十六烷基三甲基溴化铵强化产氢发酵
2014-07-13任云利杨松波
任云利,李 欣,张 柯,杨松波
(河南科技大学化工与制药学院,河南洛阳471023)
0 引言
化石燃料是当今人类生产、生活的主要能源。随着全球能量需求的增长,化石燃料等不可再生能源将日趋枯竭。氢能因具有可再生性和对环境友好的特性,成为最理想的能源载体之一。在各种产氢方法中,厌氧发酵制氢因其能耗低且可利用的底物广泛,受到国内外学者的广泛关注。通过优化发酵条件(如发酵温度、pH和底物浓度等)实现产氢过程的强化已经做了大量的研究[1]。近几年,一些研究者尝试在培养基中添加微生物生长代谢所需要的微量化学物质(如金属离子、烟酸、乳酸和L-半胱氨酸)实现强化发酵产氢[2-5]。
各种类型微生物的产氢过程都是氢酶催化的可逆的氧化还原反应。降低氢气分压,有利于可逆反应向着产氢方向进行。一些研究者通过降低发酵容器上空的氢气分压实现了累计产氢量的提高[6-7]。事实上,微生物细胞内所产生的氢气首先要经过细胞膜释放到培养基,然后再扩散至反应器的上空。而由细胞膜的流动性所形成的孔径(不超过0.5~1.0 nm)仅仅略大于氢气分子的空气动力学直径(0.3 nm)。因此,推测出提高微生物胞外膜的通透性,有可能从根本上降低胞内的氢气分压,从而促进产氢代谢。文献[8]报道经表面活性剂处理过的红酵母细胞或者重组大肠杆菌细胞能显著提高胞外膜的通透性,最终有效提高反式肉桂酸的转化率。文献[9]研究得出将十六烷基三甲基溴化铵(CTAB)加入产气肠杆菌(一种高效产氢菌)整细胞的悬浮液中,胞内氢酶的活性可以得到检测。因此,本文尝试添加CTAB以提高产氢菌胞外膜的通透性,从而实现强化产氢的目的。
木糖是自然界中大量存在的植物纤维水解后的重要单糖之一。以木糖为碳源,研究混合菌发酵过程中CTAB对菌体胞外膜通透性、对细菌生长和对累计产氢量、产氢速率的影响。这种通过添加微量的表面活性剂以强化产氢的方法为将来厌氧发酵制氢的研究拓展了思路。
1 材料和方法
1.1 菌种和培养基
试验用的混合菌取自洛阳市生生乳业牛粪沼气发酵池。接种之前,混合菌先在100℃条件下处理
培养基组成:木糖10 g/L,蛋白冻5 g/L,磷酸二氢钾14 g/L,磷酸氢二钾6 g/L,硫酸铵2 g/L,柠檬酸钠1 g/L,七水硫酸镁0.2 g/L。
1.2 试验方法
