Kuwana) (Homoptera: Margarodidae) in China using GARP modeling system. ActaEntomologicaSinica, 2005, 48(1): 95- 100.
[5] Lin C G, Wang Z M, Ma S H, Li X F, Peng G Z, Zhang L. The risk analysis ofMatsucoccusmatsumuraedispersal.Journal of Liaoning Forestry Science and Technology,2011, (5): 42- 44.
[6] Wang Z M, Du J S, ZhangL J, Zhong X W, Pi Z Q. The Analysis for the Pine Bark Scale,Matsucoccusmatsumurae(Kuwana)Invasion and Spread Possible in the Northeast China.Chinese Agricultural Science Bulletin, 2009, 25(10): 107- 111.
[7] Fan S R, Wang M L, Hu R J, Hu H L, Yang M D, Qiu X J, Lu M H, Du Z Q. The preliminary study onMatsucoccusmatsumuraedispersal path.Forest Pest and Disease,1984, (1): 6- 8.
[8] Xu F Y, Li G W. Effects of temperature, humidity and rainfall toMatsucoccusmatsumuraeparasitized nymphs. Practical Forestry Technology, 1985, (11): 24- 27.
[9] Zhou Y J, Zhang X D, Guo A H, Xiao Y, Zhao Y R. Application of the effective accumulated temperature model in the forecast of the occurrence period ofMatsucoccusmatsumuraenymph under natural conditions.Forest Pest and Disease, 2012, 31(4): 15- 17.
[10] Xiao Y, Fan X H, Zhao Y R, Huang F Y, Wang L R. Application of method of forecasting outbreak ofMatsucoccusmatsumurae. Journal of Liaoning Forestry Science and Technology,1999, (1): 31- 34.
[11] Cao L J. Study on disease prevention, interdiction and silvicultural measures ofMatsucoccusmatsumuraein Jilin province //Alien Pest Quarantine and Control Technology Symposium Proceedings. Nanjing: Entomological Society of China, Chinese PhytopathologicalSociety, 2005: 159- 165.
[12] Yuan F X, Liu S, Hu Y Q, Zhang Y S, Zhang T L. Effect of meteorological condition on overwintering mortality rate ofMatsucoccusmatsumurae.Chinese Journal of Agrometeorology,2012, 33(2): 310- 314.
[13] Jing XH, Kang L. Research progress in insect cold hardiness. ActaEcologicaSinica, 2002, 22(12): 2202- 2207.
[14] Jing XH, Kang L. Overview and evaluation of research methodology for insect cold hardiness. EntomologicalKnowledge, 2004, 40(1): 7- 10.
[15] Li B X, Chen Y L, Cai H L. Supercooling and insect cold hardiness. Entomological Knowledge, 1998, 35(6): 361- 364.
[16] Mei Z X, Li J Q. The physiological mechanism and influential factors of insect cold tolerance.Journal of Binzhou University, 2006, 22(3): 57- 61.
[17] Chen Y, Ma C S. Effect of global warming on insect: a literature review. ActaEcologicaSinica, 2010, 30(8): 2159- 2172.
[18] WaltersRJ, HassallM, TelferMG, HewittG M, PalutikofJP. Modelling dispersal of a temperate insect in a changing climate. Proceedings of the Royal Society B: Biological Sciences, 2006, 273(1597): 2017- 2023.
[19] Fischer D, Moeller P, Thomas SM, Naucke TJ, Beierkuhnlein C. Combining climatic projections and dispersal ability: amethod for estimating the responses of sandflyvector species to climate change. PLoS Neglected Tropical Diseases, 2011, 5(11): e1407.
[20] Jaeschke A, Bittner T, Reineking B, Beierkuhnlein C. Can they keep up with climate change?---Integrating specific dispersal abilities of protected Odonata in species distribution modelling. Insect Conservation and Diversity, 2013, 6(1): 93- 103.
[21] RobinetC, RoquesA, PanH, Fang G F, Ye J R, Zhang Y Z, Sun J H. Role of human-mediated dispersal in the spread of the pinewood nematode in China. PLoS ONE,2009, 4(2): e4646.
