湖北神农架川金丝猴食物营养的季节性特征1)
2014-07-27李进宇张志翔杨敬元
李进宇 张 晶 张志翔 杨 占 杨敬元
(北京林业大学,北京,100083) (辽宁省林业调查规划院) (湖北神农架自然保护区)
湖北神农架川金丝猴食物营养的季节性特征1)
李进宇 张 晶 张志翔 杨 占 杨敬元
(北京林业大学,北京,100083) (辽宁省林业调查规划院) (湖北神农架自然保护区)
探讨了川金丝猴(Rhinopithecusroxellana)对食物选择与食物营养成分间的关系,分析了不同季节川金丝猴在营养摄取上的响应策略。在湖北神农架千家坪地区设置临时观测点,采集不同季节川金丝猴的食源植物,测定和分析其粗蛋白、粗脂肪、酸性洗涤纤维以及灰分质量分数。结果表明:川金丝猴的食源植物中粗蛋白的含量在春季最高,其后呈现下降趋势;粗纤维的含量比较稳定,在春、夏、秋3季变化幅度较小,仅在春季含量低于粗蛋白,在其他季节均高于其余几种营养成分。川金丝猴倾向于选择高蛋白质和低纤维素的植物作为食物。食源植物和食物的充分与否呈现季节性变化的规律,使得川金丝猴在营养摄入方面具有季节性特征。
川金丝猴;食源选择;营养成分;粗蛋白;蛋白纤维比
Journal of Northeast Forestry University.-2014,42(7).-148~153
We analyzed the macronutrient data collected separately in spring, summer and autumn over three years from leaves and fruits of plant species available as food for the snub-nosed monkey in Shennongjia National Nature Reserve, China. We measured the nutritional components including crude protein (CP), crude lipid (CL), acid detergent fiber (ADF), ash (ASH), and protein-to-fiber ratio (PFR). The food choice ofRhinopithecusroxellanaacross the year does fit the protein to fiber model, but had a seasonal shift in response to the change of diet macronutrients. The protein content in the diet food decreased from spring to autumn, but the fiber amount almost was a fixed level. Therefore, though fruits join in the diets of the monkeys in summer and autumn, there is still a tendency of the diet nutrients of higher PFR in spring than in summer and autumn. The food choice ofR.roxellanais in response to the nutrient chemistry of a plant species in a certain season.
