贵州南部水稻田白背飞虱种群数量动态与分析
2014-08-10薛文鹏金道超
薛文鹏, 金道超, 杨 洪
(贵州大学昆虫研究所,贵州山地农业病虫害重点实验室,贵阳 550025)
调查研究
贵州南部水稻田白背飞虱种群数量动态与分析
薛文鹏, 金道超, 杨 洪*
(贵州大学昆虫研究所,贵州山地农业病虫害重点实验室,贵阳 550025)
采用灯光诱集和田间系统调查的方法,对贵州南部三都县2008-2012年的白背飞虱灯下和田间种群进行了分析研究,结果显示:白背飞虱在4月中下旬开始迁入贵州南部,在5月中下旬到达迁入高峰;灯下主峰日与灯下始见日(r=0.996**)和始盛日(r=0.997**)均呈显著相关;灯下种群中雌虫数量略高于雄虫,年度间雌、雄性比差异不显著。若虫是田间种群的主要构成部分,在6月上旬达到发生高峰,其发生为害期较成虫短,一般到7月中下旬结束;成虫发生期贯穿水稻的整个生育期,在5月下旬至6月上旬出现高峰。田间主要为害世代的发生量与6月中下旬的灯下虫量呈显著正相关。
白背飞虱; 灯诱种群; 田间种群; 贵州南部
水稻是贵州省重要的粮食作物,2011年贵州省水稻种植面积为681 530 hm2,产量达3 039 300 t,占全省粮食作物总产量的34.7%,在各种粮食作物中位居第一;贵州南部则是贵州水稻的主要产区:2011年贵州南部(黔南布依族自治州,下称黔南州)与东南部(黔东南苗族侗族自治州,下称黔东南州)水稻种植面积为235 720 hm2,占到全省水稻种植总面积的34.6%[1],在全省的粮食供给中占有举足轻重的地位,黔南地区处于云贵高原东南部向广西丘陵过渡的斜坡地带,终年受海洋暖流气候影响,夏季湿热多雨,加之独特的喀斯特立体气候特点,极利于白背飞虱的发生,白背飞虱[Sogatellafurcifera(Horváth)](white-backed planthopper, WBPH)属半翅目(Hemiptera)飞虱科(Delphacidae),其刺吸水稻,使水稻生长缓慢,分蘖延迟,瘪粒增加,为害严重时,造成稻株枯死[2],此外白背飞虱还是南方水稻黑条矮缩病毒(Southernriceblack-streakeddwarfvirus, SRBSDV)的传播媒介[3],这种病毒病更是给水稻种植带来了毁灭性的打击。
作为一种迁飞性昆虫,白背飞虱每年春季随西南气流由中南半岛进入我国,而后逐代向北迁飞,至秋季又从北往南回迁[4-6],同时我国的稻飞虱也可向日本、韩国迁飞[7-8],但是详尽的区域性迁飞规律尚不明确;在迁飞过程中,昆虫遇到降雨、下沉气流、风向幅合等外力作用时会被集中迫降;飞行能源物质耗尽或温度骤变至低于其飞行温度阈值时也会选择主动降落[9],这种难以预测的大规模降虫,非常有利于白背飞虱的暴发式发生,严重时甚至可以落地成灾,让人防不胜防,为明确其灯下和田间种群动态,从而有效地预防白背飞虱暴发并控制其为害,本文在黔南州代表稻区——三都水族自治县(下称三都县)开展了白背飞虱的灯光诱集和田间系统调查工作,并对近五年(2008-2012年)的白背飞虱灯下和田间种群进行了分析研究。
1 材料与方法
1.1 材料与试验点
当地主栽品种:‘中优5617’,成都国康农作物研究所;‘Ⅱ优63’,湖南省水稻杂交中心;‘黔优98’,贵州省水稻研究所;‘内香优18’,德农正成种业有限公司。
试验点设在三都县三合镇猴场村,海拔390 m,属亚热带温润季风气候,水田一年一熟,一季中稻占主要比重,每年清明后播种育秧,5月上旬至下旬移栽大田,8月下旬至9月初黄熟收割。
化学防治选用农药:25%噻嗪酮(buprofezin)可湿性粉剂,河北万特生物化学有限公司;5%啶虫脒(acetamiprid)乳油,开封市普朗克生物化学有限公司;80%敌敌畏(dichlorvos)乳油,南通江山农药化工股份有限公司;25%杀虫双(bisultap)水剂,安徽华星化工股份有限公司;1.8%阿维菌素(abamectin)水剂,郑州先利达化工有限公司;30%井冈霉素(validamycin)水剂,浙江农友化工有限公司。