胞外膜通透性试验:将37℃条件下厌氧培养24 h后的新鲜菌液离心、收集菌体,然后用100 mmol的磷酸盐缓冲液(pH=7)洗涤、离心、重悬后得到OD600(600 nm波长下光密度)为3.31的菌体悬浮液。取4 mL菌体悬浮液和0.2 mL、0~1.2 g/L的CTAB溶液一并装入7 mL的血清瓶,在37℃ 放置15 min、90 min或135 min。然后在8 944g条件下离心2 min,测上清液在280 nm下的吸光度(A280)。比较不同条件下A280值,分析从胞内释放出蛋白的相对量,从而比较不同条件下胞外膜的通透性。
批式发酵试验:在500 mL的发酵瓶中装入100 mL的培养基、5mL浓度为0~1.0 g/L CTAB水溶液,然后接入5 mL热处理过的混合菌菌液,用橡胶塞密封。向发酵瓶中鼓吹N210 min,以确保发酵的厌氧环境。将发酵瓶置于水浴摇床,在37℃转速为150 r/min的条件下发酵,所产生的气体经NaOH碱洗后用排水法收集[10]。
1.3 分析方法
气相色谱检测生物气含量:Porapak Q不锈钢填充柱,柱温80℃,载气为氮气,流速为30 mL/min,进样室温度为80℃,检测室热导检测器(TCD),检测室温度120℃,桥电流120 mA。
1.4 产氢速率的计算
批式发酵的产氢过程可由Logistic模型[11]

进行描述,其中,H为t时刻的累计产氢量,mL;P为产氢过程的产氢潜能,mL;λ为产氢过程的停滞时间,h;Rm为最大产氢速率,mL/h。
2 结果和讨论
2.1 CTAB对胞外膜通透性的影响
CTAB与细菌胞外膜的重要组成部分脂多糖的阳离子结合位点相互作用引起少量脂多糖的溶解,造成胞内的一些蛋白质和遗传物质释放到胞外。因此,CTAB可提高细胞膜的通透性。CTAB的浓度和对菌体的作用时间对上清液A280的影响见图1。图1底部的水平线表明不同浓度的 CTAB水溶液的A280相同,这意味着试验条件下CTAB浓度的高低对溶液的A280值没有影响。因此,一定浓度的CTAB与菌体作用后所得上清液的A280的变化与CTAB的浓度无关,而是单纯由于胞内的一些蛋白质和遗传物质释放到胞外所引起的;不同CTAB浓度下所得上清液A280的大小表征了不同CTAB浓度下细胞膜的通透性的高低。