[22] Matsucoccus matsumurae Prevention Trials Cooperative Group in Liaoning Province. The investigation in propagation law ofMatsucoccusmatsumurae.Journal of Liaoning Forestry Science and Technology,1977, (3): 8- 14.
[23] The whole nation four provinces cooperation researchconference summary ofMatsucoccusmatsumurae. Journal of Liaoning Forestry Science and Technology,1975, (1): 8- 9.
[24] Shen F J, Hui B N, Hou Y. To exploreMatsucoccusmatsumuraedamage characteristics and control method in Dandong.Journal of Liaoning Forestry Science and Technology,1987, 5(4): 34- 45.
[25] Li y C.Matsucoccusmatsumuraecooperative research group held a symposium. Forest Pest and Disease,1985(1): 24- 24.
[26] Wang H Q, Wu Z Y. Jilin Climate. Beijing: Meteorological Press, 1997: 75- 82.
[27] Wang S L, Zhuang L W, Wang F T. Impacts of climate warming on thermal and moisture conditions in northeast China in recent 20 years. Journalof Applied Meteorological Science,2003, 14(2): 152- 164.
[28] Wang T, Yu D, Li J F, Ma K P. Advances in research on the relationship between climatic change and tree-ring width. ActaPhytoecologicaSinica,2003, 27(1): 23- 33.
参考文献:
[1] McClure M S, 姬兰柱. 在亚洲和北美温度与寄主对日本松干蚧分布的有效影响. 辽宁林业科技, 1988, (5): 72- 75.
[2] 山广茂, 高峻崇, 高立军, 任利伟, 于艳萍, 皮忠庆, 曹丽君, 李飞, 曹玉坤. 日本松干蚧疫情在吉林省发生发展趋势及治理情况分析报告. 吉林林业科技, 2003, 32(3): 23- 28.
[3] 钟晓巍, 王志明, 皮忠庆. 日本松干蚧在吉林省的分布现状及其扩散可能性分析.中国植保导刊, 2008, 28(3): 27- 29.
[4] 李红梅, 韩红香, 薛大勇. 利用GARP生态位模型预测日本松干蚧在中国的地理分布.昆虫学报, 2005, 48(1): 95- 100.
[5] 蔺成阁, 王志明, 马书华, 李秀峰, 彭广增, 张来. 日本松干蚧扩散的危险性分析. 辽宁林业科技, 2011, (5): 42- 44.
[6] 王志明, 杜继生, 张利军, 钟晓巍, 皮中庆. 日本松干蚧对东北地区的入侵及其蔓延可能性分析. 中国农学通报2009, 25(10): 107- 111.
[7] 樊尚仁, 王美莲, 胡润吉, 胡鹤龄, 杨牡丹, 裘学军, 卢苗海, 杜增庆. 日本松干蚧传播途径的初步研究. 中国森林病虫,1984, (1): 6- 8.
[8] 徐福元, 李广武. 温、湿度和降雨对日本松干蚧寄生若虫的影响. 林业实用技术, 1985, (11): 24- 27.
[9] 周玉江, 张旭东, 郭安红, 肖艳, 赵彦荣.自然变温有效积温模型研究及在日本松干蚧若虫发生期预测上的应用.中国森林病虫, 2012, 31(4): 15- 17.
[10] 肖艳, 范学恒, 赵彦荣, 黄凤玉, 王连荣.日本松干蚧发生期预测预报方法的应用研究.辽宁林业科技, 1999, (1): 31- 34.
[11] 曹丽君. 吉林省日本松干蚧疫情预防、封锁及营林措施的研究 //外来有害生物检疫及防除技术学术研讨会论文汇编. 南京: 中国昆虫学会, 中国植物病理学会, 2005: 159- 165.
[12] 袁福香, 刘实, 胡艳全, 张玉书, 张铁利. 气象条件对日本松干蚧越冬死亡率的影响.中国农业气象, 2012, 33(2): 310- 314.
[13] 景晓红, 康乐. 昆虫耐寒性研究. 生态学报, 2002, 22(12): 2202- 2207.
[14] 景晓红, 康乐. 昆虫耐寒性的测定与评价方法. 昆虫知识, 2004, 40(1): 7- 10.