1.2 方法
1.2.1 诱虫灯设置
采用佳多牌频振式自动虫情测报灯,以200 W白炽灯为诱虫光源;将测报灯安装固定在高60 cm,宽110 cm的基台上,远离其他光源;每天黄昏时段开灯,次日清晨关灯,逐日取虫,并进行室内鉴别,记录白背飞虱的雌、雄成虫数量;记录每日天气及气温情况:采用中央气象台(http:∥www.nmc.gov.cn)的每日天气数据;测报灯每年3月1日启用,11月中旬左右(未见虫10 d后)停用。
1.2.2 系统调查田设置和调查方法
系统调查田设置在三合镇猴场村。于常规稻作区设置系统观察田2块(早栽、晚栽类型田各一块,早栽田5月上旬移栽,晚栽田5月下旬移栽),分蘖期至灌浆期每隔10~20 d由当地农户进行正常施药防治,按药剂说明进行单施或混合喷施;设置预测圃2块(早栽田和晚栽田各1块)作为对照,在水稻生育期内不进行施药行为,用于观测自然条件下白背飞虱的田间种群动态。
采用《贵州省稻飞虱监测技术规范》[10](以下简称《规范》)的标准,从移栽返青后开始,每5 d调查1次,直至水稻黄熟收割结束。调查方法采用盘拍法,用白瓷盘(33 cm×45 cm)进行平行跳跃式取样。调查时,在盘内均匀涂上机油,将盘轻轻地倾放在稻秆基部,与稻秆成45°角,快速拍打稻株,连拍3次,立即端起检查计数,每块田调查10点,每点查2~5丛(视虫量而定,虫多少查,虫少多查),详细记录白背飞虱低龄若虫(1~3龄)、高龄若虫(4~5龄)、长翅型成虫和短翅型成虫的数量,然后折算成百丛虫量。
1.2.3 白背飞虱发生世代的划分
采用《规范》的世代划分标准,即4月20日以前发生的划分为第1代;4月21日至5月20日为第2代;5月21日至6月20日为第3代;6月21日至7月20日为第4代;7月21日至8月20日为第5代;8月21日至9月20日为第6代;9月21日至10月20日第7代;10月21日以后划分为第8代。
1.3 数据处理
采用Microsoft Excel 2003进行统计量的整理计算并绘制图表,使用SPSS 17.0对数据进行差异性及相关性分析。
2 结果与分析
2.1 白背飞虱灯诱种群
2.1.1 白背飞虱灯诱种群的始见日、末见日及年度总虫量
三都县无本地越冬虫源,因此白背飞虱灯下始见即表明外地虫源开始迁入,灯诱资料显示:2010年白背飞虱灯下始见日最早,为3月6日;2011年最晚,为4月25日,两者相差近2个月。始见日4次出现在4月份,其中4月下旬2次,上、中旬各1次。
末见日多为向境外回迁的白背飞虱上灯,集中在10月(3次)和11月(2次),最早的年份是2012年,在10月上旬,最晚的年份是2011年,在11月中旬,较2012年晚1月有余。另外,无论始见日还是末见日,其灯下虫量都较少,少则1头,多不过24头(表1)。
表12008-2012年三都县白背飞虱灯下各发生期及诱虫量1)
Table1OccurrencedateandlighttrapcatchesofWBPHinSanduCountyfrom2008to2012
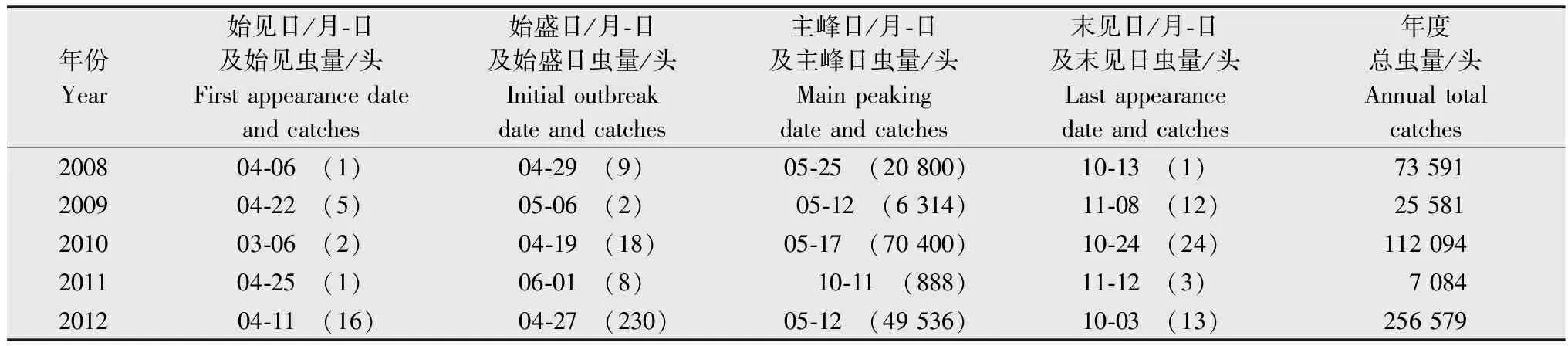