Keywords Snub-nosed monkey (Rhinopithecusroxellana); Macronutrient; Crude protein; Protein-to-fiber ratio
在保护生物学研究中,对影响濒危以及受威胁物种种群丰富度的研究变得越来越迫切[1]。而影响动物自身发展的因素较多,其中营养的摄入便是影响物种丰富度的主要因素之一。作为动物种群的主体,动物个体所需的营养物质,如蛋白质、脂类、碳水化合物、维生素和微量元素等,它们是能确保动物自身新陈代谢等功能正常运转的基本物质基础[2-6],同时进一步促进了动物种群的稳定和繁荣。自然界中的灵长类绝大多数以植物为食,因此对于灵长类的种群稳定以及延续,灵长类如何保证能够获取足够的食物,来满足其自身营养需求就显得十分重要,而相关的灵长类取食策略或者是营养选择方式的研究在灵长类保护中起着突出的作用和意义[7-11]。疣猴类作为灵长类动物的一个重要组成部分,最显著的特点就是增大的囊状前胃,有大量的微生物来发酵摄入的食物,可以更快的吸收营养[12]203-229,[13]189-202,[14-17],而这种特性与反刍动物相似[13]189-202,因此,疣猴可以通过取食大量的纤维素食物来满足自身营养的需求[18-20]。蛋白纤维比的提出对于那些不确定是否有特定食源植物的食叶灵长类是一个很好的预测模型。这个预测模型在鬃毛吼猴(Alouattapalliata)[20-21]、栗红叶猴(Presbytisrubicunda)[22]、黑疣猴(Colobussatanas)[23]、长鼻猴(Nasalislarvatus)[24]、低地大猩猩(Gorillagorilla)[25]和其他几种狐猴[26]中都得到了科学的验证。同时该模型也可以用来解释那些饮食中有明确偏好的一些灵长类物种,例如:西方红疣猴(Procolobusbadius)[27]、黑白疣猴(Colobusguereza)[28-29]以及狒狒属(Papiospp.)[30]。川金丝猴是中国特有的一种生活在温带地区的灵长类,属疣猴亚科,主要分布于四川、陕西、甘肃、湖北以及重庆[31-33],[34]79-89,[35]等地,一般生活在海拔1 500~4 100 m的针阔混交林或阔叶混交林内。根据已有研究数据表明,川金丝猴的食性主要为叶食性,但是在食物组成、质量和数量上都有较为明显的季节性变化趋势。川金丝猴的取食习惯偏向叶食性灵长类,以芽、叶片、果实、树皮为主,以松萝作为补充[34]126-133,[36-37]。迄今,关于川金丝猴以及仰鼻猴属的研究大多集中在动物行为以及食源植物等方面,而对于其摄入营养的研究较少。本研究选择川金丝猴分布区之一的湖北神农架国家自然保护区,通过设置半固定样点,对川金丝猴不同季节食物选择和营养成分间的关联做了研究,分析不同季节川金丝猴在营养摄取上的响应策略,进一步为该物种提供有效的保护和管理策略信息。
1 研究区概况