由图1可看出:当CTAB的浓度小于0.004 8 g/L时,A280未见明显变化;当CTAB的浓度大于0.009 5 g/L时,A280随着CTAB浓度的增加而显著增加。这表明当CTAB的浓度小于0.004 8 g/L时,CTAB对菌体胞外膜的通透性几乎没有影响;而当CTAB的浓度大于0.009 5 g/L时,细胞膜的通透性随着CTAB浓度的提高而提高。浓度为0.057 0 g/L的CTAB与作用15 min后得到A280的值达到2.05,是0.009 5 g/L时的3倍以上,此时细胞膜的通透性严重提高。此外,在相同的CTAB浓度下,CTAB和菌体作用时间由15 min延长至90 min或135 min,A280未见显著提高,这暗示CTAB对菌体胞外膜通透性的改变在15 min内已经完成。

图1 CTAB的浓度和对菌体的作用时间对上清液A280的影响
2.2 CTAB对菌体生长和产酸代谢的影响
不同浓度的CTAB对发酵过程中的生物量的影响如图2所示。试验数据表明:在发酵进行的最初9 h,CTAB的存在对生物量的增长有一定的抑制作用,且随着CTAB浓度的增加抑制作用增强。这与文献[12]报道的高浓度的CTAB有一定的杀菌作用是一致的。发酵进行9 h后,CTAB对生物量增长的抑制作用减弱。这可能是由于生物量的增长使得单位胞外膜所结合的CTAB的分子数下降所致。另一方面,当菌外膜结合适当数量的CTAB时,细菌胞外膜通透性的提高有助于菌体与环境进行物质交换,很可能促进生物量的提高。发酵进行12 h后,添加0.009 5 g/L和0.027 0 g/L CTAB的试验所得生物量达到并略超过空白试验的生物量水平。此外,当发酵进行17 h后,4组试验的微生物的增长都进入稳定期,表明CTAB对微生物的生长周期无明显影响。在0~0.045 0 g/L的CTAB的条件下,发酵终端稳定的OD600值和pH值见图3,图3结果表明:该浓度下CTAB对产酸代谢和最终的生物量无明显影响。

图2 不同浓度的CTAB对发酵过程中的生物量的影响
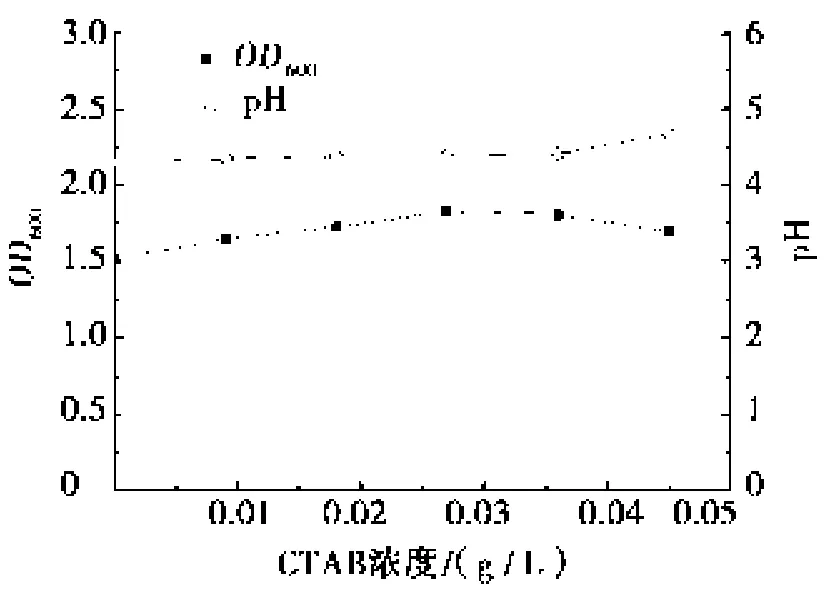
图3 培养基中不同浓度的CTAB对发酵终端的pH和OD600的影响
2.3 CTAB对发酵产氢过程的影响
厌氧发酵所产生的生物气经气相色谱分析未检测到CH4和H2S,仅含有H2和CO2两种气体。这意味着混合菌中的产甲烷菌和硫酸盐还原菌,在初始pH为6.0缺乏有机酸的培养基中几乎没有生长繁殖。生物气经过碱洗装置后所得氢气的含量大于95%。