年份Year始见日/月⁃日及始见虫量/头Firstappearancedateandcatches始盛日/月⁃日及始盛日虫量/头Initialoutbreakdateandcatches主峰日/月⁃日及主峰日虫量/头Mainpeakingdateandcatches末见日/月⁃日及末见日虫量/头Lastappearancedateandcatches年度总虫量/头Annualtotalcatches200804⁃06 (1)04⁃29 (9) 05⁃25 (20800)10⁃13 (1)73591200904⁃22 (5)05⁃06 (2)05⁃12 (6314)11⁃08 (12)25581201003⁃06 (2)04⁃19 (18) 05⁃17 (70400)10⁃24 (24)112094201104⁃25 (1)06⁃01 (8)10⁃11 (888) 11⁃12 (3) 7084201204⁃11 (16) 04⁃27 (230)05⁃12 (49536)10⁃03 (13)256579
1) 表中括号里的数字是诱到白背飞虱的数量。
The figures in brackets in the table were light trap catches of WBPH.
灯下诱虫总量可以在一定程度上反映田间的实际发生程度,2012年三都县白背飞虱灯下诱虫总量最多,达到256 579头(表1),分别是2008、2009、2010、2011年的3.5倍、10.0倍、2.3倍和36.2倍,而2011年全年灯下诱虫量不过万,由此可以看出,2012年白背飞虱灯下发生最为严重,而2011年白背飞虱发生较轻。
2.1.2 白背飞虱灯诱种群的始盛日和主峰日
始盛日是指从该日起连续多日(5日以上)均能诱获白背飞虱,且诱获白背飞虱数量有逐日增加的趋势。通过整理分析:2010年白背飞虱灯下始盛日出现最早,在4月19日,而2011年白背飞虱灯下始盛日最晚,在6月1日,各年份从始见日到始盛日的间隔天数也不尽相同,2009年间隔最短(13 d),2010年间隔最长(42 d),平均间隔25.6 d,从近五年的数据看,始见日出现较早的年份,始盛日出现的时间也较为提前(表1)。
5年之中,除2011年以外,白背飞虱灯下主峰日均出现在5月份,最早在5月12日,最晚在5月25日,主峰日与始见日(r=0.996**)、始盛日(r=0.997**)均呈显著正相关。2010年主峰日的诱虫数量最多,达到70 400头,接近其余4年主峰日诱虫量的总和,分别是2008年、2009年、2011年和2012年的3.4倍、11.1倍、79.3倍和1.4倍(表1)。2011年主峰日出现在10月11日,较往年大大推迟,且诱获虫量也急剧减少,究其原因,是因为2011年上半年整体温度偏低,低温屏障使白背飞虱的迁入大幅受挫,基本无大的迁入峰降至三都,导致灯下无明显诱虫高峰,直至10月中旬,才出现相对较大的虫峰。但此时三都全境水稻收割完毕,该虫峰只能是向境外回迁的越冬种群。
2.1.3 白背飞虱灯诱各世代的发生数量
从灯诱数据来看,白背飞虱在三都县一年可发生8代,灯下各代诱捕虫量间存在较大差异。第1~4代主要是迁入代:其中第1代为零星发生,灯下诱获数量较少,诱获最多的年份仅474头(2010年),其余年份第1代的诱获数量均不过百;迁入虫量主要集中在第2、3代,2008年主迁入代为第3代(56 598头),2009、2010、2012年主迁入代是第2代(14 167头、95 574头、218 406头),2011年较晚,主迁入代推迟至第4代(1 265头)。
第5代多为本地迁出代,不同年份迁出代的虫量差异较大,最少的是2010年(55头),最多的是2012年(915头);第6代灯下发生量(平均1 409头)显著多于第5代(平均434头),但此时当地水稻已进入生长末期,个别早收田块已经收割完毕,由于食料条件的恶化,本地虫源显然不能提供如此多的上灯虫量,所以第6代上灯的虫量包含了本地外迁和向南回迁两种虫源性质,且以向南回迁的虫源性质为主。
第7~8代为向南回迁的过境虫源,第8代零星发生,虫量主要集中在第7代,其中2011年的诱集虫量最多,表明其回迁种群的规模也大于其他年份(图1)。

图1 2008-2012年三都县白背飞虱各世代灯下发生数量动态
2.1.4 白背飞虱灯诱各世代的性别结构