研究区设在湖北神农架自然保护区的千家坪地区(31°22′~31°37′N,110°03′~110°34′E),该区面积约40 km2,主要以温带阔叶落叶混交林和针阔混交林为主,海拔1 500~2 650 m。属于亚热带湿润,季风性气候。该地区春季从4月初到5月末,夏季是6月到8月中旬,秋季为8月下旬到10月,接着便是漫长的冬季,从11月到第2年的3月底。该地区平均海拔为1 700 m,年降水量为1 800 mm,7月份平均温度为17.8 ℃,1月份平均温度-2.8 ℃。从12月中旬开始到第2年的3月中旬为冰雪覆盖期[33]。在海拔1 700 m以上的植被主要可以划分为两大类型:温带落叶阔叶针叶混交林(1 700~2 600 m)和温带常绿针叶林(>2 600 m)。
2 研究方法
样品采集:通过调查统计已有的川金丝猴食源植物数据以及文献资料[32-33],[34]126-133,[35],选择了约40种具有代表性的食源植物作为取样主体(主要标准就是主要植被的优势物种及大量分布物种,不包含特殊的分布较少的物种),及部分非食源植物来进行营养成分分析(表1)。从2010年到2012年,按照季节(春季、夏季、秋季,冬季由于大雪封山以及落叶的原因没有选择)来搜集目标植物样品。春季主要以嫩叶、嫩芽为主,夏、秋季主要是成熟叶片以及部分果实。在千家坪不同海拔地区设立固定样品采集点(植株)并标记,在每个季节中尽量选择固定的目标树种作为样品采集的对象,如果出现该植株死亡或者其他原因,则选择其周围的相同树种替代。采集样品的位置为植株树冠的中部,这是因为川金丝猴活动和取食的部位主要集中在树冠的中层;将采集下来的叶片立即放入密封塑料袋,带回基地后,于烘干箱内,60 ℃烘干;将干样品放入新的干燥的封口塑料袋中保存,编号,记录。

表1 神农架川金丝猴食源植物及非食源植物的营养成分
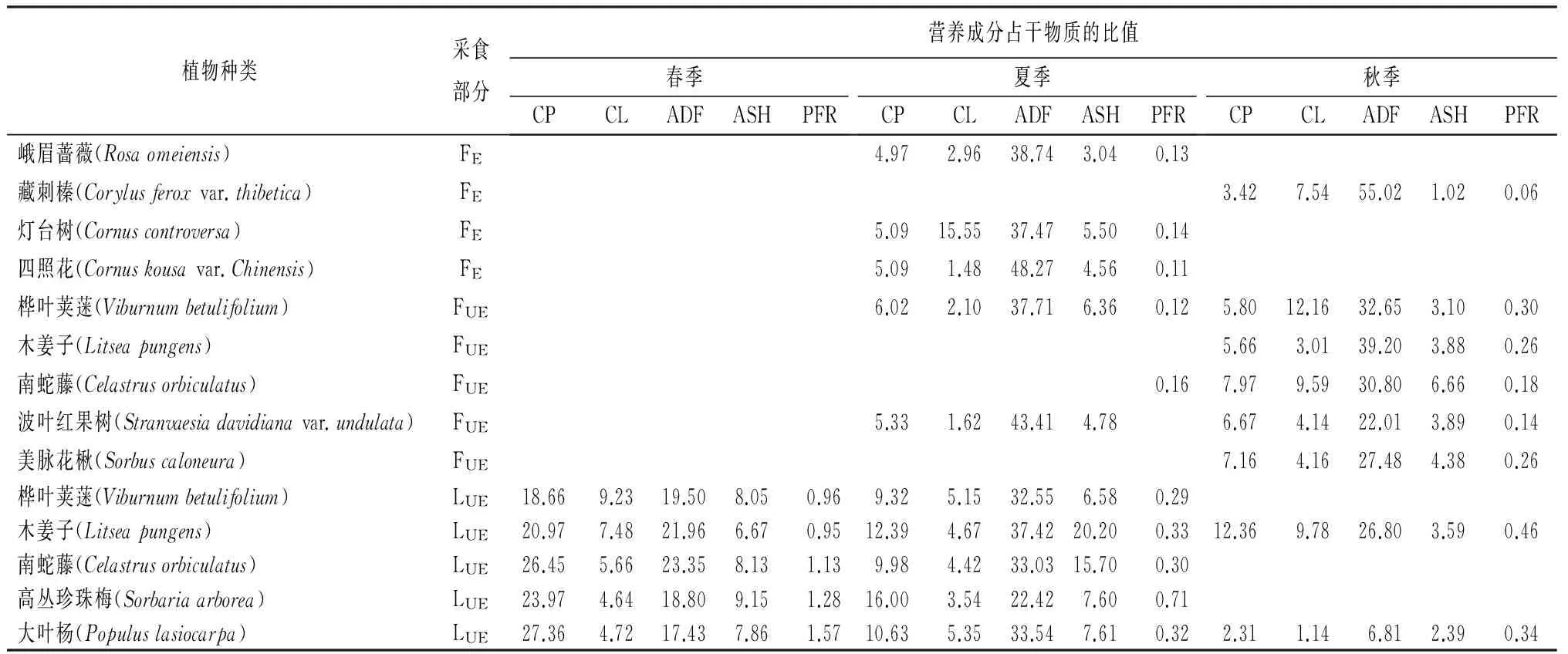
续(表1)
注:L.叶;F.果实或种子;All.整株;E.食源植物;UE.非食源植物;CP.粗蛋白;CL.粗脂肪;ADF.酸性洗涤纤维;ASH.灰分;PFR.蛋白纤维比。
营养成分测定:采集的样品在中国农业科学院饲料研究所进行检测分析,检测成分包括粗蛋白、粗脂肪、酸性洗涤纤维以及灰分。
数据分析:用SPSS 19.0进行数据分析。按照3个季节(春季、夏季、秋季)来分析数据,用单因素方差分析来分析不同的营养元素的季节性差异,使用T检验来比较每个季节里食源植物和非食源植物营养成分的差异,同时利用Spearman检验不同营养成分与季节间的关联性。
3 结果与分析
3.1 不同季节食源植物营养的比较
通过对2010年到2012年间春季、夏季、秋季的食源植物样品的检测,得到了3个季节中样品的粗蛋白、粗脂肪、粗纤维以及灰分的干物质质量分数(表1)。粗蛋白的均值在春季里质量分数超过了22%,是3个季节中最高的,超过夏季粗蛋白质量分数(9.91%±3.19%,n=26)的2倍,是秋季粗蛋白(7.6%±2.4%,n=20)的3倍多(表2)。粗蛋白含量最高的物种是西南卫矛(Euonymushamiltonianus),约为29.75%,此外在垂丝丁香(Syringakomarowiivar.reflexa)、卫矛(Euonymusalatus)、多枝柳(Salixpolyclona)和米心水青冈(Fagusengleriana)中含量也比较高(表1)。