[15] 李冰祥, 陈永林, 蔡惠罗. 过冷却和昆虫的耐寒性. 昆虫知识, 1998, 35(6): 361- 364.
[16] 梅增霞, 李建庆. 昆虫抗寒性的生理机制及影响因子. 滨州学院学报, 2006, 22(3): 57- 61.
[17] 陈瑜, 马春森. 气候变暖对昆虫影响研究进展. 生态学报, 2010, 30(8): 2159- 2172.
[22] 辽宁省松干蚧防治试验协作组. 松干蚧的蔓延及传播规律调查. 辽宁林业科技, 1977, (3): 8- 14.
[23] 全国四省松干蚧科研协作会议纪要. 辽宁林业科技, 1975, (1): 8- 9.
[24] 沈夫钧, 回柏年, 侯义. 丹东地区日本松干蚧为害特点及防治方法的探讨.辽宁林业科技, 1985, (4): 34- 45.
[25] 李永成. 日本松干蚧科研协作组举行学术讨论会.中国森林病虫.1985, (1): 24- 24.
[26] 王惠清, 吴忠义. 吉林省气候. 北京: 气象出版社, 1997: 75- 82.
[27] 王石立, 庄立伟, 王馥棠. 近20年气候变暖对东北农业生产水热条件影响的研究.应用气象学报, 2003, 14(2): 152- 164.
[28] 王婷, 于丹, 李江风, 马克平. 树木年轮宽度与气候变化关系研究进展. 植物生态学报, 2003, 27(1): 23- 33.
The impact of climate change on matsucoccus matsumurae dispersal in Northeast of China
YUAN Fuxiang1,*, LIU Shi1, HU Yanquan1, ZHANG Yushu2, DAI Yong3,CAO Rui3
1MeteorologicalScienceInstituteinJilin,Changchun130062,China2InstituteofAtmosphericEnvironmentChinaMeteorologicalAdministration,Shenyang110016,China3TheForestDiseaseandPestControlandQuarantineStationofMeihekouofJilinProvince,Meihekou135000,China
Based on the material of climate and Matsucoccus matsumurae dispersal, according to the dispersal areas and the climate characteristics of the place infested, the effect was analyzed about the main meteorological factors to Matsucoccus matsumurae dispersal in the northeastern China. The results showed that: the mean-minimum-temperature of every ten-day and month of January presented the increasing trend (r=0.86,P<0.05). The extreme-minimum-temperature in winter increased(r=0.93,P<0.01) slowly,too. The changes in temperature of the years had something to do with Matsucoccus matsumurae dispersal in the northeast of China. The more rapidly was Matsucoccus matsumurae dispersed, the higher had the pest population density and had the more serious extent of injury when the mean-minimum-temperature and the extreme-minimum-temperature in January were increased evidently in the 1970s and the 1990s of last century. Therefore there was a significant correlation between the minimum-temperature and Matsucoccus matsumurae dispersal. The higher the ten-year of the mean-minimum-temperature was, the faster Matsucoccus matsumurae dispersal was. The lower the minimum-temperature was, the slower its dispersal was. The great range of dispersal and outbreaking of Matsucoccus matsumurae took place in the years of high minimum-temperature in January. The minimum-temperature in January and the extreme-minimum-temperature in winter increasing were the main factors of Matsucoccus matsumurae dispersal. There was no relation between dispersal and variation of precipitation after conscious, air relative humidity and the maximum summer temperature.
Matsucoccus matsumurae;dispersal; climate change; northeast of China
国家自然科学基金(41175100); 公益性行业(气象)科研专项(GYHY200906028)
2013- 02- 17; 网络出版日期:2014- 03- 17
10.5846/stxb201302170272
*通讯作者Corresponding author.E-mail: yfx1998@sina.com
袁福香, 刘实, 胡艳全, 张玉书, 戴勇, 曹蕊.气候变化对中国东北地区日本松干蚧传播扩散的影响.生态学报,2014,34(22):6711- 6721.
Yuan F X, Liu S, Hu Y Q, Zhang Y S, Dai Y,Cao R.The impact of climate change on matsucoccus matsumurae dispersal in Northeast of China.Acta Ecologica Sinica,2014,34(22):6711- 6721.