性别结构是决定种群繁殖的重要因素,对于前期迁入为害的白背飞虱而言,灯下性别结构尤为重要,是决定其田间发生的重要指标之一,白背飞虱的灯下迁入数量主要集中在第2~4代,可以认为这3个世代是灯下主发生代,通过对各年份的(2011年虫量较少,故略)3个灯下主发生代进行分析,得出年度间白背飞虱的灯下雌雄比无显著差异,3个主发生代之间的雌雄比同样无显著差异。最大雌雄比出现在2010年的第3代,达到了1.594 2,多数年份的雌雄比大于1(表2)。总体而言,白背飞虱灯下种群中,雌虫数量略占优势,这对迁入种群的繁殖也有一定的促进作用。
表22008、2009、2010、2012年三都县白背飞虱灯诱种群雌雄比
Table2Theratiooffemaleandmaleoflight-trappopulationofWBPHin2008,2009,2010and2012inSanduCounty
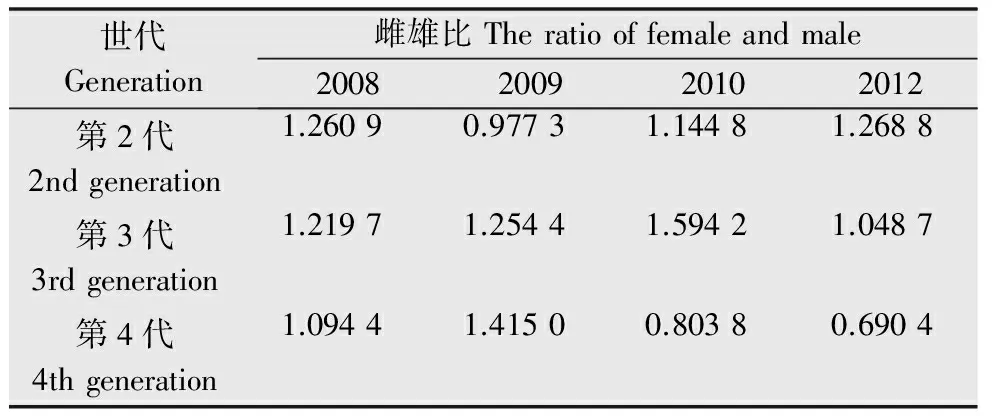
世代Generation雌雄比Theratiooffemaleandmale2008200920102012第2代2ndgeneration1.26090.97731.14481.2688第3代3rdgeneration1.21971.25441.59421.0487第4代4thgeneration1.09441.41500.80380.6904
2.2 白背飞虱田间种群
2.2.1 白背飞虱田间发生总量
2008年白背飞虱田间发生最为严重,不论系统田还是预测圃,调查期百丛总虫量(调查期内每次调查的虫量之和)都超过了30 000头;2011年田间发生最轻,调查期百丛虫量略超10 000头;从2008年至2011年,白背飞虱的田间发生数量逐年减少,但在2012年其田间发生量又出现了反弹上升的趋势;系统田的调查期百丛总虫量相对于预测圃有一定减少,说明施药防治对减轻白背飞虱的种群数量有一定作用,在防效最好的2009年,系统田的白背飞虱数量比预测圃减少了将近1/3,而在2011年,系统田的白背飞虱数量反而略微超过了预测圃。

图2 2008-2012年三都县白背飞虱百丛总虫量
2.2.2 白背飞虱卵的发生数量动态
在2010年进行了白背飞虱的田间卵量调查,由图3可以看出:系统田与预测圃的卵量均在7月上旬(7月5日)达到高峰,具有较高的同步性,最高卵量分别达到了百丛4 305粒和百丛5 400粒,系统田的百丛有效卵量明显低于预测圃,7月中旬以后,这种差异逐渐减小,两者的百丛有效卵量基本持平;带卵稻株率同样在7月5日达到最高,系统田和预测圃差异不大,但在高峰之前和之后,预测圃的带卵稻株率明显高于系统田,在整个调查期内,系统田的带卵稻株率都低于预测圃。
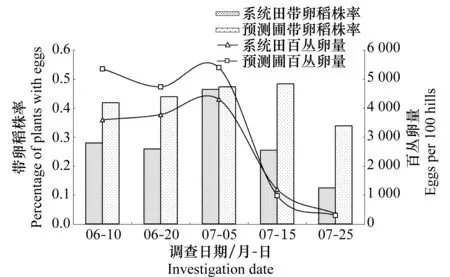
图3 2010年三都县白背飞虱田间卵量和带卵稻株率
2.2.3 白背飞虱若虫的田间发生动态
系统田中,2008年和2009年白背飞虱若虫在6月10日达到数量高峰,百丛虫量分别为13 085头和3 921头;2010和2012年白背飞虱若虫的数量高峰出现在6月20日,达到数量高峰,百丛虫量依次为2 712头和2 468头;而2011年系统田的若虫高峰出现最晚,为6月25日(百丛1 525头)。可以看出,2008年系统田的若虫发生最为严重,然后逐年减少,直至2012年系统田白背飞虱若虫发生量又有了上升(图4)。
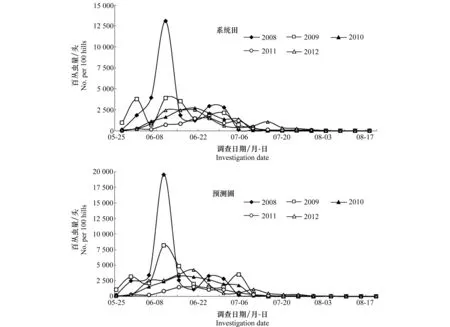
图4 三都县2008-2012年白背飞虱若虫田间发生动态
预测圃中,2008年和2009年白背飞虱若虫同样在6月10日达到数量高峰,为百丛19 541头和8 170头,显著大于系统田的百丛虫量;2010年若虫高峰在6月15日(百丛3 248头)到来,较系统田有所提前;2011年和2012年若虫高峰均出现在6月20日,百丛虫量分别为1 537头和4 250头(图4)。