粗脂肪在3个季节都低于7%。其中秋季粗脂肪质量分数最高(6.92%±11.26%,n=16),约为春季(3.74%±1.15%)和夏季(3.88%±3.60%)的2倍(表2),主要原因是秋季食物中有大量的种子和果实。在所有样品中,粗脂肪质量分数最高的是华山松的种子(54.89%),而其也是川金丝猴在该地区秋季的主要食物来源之一(表1)。3个季节的粗纤维含量均超过了20%,春季的粗纤维含量最低(22.13%±6.86%,n=15),夏季的粗纤维含量最高(32.04%±7.82%,n=26)。在春季,粗纤维的摄入量最低,但是粗蛋白的摄入为三季最高(表2)。灰分在3个季节里都低于7%。在夏季,灰分的均值最高,达到了8.22%;秋季最低,约为5.10%;春季居于夏秋之间,约为7.22%(表2)。蛋白纤维比的最高点出现在春季(1.15%±0.48%,n=15),夏季(0.33)和秋季(0.35)几乎相同(表2)。
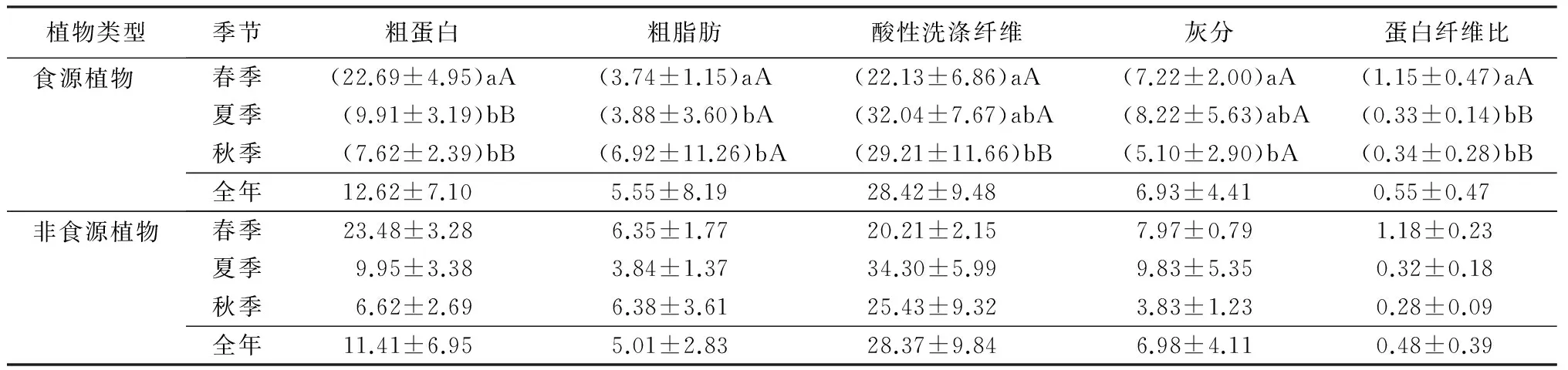
表2 神农架川金丝猴食源植物及非食源植物营养成分均值以及食源植物季节间多重比较
注:表中数据为平均值±标准差,3个季节中食源植物营养成分数据后的不同小写字母表示差异显著(P<0.05),大写字母表示差异极显著(P<0.01)。
通过对比不同季节食源植物营养成分发现粗蛋白(F=66.256,P<0.01)、粗纤维(F=4.154,P<0.05)和蛋白纤维比(F=26.144,P<0.01)在春、夏、秋3个季节内呈显著差异,粗脂肪(F=1.754,P>0.05)和灰分(F=2.804,P>0.05)无显著差异。分析几种营养元素之间的关联性,发现粗蛋白和灰分(rs=0.528,P<0.01,n=57),粗蛋白和蛋白纤维比(rs=0.831,P<0.01,n=57)之间存在显著正相关;粗蛋白和酸性洗涤纤维存在显著负相关(rs=-0.328,P<0.05,n=57);粗脂肪和其他成分间无关联性;粗纤维与蛋白纤维比存在负相关(rs=-0.720,P<0.01,n=57),与灰分无关联(表3)。

表3 不同营养成分的关联性
注:*为P<0.05;** 为P<0.01。
3.2 食源植物和非食源植物间营养的差异
根据3个季节里食源植物与非食源植物营养成分的比较,发现在春季里,粗脂肪质量分数存在显著差异(t=3.418,P<0.01),但粗蛋白、粗纤维、灰分以及蛋白纤维比无差异;夏季的情况也和春季类似,粗脂肪在夏季存在差异(t=2.597,P<0.05);秋季里,食源植物与非食源植物间无显著差异(表2、表4)。
表4 不同季节食源植物与非食源植物营养成分的T检验
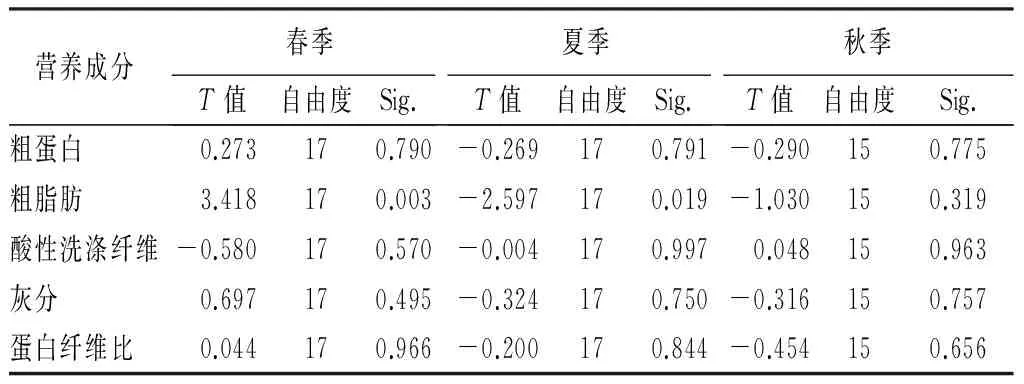
营养成分春季T值自由度Sig.夏季T值自由度Sig.秋季T值自由度Sig.粗蛋白0.273170.790-0.269170.791-0.290150.775粗脂肪3.418170.003-2.597170.019-1.030150.319酸性洗涤纤维-0.580170.570-0.004170.9970.048150.963灰分0.697170.495-0.324170.750-0.316150.757蛋白纤维比0.044170.966-0.200170.844-0.454150.656