发酵过程中所产生的氢气体积随时间的变化曲线如图4所示。由图4可以看出:发酵的前8~10 h,CTAB对产氢过程有一定的抑制作用,且随着CTAB浓度的增加抑制作用增强;发酵进行8~10 h后抑制作用减弱。且当CTAB浓度小于0.027 0 g/L时,产氢速率显著提高。这与CTAB对混合菌生物量增长的影响规律一致(见图2)。产氢发酵进行28 h后,不同CTAB浓度条件下的产氢过程全部终止,这暗示CTAB对产氢代谢持续的时间没有影响。值得一提的是,当CTAB浓度在0~0.027 0 g/L的条件下,发酵所得累计产氢量随着CTAB浓度的增加而增加。在0.027 0 g/L的CTAB环境下,获得最大的累计产氢量为304 mL,此时产氢效率为13.6 mmol每克木糖,是对照试验(未添加CTAB)的1.38倍。

图4 不同浓度的CTAB条件下累计产氢体积随发酵时间的变化
用优化的Logistic模型拟合产氢量随时间的变化关系得到较高的相关性(R2≥0.99)。不同CTAB浓度条件下的拟合值,产氢潜力Pm、最大产氢速率Rm和停滞时间λ列于表1中。从表1中可以看出:当培养基中添加0~0.027 0 g/L CTAB时,累计产氢量、最大产氢速率以及产氢潜力随着CTAB浓度的增加而增加。进一步提高CTAB的浓度,累计产氢量、最大产氢速率以及产氢潜力均下降。在添加0.027 0 g/L CTAB的条件下得到最大产氢潜力327 mL和最大产氢速率19.57 mL/h,分别较对照试验提高了45%和44%。这说明通过改变胞外膜通透性能有效促进产氢代谢,这可能与胞内氢气分压的下降对产氢反应的促进作用和对吸氢反应的抑制作用有关。然而进一步提高培养基中CTAB的浓度,又表现出对产氢代谢的显著抑制作用。0.045 0 g/L CTAB的条件下,所得累计产氢量和产氢速率分别为0.027 0 g/L CTAB的条件下的54%和60%,而在该浓度条件下CTAB对终端生物量和产酸代谢并无明显抑制(见图3)。文献[13]报道产气肠杆菌中产氢的关键性酶(氢酶)是膜结合酶。文献[14]报道随着CTAB浓度的增加,生物酶的活性下降。因此,过高浓度的CTAB对产氢过程的负面作用可能是由于氢酶的活性受到高浓度的CTAB的抑制引起的,这有待于进一步研究。
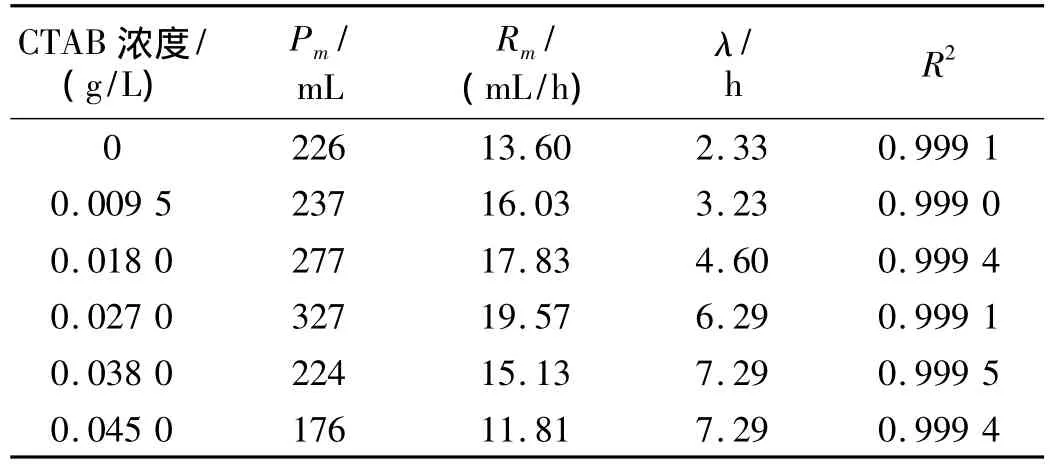
表1 不同浓度的CTAB 对发酵过程中的动力学参数的影响
另外,CTAB对产氢发酵的停滞时间也有显著影响,且随着CTAB浓度的增加停滞时间增加。在0.045 0 g/L的CTAB条件下,停滞时间由对照试验的2.33 h增加至7.29 h。CTAB对产氢发酵的停滞时间的影响与CTAB对生物量增长的影响有关(见图2)。
2.4 化学添加剂强化产氢发酵的文献对比