由此可以看出,系统田和预测圃的白背飞虱若虫数量消长有较高的一致性,均在6月上中旬达到数量高峰,7月中上旬在田间终见,但2011年和2012年白背飞虱若虫的田间终见日延后,推至8月上旬;在调查期内,前期(6月中旬之前)预测圃的若虫虫量大都高于系统田,但后期也会出现预测圃若虫虫量少于系统田的情况。
2.2.4 白背飞虱成虫的田间发生动态
系统田中,2008年有2次明显的成虫高峰,分别是5月25日的百丛860头和6月30日的百丛1 338头;2009年成虫数量在7月10日达到最高,为百丛491头;2010年5月30日的成虫数量最多,为百丛1 505头;2012年成虫发生最高峰在5月30日(百丛591头);2011年成虫数量则在6月25日(百丛750头)达到高峰,超过了2009 年和2012年的田间成虫高峰,另外,在7月下旬至8月上旬,相对于其他年份,成虫数量还维持在了较高的500头级别,出现这种情况,可能是因为2011年白背飞虱迁入较晚的缘故:2011年白背飞虱迁入高峰集中在第4代(6月21日-7月20日),而田间虫量高峰(6月25日)也出现在这一时期,可能是直接迁入的白背飞虱形成了田间高峰;迁入的成虫基数加上其在田间增殖羽化的成虫数量可能是维持7月下旬至8月上旬较高虫量的原因。
预测圃中,2008年也有两次明显的虫量高峰,同样是5月25日(百丛1 220头)和6月30日(百丛916头);2009年预测圃的成虫数量在6月20日达到高峰(百丛654头),比系统田大为提前;2010年5月30日预测圃的成虫数量最多,为每百丛1 463头;2011年预测圃的虫量高峰出现在8月5日(百丛557头),比系统田大大推迟,此外,在7月下旬至8月上旬,预测圃成虫数量也维持在了较高的500头级别,与系统田保持一致;2012年预测圃成虫发生最高峰在5月25日,虫量为百丛715头(图5)。

图5 三都县2008-2012年白背飞虱成虫田间发生动态
与若虫相比,成虫的活动迁移能力和避害能力都大为增强,导致了系统田和预测圃的成虫数量关系更加复杂。随年份不同,差异也较大:2008年5月下旬至6月中旬,预测圃的虫量始终高于系统田,6月30日以后,系统田的虫量却略高于预测圃,但两者的虫量消长均保持着一致性;2009年两者的虫量则在7月上中旬保持一致消长,6月25日和7月5日的系统田的调查虫量略高于预测圃;2010年预测圃和系统田的虫量具有高度的消长一致性,预测圃的虫量略多,但两者十分接近;2011年预测圃和系统田的成虫数量消长也表现出良好的一致性,但是在7月中旬后,预测圃的成虫数量反而略少于系统田;2012年6月中旬以前,系统田的虫量消长变化略快于预测圃,但从6月下旬开始,两者又基本回到了一致的消长节奏。
综合上述5年的白背飞虱田间虫情表明:白背飞虱的田间种群构成中,若虫数量占有绝对优势,若虫在田间的发生多表现为单峰型,即在6月上中旬出现一次高峰,此后数量逐渐减少,末见期多集中在7月下旬,但是2011年和2012年的若虫末见期则有所推迟,延后至8月上旬,同时发生高峰期也相应延后至6月下旬,可以推测,在未来年份的白背飞虱田间种群中,其若虫发生高峰将推迟,发生期也会相应延长。
相对于若虫,白背飞虱成虫的田间种群略显微小,但是其发生期持续时间长,持续为害水稻;田间发生表现为前期大高峰为主,后期小高峰为辅的特点,一般在5月下旬至6月上旬出现一次大高峰,此后在6月下旬至7月中旬出现几次小高峰;成虫的田间发生高峰同样有向后延迟的趋势。
由于设置了预测圃和系统田,化学施药对白背飞虱的防治作用也可见一斑,通过5年的数据可以看出:化学农药对白背飞虱卵的防效最为显著,对若虫的控制作用要明显高于成虫,而且在水稻生长前期的防控作用要明显好于水稻生长中后期。
2.3 白背飞虱灯诱种群与田间种群的关系