4 结论与讨论
在温带及寒温带地区,由于低温、降水少而且海拔高,每年的冬季都会持续较长时间,在这段时间内,由于食物数量的减少而导致食物来源匮乏,因此动物缺乏足够的食物(或营养)来维持日常生活需求,从而使得动物种群的多样性降低。灵长类动物一般分布在热带以及亚热带地区,在温带区域分布的种类极少[38-40]。川金丝猴是少数的生活在温带地区的叶食性灵长类,主要以该区域的阔叶混交林和针阔混交林为主要栖息场所,川金丝猴主要以植物的叶片、嫩枝等为主要的食物,在夏季和秋季也食用一些植物的果实(或种子)[34]127-130。川金丝猴的生活季节大致可以划分为食物匮乏期和食物丰富期[34]127-130。
蛋白质的摄入[41]是疣猴在选择食物过程中的主要影响因子。疣猴在其生长、发育和繁殖的阶段都会倾向选择富含蛋白质的食物[11,17,20,42-43]。Milton[17]的研究认为蛋白纤维比可以很好地揭示吼猴选择食物的倾向性,进而认为在一些小型的植食性哺乳动物中,蛋白纤维比亦能解释这些动物在食物选择中的取舍和策略[23]。其他的一些灵长类专家通过对不同灵长类营养策略的研究也认为蛋白纤维比在各自研究的对象中可以很好的解释取食的内在原因。这就表明,灵长类在选择食物的过程中很大程度上取决于食物中蛋白质和纤维的含量,即蛋白纤维比在食物选择过程中占据重要的决定性作用。
通过研究发现,神农架地区的川金丝猴其食源植物中粗纤维的含量比较稳定,在3个季节里变化幅度较小,仅在春季里含量低于粗蛋白,在其他的季节里都要高于其他几种营养成分。粗蛋白的平均含量为12.62%,与现有生活在温带地区的其他灵长类相似,如日本猴(Macacafuscata)10.3%~12.1%[12]203-229、滇金丝猴(Rhinopithecusbieti)11.01%[38]482-495,而且明显高于亚热带地区的灵长类,如黑叶猴在冬季蛋白摄入的值为5.4%,秋季则为4.2%[40-41]。但是,神农架地区的川金丝猴粗纤维摄取量约为28.42%,接近于其他温带地区灵长类的最低值,如日本猴42.2%~42.4%[11]、滇金丝猴21.3%[38]482-495、白头叶猴(Presbytisleucocephalus)31.1%[40]和30.8%[41]。因此,通过对同一地域的几种灵长类之间营养成分摄取的比较,川金丝猴倾向于选择那些含较多蛋白质和较少纤维素的植物作为食物来源。这个结果比较符合Milton[17]的理论——灵长类动物一般选择那些蛋白纤维比较高的食物,换言之就是灵长类在选择食物时,尽量保证最大化摄入蛋白质,同时减少纤维素的摄入,也就是蛋白纤维比高的食物。这种假说同样在其他非人类灵长类物种中得到验证,比如黑白疣猴(Colobusguereza)[1,29]、东部大猩猩(Gorillaberingei)[42]、西方红疣猴(Procolobusbadius)[1]以及滇金丝猴[14]。