本文研究得出CTAB是用于暗发酵产氢的一种有效的化学添加剂。文献报道的化学添加剂对光发酵和暗发酵产氢的影响总结如表2所示。由表2可以看出:烟酸和MoO2-4对光发酵产氢过程的强化作用显著,在适宜浓度的烟酸或者MoO2-4的环境下,产氢速率的强化倍数可达到3以上。各种添加剂对暗发酵过程中产氢速率和累计产氢量的强化倍数一般都小于2。本研究中CTAB与其他用于暗发酵的添加剂相比,累计产氢量和产氢速率的强化倍数相当,但CTAB的用量远远小于其他添加剂。
各种添加剂对产氢过程的作用机理不同。Ni2+,Fe2+和MoO2-4是氢酶或者钼固氮酶的组成成分,所以添加Ni2+,Fe2+和MoO24促进了产氢的原因可能与氢酶或者钼固氮酶活性的提高有关。添加乳酸对产氢过程的强化可能是由于甲酸和乙醇的代谢途径向丁酸代谢途径转换所致[3]。有关L-胱氨酸的强化机理目前尚不清楚。本研究中CTAB对产氢发酵的强化作用可能是由于CTAB与胞外膜结合引起胞外膜溶解而使胞外膜的通透性提高。
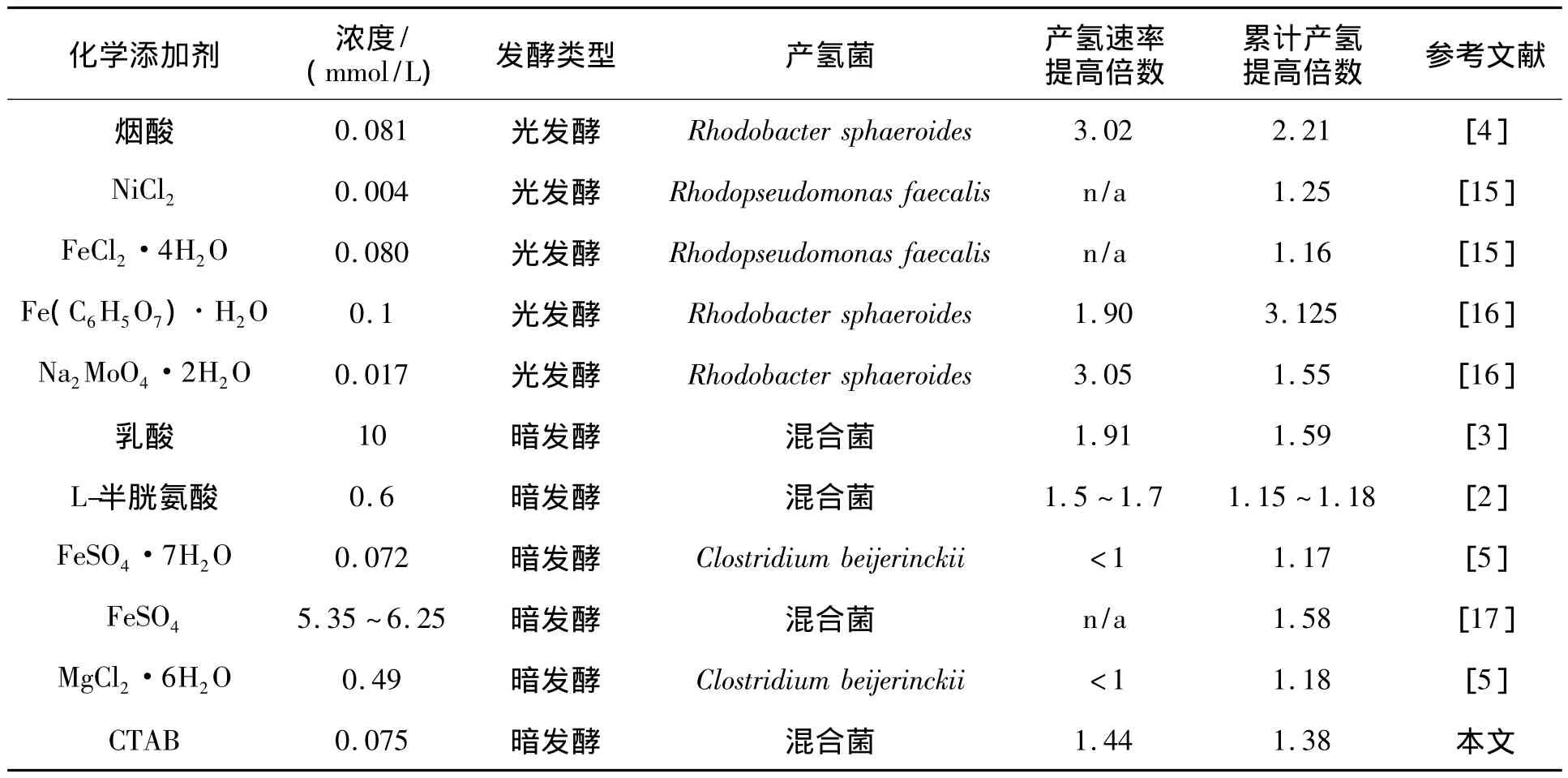
表2 通过化学添加剂强化产氢发酵的文献对比
3 结论
本研究通过在培养基中添加微量的CTAB,实现了强化发酵产氢的目的。CTAB对混合菌胞外膜通透性,对发酵过程中产氢速率、累计产氢量、生物量增长和产氢停滞时间都有一定程度的影响。
(1)浓度大于0.009 5 g/L的CTAB溶液可有效提高混合菌菌体胞外膜的通透性。
(2)在发酵过程中的前9 h内,CTAB对混合菌的生长繁殖有一定的抑制作用,但浓度小于0.045 0 g/L的CTAB对发酵终端的生物量没有明显影响;CTAB的存在使得产氢发酵过程中的停滞时间延长,且随着CTAB浓度的增加而增加。
(3)0.095 0~0.027 0 g/L的CTAB可有效地强化混合菌的产氢发酵过程。并在CTAB浓度为0.027 0 g/L时得到最大产氢效率(13.6 mmol每克木糖)和最大的产氢速率(19.6 mL/h),与对照试验相比分别提高38%和44%。
[1]Li Z,Wang H,Tang Z,et al.Effects of pH Value and Substrate Concentration on Hydrogen Production from the Anaerobic Fermentation of Glucose[J].Int J Hydrogen Energ,2008,33(24):7413-7418.