通过系统调查数据可以看出,黔南地区白背飞虱在大田可发生3代(第3代~第5代),主害代为第3代和第4代,对5年来(2008-2012年)4月至8月各旬的灯诱虫量和田间发生世代的百丛虫量(包含若虫和成虫)进行相关性分析,得出以下结果:田间第3代的日平均百丛虫量和高峰日百丛虫量均与6月中旬、6月下旬的灯诱虫量呈显著正相关(表3);田间第4代的日平均百丛虫量和高峰日百丛虫量同样与6月中旬、6月下旬的灯诱虫量呈显著正相关(表3);第5代的田间虫量与各旬灯下虫量均未表现出相关性。
表3白背飞虱灯诱虫量与不同世代田间发生量关系
Table3CorrelationbetweenlighttrapcatchesandfieldoccurrenceofdifferentgenerationsofWBPH

项目Item各世代日均百丛虫量MeandailyamountofWBPHper100hillsofeachgenerationduringinvestigation第3代3rdgeneration第4代4thgeneration第5代5thgeneration各世代高峰日百丛虫量PeakdayamountofWBPHper100hillsofeachgenerationduringinvestigation第3代3rdgeneration第4代4thgeneration第5代5thgeneration6月中旬灯诱虫量LighttrapcatchesinmidJuner=0.905∗r=0.960∗∗无r=0.983∗∗r=0.972∗∗无6月下旬灯诱虫量LighttrapcatchesinlateJuner=0.889∗r=0.956∗∗无r=0.973∗∗r=0.975∗∗无
6月中下旬正值白背飞虱灯下第3、4代的迁入期,这在一定程度上说明田间第3、4代的发生量受灯下迁入虫源的影响较大,同时这也间接佐证了白背飞虱灯下第3、4代主要由迁入个体所构成,而第5代的田间种群受灯下种群影响较小,主要是由本地繁殖的个体构成。
3 讨论
20世纪60年代,白背飞虱仅在我国局部地区发生为害,时至今日,在全国大部分稻区普遍发生[11-13],而云南、广西、贵州三省构成的西南三角区域作为白背飞虱迁入我国的第一站,不仅是其发生为害的重大灾区,也是其继续北上为害的虫源基地。由于更加靠近西南边境,云南、广西两省白背飞虱的种群发生、虫源性质等问题近年来均有较多报道[14-19],但近年来在贵州省却相对少见。
20世纪末的研究表明,白背飞虱从3月上中旬便迁入贵州南部,5月下旬至6月上旬是其迁入的高峰期[20],而近5年的灯下数据中,只有2010年白背飞虱的灯下始见日出现在3月,其他年份均出现在4月,同时迁入高峰也集中出现在5月中旬,这说明近年来白背飞虱的始迁入期较20世纪末有所推迟,迁入高峰却有略微提前,对贵州江口县褐飞虱的研究也表明其灯下高峰期有提前的趋势[21],这可能与低空急流的时空分布变化和气候条件变迁有关[22-23]。田间方面,前人的研究认为,白背飞虱在6月中下旬出现为害高峰[20],近年的数据却证实其田间高峰在5月下旬至6月上旬出现,田间发生高峰的提前可能与灯下高峰的提前也有一定联系。
此外,本研究认为,农户大田化学防治对若虫的控制作用要明显高于成虫,而且在水稻生长前期的防控作用要明显好于水稻生长中后期。这与喷施农药的种类不无关系:用于化学防治之一的噻嗪酮的作用机理是抑制昆虫表皮几丁质的合成,从而导致稻飞虱若虫在蜕皮过程中死亡,但对成虫无直接杀死作用[24-25],田间防治试验的结果也表明噻嗪酮在稻飞虱若虫高峰期施药效果较好[26],并且在水稻生长前期,田间白背飞虱若虫较多,后期则以成虫为主,这可能是造成上述结论的原因。