此外,研究结果还显示川金丝猴在营养摄入上存在季节性的变化,虽然整体上食物选择符合蛋白纤维比高的食物。但在春季里,蛋白纤维比值大于1,而夏季和秋季里,这个比值小于1,说明在夏季和秋季里,川金丝猴的营养摄入存在一个蛋白质量下降和纤维量上升的过程,由于纤维的含量较稳定,川金丝猴在夏秋季减少了蛋白质的摄入。也就是说,川金丝猴在资源匮乏时期倾向于摄入营养的蛋白最大化,而在食物丰富的时期,蛋白就没有那么明显和重要。出现这种情况主要是因为:首先在春季里,川金丝猴主要以嫩叶和嫩芽为食,相对于成熟叶片来说,嫩叶和嫩芽纤维含量较低,而且富含蛋白质;其次,在夏末和秋季,大量的果实和种子的取食,导致了碳水化合物以及脂肪摄入的增加,同时也减少了蛋白质的摄入。最后,不同植物次生代谢产物的作用以及栖息地的环境条件也会影响川金丝猴的食物选择,进而影响营养元素的摄入。这种营养摄入的转换,在Agilewallaby中也有类似的结论,在干旱季节里,Agilewallaby热衷于选择低蛋白和高纤维的食物[43]。
总的来说,神农架地区川金丝猴的食性选择符合和支持了Milton[17]的蛋白纤维比的指标,偏好高蛋白和低纤维的食物。但是在考虑单个季节来说,由于每个季节的食物种类变化不一,因此川金丝猴每年都会遭遇食物缺乏期——食物充足期——食物缺乏期的食物供应季节性特征,这样的变化造成的结果就是川金丝猴食源植物和食物充分与否呈现季节性变化规律,导致了川金丝猴在营养方面的摄入产生季节性变化。
[1] Wasserman M D, Chapman C A. Determinants of colobine monkey abundance: the importance of food energy, protein and fibre content[J]. Journal of Animal Ecology,2003,72(4):650-659.
[2] Milton K. The foraging strategy of howler monkeys: a study in primate economics[M]. New York: Columbia University Press,1980.
[3] Boutin S. Food supplementation experiments with terrestrial vertebrates: patterns, problems and the future[J]. Canadian Journal of Zoology,1990,68(2):203-220.
[4] Behie A M, Pavelka M S. The role of minerals in food selection in a black howler monkey (Alouattapigra) population in Belize following a major Hurricane[J]. American Journal of Primatology,2012,74(11):1054-1063.
[5] Casimir M J. Feeding ecology and nutrition of an eastern gorilla group in the Mt. Kahuzi region (République du Zaïre)[J]. Folia Primatologica,1975,24(2/3):81-136.