[2]Yuan Z,Yang H,Zhi X,et al.Enhancement Effect of L-cysteine on Dark Fermentative Hydrogen Production[J].Int J Hydrogen Energ,2008,33(22):6535-6540.
[3]Baghchehsaraee B,Nakhla G,Karamanev D,et al.Effect of Extrinsic Lactic Acid on Fermentative Hydrogen Production[J].Int J Hydrogen Energ,2009,34(6):2573-2579.
[4]Li X,Shi H,Wang Y,et al.Effects of Vitamins(Nicotinic Acid,Vitamin B1 and Biotin)on Phototrophic Hydrogen Production by Rhodobacter Sphaeroides ZX-5[J].Int J Hydrogen Energ,2011,36(16):9620-9625.
[5]Zhao X,Xing D,Liu B,et al.The Effects of Metal Ions and L-cysteine on HydA Gene Expression and Hydrogen Production by Clostridium Beijerinckii RZF-1108[J].Int J Hydrogen Energ,2012,37(18):13711-13717.
[6]Clark I C,Zhang R H,Upadhyaya S K.The Effect of Low Pressure and Mixing on Biological Hydrogen Production via Anaerobic Fermentation[J].Int J Hydrogen Energ,2012,37(15):11504-11513.
[7]Lee K S,Tseng T S,Liu Y W,et al.Enhancing the Performance of Dark Fermentative Hydrogen Production Using a Reduced Pressure Fermentation Strategy[J].Int J Hydrogen Energ,2012,37(20):15556-15562.
[8]崔建东,贾士儒,谭之磊.改善苯丙氨酸解氨酶重组大肠杆菌细胞通透性提高反式肉桂酸转化率[J].高校化学工程学报,2008,22(6):1015-1019.
[9]Ren Y L,Xing X H,Zhang C,et al.A Simplified Method for Assay of Hydrogenase Activities of H2Evolution and Uptake in Enterobacter Aerogenes[J].Biotechnol Lett,2005,27(14):1029-1033.
[10]Ren Y L,Wang J,Liu Z,et al.Hydrogen Production from the Monomeric Sugars Hydrolyzed from Hemicellulose by Enterobacter Aerogenes[J].Renew Energ,2009,34(12):2774-2779.
[11]Wang J,Wan W.The Effect of Substrate Concentration on Biohydrogen Production by Using Kinetic Models[J].Sci China Ser B,2008,51(11):1110-1117.
[12]Salton M R J.The Adsorption of Cetyltrimethylammonium Bromide by Bacteria,Its Action in Releasing Cellular Constituents and Its Bactericidal Effects[J].J Gen Microbiol,1951,5(2):391-404.
[13]Nakashimada Y,Rachman M A,Kakizono T,et al.Hydrogen Production of Enterobacter Aerogenes Altered by Extracellular and Intracellular Redox States[J].International Journal of Hydrogen Energy,2002,27:1399-1405.
[14]Liu H Z,Yang W J,Chen J Y.Effects of Surfactants on Emulsification and Secondary Structure of Lysozyme in Aqueous Solutions[J].Biochem Eng J,1998,2(3):187-196.
[15]Liu B F,Ren N Q,Ding J,et al.The Effect of Ni2+,Fe2+and Mg2+Concentration on Photo-hydrogen Production by Rhodopseudomonas Faecalis RLD-53[J].Int J Hydrogen Energ,2009,34(2):721-726.
[16]Eroglu E,Gunduz U,Yucel M,et al.Effect of Iron and Molybdenum Addition on Photofermentative Hydrogen Production from Olive Mill Wastewater[J].Int J Hydrogen Energ,2011,36(10):5895-5903.
[17]Wang J,Wan W.Effect of Fe2+Concentration on Fermentative Hydrogen Production by Mixed Cultures[J].Int J Hydrogen Energ,2008,33(4):1215-1220.