稻飞虱的世代重叠现象十分明显,加上灯下迁入种群的影响,使田间种群的虫态组成复杂,大大增加了防治的难度,以后亟须结合白背飞虱虫源区域的水稻种植制度更改、水热条件变化、高空气象异动等多方面因素进行综合考虑,进一步深入探究其灯下种群与田间种群间的复杂关系,进而对其预测与防治做出有效指导。由于我国独特的地理气候条件和地面资源配置状况,国内各大稻区白背飞虱的发生规律也有相当的差异[12],明确各不同稻区的白背飞虱种群动态是当务之急,此外,走出国门加强与虫源地国家的合作与交流也是将来稻飞虱研究的必由之路。
[1]贵州省统计局, 国家统计局贵州调查总队. 贵州统计年鉴-2012[M]. 北京: 中国统计出版社, 2012.
[2]沈君辉, 尚金梅, 刘光杰. 中国的白背飞虱研究概况[J]. 中国水稻科学,2003,17(S1): 7-22.
[3]周国辉, 张曙光, 邹寿发, 等. 水稻新病害-南方水稻黑条矮缩病发生特点及为害趋势分析[J]. 植物保护,2010,36(2): 235-237.
[4]全国白背飞虱科研协作组. 白背飞虱迁飞规律的初步研究[J]. 中国农业科学,1981(5): 25-31.
[5]四川、贵州、云南三省稻飞虱科研协作组. 我国西南稻区白背飞虱, 褐稻虱的迁飞和发生特点[J]. 植物保护学报,1982,9(3): 179-186.
[6]Kisimoto R, Rosenberg L J. Long-distance migration in delphacid planthopper[M]∥Denno R F, Perfect T J, eds. Planthopper: their ecology and management. New York: Chapman and Hall,1994.
[7]Otuka A, Matsumura M, Sanada-Morimura S, et al. The 2008 overseas mass migration of the small brown planthopper,Laodelphaxstriatellus(Fallen), and subsequent outbreak of rice stripe disease in western Japan[J]. Applied Entomology and Zoology,2010,45(2): 259-266.
[8]Syobu S, Otuka A, Matsumura M. Trap catches of the small brown planthopper,Laodelphaxstriatellus(Fallen) (Hemiptera: Delphacidae), in northern Kyushu district, Japan in relation to weather conditions[J]. Applied Entomology and Zoology,2011,46(1): 41-50.
[9]翟保平, 张孝羲. 迁飞过程中昆虫的行为: 对风温场的适应与选择[J]. 生态学报, 1993,13(4): 356-363.
[10]贵州省质量技术监督局. 贵州省稻飞虱监测技术规范[EB/OL]. (DB52/T 396-2005)[2005-07-01]. http:∥www.gzqts.gov.cn/cszz/bzhc/gzsdfbzzdxdyjzx/200601/4211.shtml.
[11]秦厚国, 叶正襄, 舒畅, 等. 白背飞虱种群治理理论与实践[M]. 南昌: 江西科学技术出版社, 2003.
[12]翟保平. 稻飞虱: 国际视野下的中国问题[J]. 应用昆虫学报,2011,48(5): 1184-1193.
[13]翟保平, 周国辉, 陶小荣, 等. 稻飞虱暴发与南方水稻黑条矮缩病流行的宏观规律和微观机制[J]. 应用昆虫学报,2011,48(3): 480-487.