[6] Chivers J D. The feeding of the siamang (Symphatangussiamang)[M]//Clutton-Brock T H. Primate ecology. London: Academic press,1977.
[7] Hladik M C. Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes[M]. London: Academic press,1977.
[8] Smith C C. Feeding behaviour and social organization of howling monkeys[M]//Clutton-Brock T H. Primate ecology. London: Academic press,1977.
[9] Oates J W, Waterman P G, Choo G. Food selection by the south Indian leaf-monkey,Presbytisjohnii, in relation to leaf chemistry[J]. Oecologia,1980,45(1):45-56.
[10] Kay R N, Davies A G. Digestive physiology[M]//Davies G, Oates J. Colobine monkeys: their ecology, behaviour and evolution. Cambridge: Cambridge university press,1994.
[11] Kirkpatrick R C. The natural history and conservation of the snub-nosed monkeys (genusRhinopithecus)[J]. Biological Conservation,1995,72(3):363-369.
[12] Fashing J P. African colobines monkeys: patterns of between-group interaction[M]//Campbell C J. Primate in perspective. New York: Oxford university press,2007.
[13] Kirkpatrick C R. The Asian colobines: diversity among leaf-eating monkeys[M]//Campell C J. Primate in perspective. New York: Oxford university press,2007.
[14] Huang Zhipang, Huo Sheng, Yang Shuguo, et al. Leaf choice in black-and-white snub-nosed monkeysRhinopithecusbietiis related to the physical and chemical properties of leaves[J]. Current Zoology,2010,56(6):643-649.
[15] Struhsaker T T, Leland L. Colobines: infanticide by adult males[M]//Smuts B B, Cheney D L, Seyfarth R M, et al. Primate societies. Chicago: University of Chicago Press,1987.
[16] Kirkpatrick R C, Zou R J, Dierenfeld E S, et al. Digestion of selected foods by Yunnan snub-nosed monkeyRhinopithecusbieti(Colobinae)[J]. American Journal of Physical Anthropology,2001,114(2):156-162.
[17] Milton K. Factors influencing leaf choice by howler monkeys:a test of some hypotheses of food selection by generalist herbivores[J]. American Naturalist,1979,114(3):362-378.
[18] Milton K. Physiological ecology of howlers (Alouatta): energetic and digestive considerations and comparison with the colobinae[J]. International Journal of Primatology,1998,19(3):513-548.
[19] Davies A G, Waterman P G. Food selection by two South-east Asian colobine monkeys (PresbytisrubicundaandPresbytismelalophos) in relation to plant chemistry[J]. Biological Journal of the Linnean Society,1988,34(1):33-56.
[20] McKey D, Gartlan J S, Waterman P G, et al. Food selection by black colobus monkeys (Colobussatanas) in relationship to food chemistry[J]. Biological Journal of the Linnean Society,1981,16(2):115-146.
[21] Yeager C P, Silver S C, Dierenfeld E S. Mineral and phytochemical influences on foliage selection by the proboscis monkey (Nasalislarvatus)[J]. American Journal of Primatology,1997,41(2):117-128.
[22] Rogers M E, Maisels F, Williamson E A, et al. Gorilla diet in the lope reserve, Gabon: a nutritional analysis[J]. Oecologia,1990,84(3):326-339.
[23] Ganzhorn J U. Leaf chemistry and the biomass of folivorous primates in tropical forests[J]. Oecologia,1992,91(4):540-547.
[24] Chapman C A, Chapman L J. Foraging challenges of red colobus monkeys: influence of nutrients and secondary compounds[J]. Comparative Biochemistry and Physiology Part a, Molecular & Integrative Physiology,2002,133(3):861-875.
[25] Chapman C A, Chapman L J, Naughton-Treves L, et al. Predicting folivorous primate abundance: validation of a nutritional model[J]. American Journal of Primatology,2004,62(2):55-69.
[26] Fashing P J, Dierenfeld E S, Mowry C B. Influence of plant and soil chemistry on food selection, ranging patterns, and biomass ofColobusguerezain Kakamega Forest, Kenya[J]. International Journal of Primatology,2007,28(3):673-703.