[14]吕芬, 周平, 黄新动, 等. 白背飞虱灯下虫量与气象要素关系的相关分析[J]. 应用昆虫学报,2007,44(1): 48-53.
[15]沈慧梅, 吕建平, 周金玉, 等. 2009年云南省白背飞虱早期迁入种群的虫源地范围与降落机制[J]. 生态学报,2011,31(15): 4350-4364.
[16]蒋春先, 韦加贵, 张云慧, 等. 云南富宁县白背飞虱早期迁入虫源地及降落机制分析[J]. 四川农业大学学报,2012,30(2): 232-237.
[17]凌炎, 范桂霞, 龙丽萍. 广西不同白背飞虱种群对杀虫剂的敏感性测定[J]. 广西农业科学,2009,40(7): 847-849.
[18]沈慧梅, 陈晓, 胡高, 等. 2008 年广西北部湾稻区稻飞虱初迁入过程分析[J]. 应用昆虫学报, 2011,48(5): 1268-1277.
[19]武俊杰, 蒋春先, 张云慧, 等. 2011年广西兴安地区白背飞虱种群发生动态及迁飞轨迹分析[J]. 植物保护,2012,38(5): 51-57.
[20]金星, 王德其, 金道超, 等. 贵州省稻飞虱发生危害规律与防治对策[J]. 山地农业生物学报,1998,17(4): 208-214.
[21]闫香慧, 刘怀, 赵志模, 等. 水稻褐飞虱灯下发生期及种群数量动态分析[J]. 植物保护学报,2008,35(6): 501-506.
[22]肖满开, 何木兰, 余夕辉, 等. 安庆褐飞虱近三个大发生年虫源和气候条件的比较分析[J]. 应用昆虫学报,2008,45(1): 46-50.
[23]包云轩, 谢杰, 向勇,等. 低空急流对中国褐飞虱重大北迁过程的影响[J]. 生态学报,2009,29(11): 5773-5782.
[24]Izawa Y, Uchida M, Sugimoto T, et al. Inhibition of chitin biosynthesis by buprofezin analogs in relation to their activity controllingNilaparvatalugensStål[J]. Pesticide Biochemistry and Physiology,1985,24(3): 343-347.
[25]Uchida M, Sugimoto T, Asai T. Inhibition of cuticle deposition and chitin biosynthesis by a new insect growth regular, buprofezin inNilaparvatalugensStål[J]. Agricultural and Biological Chemistry,1987,49(4): 1233-1234.
[26]孙俊铭, 韦刚, 王皖伟, 等. 毒死蜱与噻嗪酮、吡虫啉混用防治稻飞虱和二化螟田间试验[J]. 应用昆虫学报,2004,41(6): 541-544.
Populationdynamicsofwhite-backedplanthopper,Sogatellafurcifera(Horváth)inricefieldofsouthGuizhou,China
Xue Wenpeng, Jin Daochao, Yang Hong
(GuizhouKeyLaboratoryforAgriculturalPestManagementofMountainousRegion,InstituteofEntomology,GuizhouUniversity,Guiyang550025,China)
Light trapping and systematic field surveys ofSogatellafurcifera(Horváth) (white-backed planthopper, WBPH) were conducted in Sandu County in southern Guizhou from 2008 to 2012. Populations dynamic of WBPH both under lamp and in field were analyzed. The results showed that immigration of WBPH began in mid-to-late April and the peak of immigration appeared from middle-to-late May. Main peaking dates of light-trap catches were significantly correlated with first appearance date (r=0.996**) and initial outbreak date (r=0.997**). The number of female was slightly higher than that of male, and there was no significant difference between years for the ratio of female and male. Nymphs which were the main components occurred shorter period than adults in field populations, and the number of nymphs arrived peak in early June and ended in mid-to-late July. Adults which occurred over whole growth period of rice achieved peak from late May to early June. The amount of the field main damage generation showed a significant positive correlation with the light trap catches in mid to late June.
Sogatellafurcifera; light-trap population; field population; southern Guizhou
2013-09-24
:2013-12-30
贵州省教育厅自然科学研究项目(黔教2010011);贵州省农业科技攻关项目(黔科合NY字[2013]3006,黔科合NY字[2010]3064);国家“973”计划前期研究专项(2009CB125908)
S 435.112
:ADOI:10.3969/j.issn.0529-1542.2014.04.025
* 通信作者 E-mail: agr.hyang@gzu.edu.cn