[27] Whiten A, Byrne R W, Barton R A, et al. Dietary and foraging strategies of baboons[J]. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences,1991,334:187-195.
[28] Kirkpatrick R C, Gu H J, Zhou X P. A preliminary report on Sichuan snub-nosed monkeys (Rhinopithecusroxellana) at Baihe Nature Reserve[J]. Folia Primatologica,1999,70(2):117-120.
[29] Li Yiming. The seasonal diet of the Sichuan snub-nosed monkey (Pygathrixroxellana) in Shennongjia Nature Reserve, China[J]. Folia Primatologica,2001,72(1):40-43.
[30] Li Yiming, Stanford C B, Yang Yuhui. Winter feeding tree choice in Sichuan Snub-Nosed monkeys (Rhinopithecusroxellana) in Shennongjia Nature reserve, China[J]. International Journal of Primatology,2002,23(3):657-675.
[31] Li Yiming. Seasonal variation of diet and food availability in a group of Sichuan snub-nosed monkeys in Shennongjia Nature Reserve, China[J]. American Journal of Primatology,2006,68(3):217-233.
[32] 铁军.神农架川金丝猴栖息地植物构成和食源植物评价研究[D].北京:北京林业大学,2009.
[33] 胡锦矗,邓其祥,余志伟,等.大熊猫金丝猴等珍稀动物生态生物学研究[J].南充师院学报:自然科学版,1980,1(2):1-39.
[34] 全国强,谢家骅.金丝猴研究[M].上海:上海科技教育出版社,2002.
[35] Grueter C C, Li Dayong, Ren Baoping, et al. Fallback foods of temperate-living primates: a case study on Snub-Nosed monkeys[J]. American Journal of Physical Anthropology,2009,140(4):700-715.
[36] Kool K M. Food selection by the silver leaf monkey,Trachypithecusauratussondaicus, in relation to plant chemistry[J]. Oecologia,1992,90(4):527-533.
[37] Cords M. Interspecific and intraspecific variation in diet of two forest guenons,CercopithecusascaniusandC.mitis[J]. Journal of Animal Ecology,1986,55(3):811-827.
[38] Lambert J. Primate nutritional ecology: feeding biology and diet at ecological and evolutionary scales[M]//Bearder S K, Fuentes A, Mackinnon K C, et al. Primates in perspective. New York: Oxford university press,2007.
[39] Waterman P, Choo G. The effects of digestibility-reducing compounds in leaves on food selection by some Colobinae[J]. Malaysian Applied Biology,1981,10(2):147-162.
[40] 李雯,郭爱伟,王彬,等.冬季麻阳河黑叶猴的食物营养组分及其对食物选择的影响[J].兽类学报,2010,30(2):151-156.
[41] 蔡锐芳,胡刚,曹晔,等.贵州麻阳河黑叶猴秋季主要食物营养分析及对食物选择的影响[J].四川动物,2011,30(3):366-371.
[42] Rothman J M, Chapman C A, Pell A N. Fiber-bound Nitrogen in gorilla diets: implications for estimating dietary protein intake of primates[J]. American Journal of Primatology,2008,70(7):690-694.
[43] Stirrat S C. Foraging ecology of the agile wallaby (Macropusagilis) in the wet-dry tropics[J]. Wildlife Research,2002,29(4):347-361.
Macronutrient Seasonal Changing on Snub-nosed Monkey (Rhinopithecusroxellana) Diet in Shennongjia National Nature Reserve, China/
Li Jinyu, Zhang Jing, Zhang Zhixiang(Beijing Forestry University, Beijing 100083, P. R. China); Yang Zhan(Liaoning Forestry Inventory and Planning Institute); Yang Jingyuan(Management Bureau of Hubei Shennongjia National Nature Reserve)//
1) 国家林业局野生动物保护管理项目;国家林业局自然保护区研究和评价中心重点资助项目。
李进宇,男,1981年11月生,北京林业大学自然保护区学院,博士研究生。
张志翔,北京林业大学自然保护区学院,教授。E-mail:zxzhang@bjfu.edu.cn。
2013年12月19日。
S718.65
责任编辑:程 红。
