自然降温过程中5种樟树的抗寒性分析
2014-04-08袁美丽苏金乐
王 宁,姚 方,袁美丽,苏金乐
(1.河南科技大学林学院, 河南 洛阳 471000; 2.河南林业职业学院, 河南 洛阳 471002; 3.洛阳市隋唐城遗址植物园, 河南 洛阳 471002; 4.河南农业大学林学院, 河南 郑州 450002)
低温是限制植物北引的主要环境胁迫因子[1].随着全球气候的逐渐变暖,南方树种有望逐渐向北方地区引种.樟树为樟科(Lauraceae)樟属(Cinnamomum)植物,中国不同种樟树有15种之多.樟树的抗寒生理性研究多集中于某一个种或类型,如张亚红等[2]、李凌等[3]结合自然降温过程中MDA含量等相关指标的变化,对香樟抗寒性进行了研究.薛建辉等[4]结合自然降温过程中叶片相对电导率和低温半致死温度(LT50)测定,对南京地区引种的浙江樟等5种常绿阔叶树种进行了抗寒性研究,得出浙江樟抗寒能力较强,且能在南京地区广泛栽植的结论.尤扬等[5]研究认为,当温度低于-10 ℃时香樟容易发生冻害.毛春英等[6,7]在山东泰安对香樟幼苗及种子分别进行了引种研究,结果表明引种种子进行播种繁殖更易成功,并能获得抗寒性强的苗木.对多种樟树进行抗寒对比性研究相对较少,仅见有王宁等[8~10]结合叶片解剖结构,以及低温胁迫过程中叶片超微结构、叶绿素荧光参数的变化,对猴樟等5种樟树的抗寒性进行了研究.姚方等[11]结合自然降温过程中渗透调节物质含量的动态变化,对尾叶樟等3种樟树的抗寒性进行了研究.宋芳林等[12]结合自然降温过程中保护酶活性及脂肪酸含量的变化,对猴樟及香樟进行了抗寒性研究.谢晓金等[13]借助电导法,对南京地区露地引种的包括香樟、浙江樟等12种常绿阔叶树种进行了抗寒性研究,研究结果表明1月中旬抗寒能力达到最强,而通常10月份或3月份期间抗寒能力最弱.本研究以郑州市上街区已引种多年不同种或类型的樟树为试材,通过对自然降温过程中其叶片膜透性、保护酶活性及渗透调节物含量动态变化的研究,以期更加全面科学的鉴定不同樟树的抗寒性,揭示其在越冬过程中的适应机制.
1 材料与方法
1.1试验地概况
试验地位于河南省郑州市上街区,地理坐标东经113°14′45″~113°19′05″,北纬34°35′~34°40′.暖温带季风气候,年平均气温13.5~15.3 ℃,极端最低温为-16.5 ℃.土壤为褐土类.
1.2试验材料与设计
选取上街区登封路行道树中引种栽植的8 a生尾叶樟(CinnamomumcamphoraKisterm)、猴樟(C.bodinieriLevel)、沉水樟(C.micranthum(Hay.) Hay)、芳樟(C.camphoraLinn (linalool-type))及本樟(C.camphoraLinn (cincol-type))共5种或类型,每种或类型选择生长势相近的3株作为研究对象,其生长状况如表1所示.为便于叙述,将上述树种依次标号为I,II,III,IV及V.
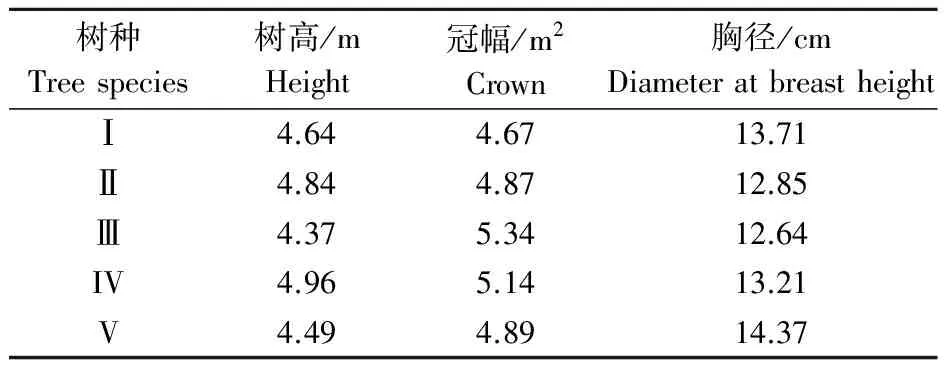
表1 供试樟树的生长状况
以秋季气温明显下降为起始时间(2009-10-30),翌年春季气温回升平缓后为终止日期(2010-04-16),共持续169 d.采样日期及日最高、最低温如表2所示.考虑到温度变化的不规律性,通常于明显降温后出现的晴天进行取样,每月至少1次,2009-11-03采第1次样,作为对照(CK),共取样11次.随机采取树冠中上部不同方向,当年生枝的第3~7枚完好叶片,每树种30枚.采后立即用潮湿纱布包裹,自封袋密封后带回试验室,依次用自来水和蒸馏水冲洗干净并吸干水分,取部分叶片立即测定相对电导率,其余置超低温冰箱保存,用于其它生理指标的测定.
1.3测定项目及方法
参照邹琦[14]的电导法进行电解质渗透率的测定.参照李合生[15]的方法,丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)法,超氧化物歧化酶(SOD)活性的测定采用NBT光照化学还原法,酸性茚三酮测定脯氨酸含量.
1.4数据处理
应用Fuzzy数学中隶属函数法进行综合评判[16,17].对与抗寒性呈负相关的电导率及MDA含量,采用公式:
f(Xij)=1-(Xij-Xj min)/(Xj max-Xj min)
(1)
对与抗寒性正相关的SOD活性和脯氨酸,采用公式:
f(Xij)=(Xij-Xj min)/(Xj max-Xj min)
(2)
式中:f(Xij)为i树种的j项指标隶属函数值;Xij为其测定值;Xj max和Xj min分别为j指标最大测定值和最小测定值.试验数据用Excel及SPSS统计软件进行相关分析和方差分析.
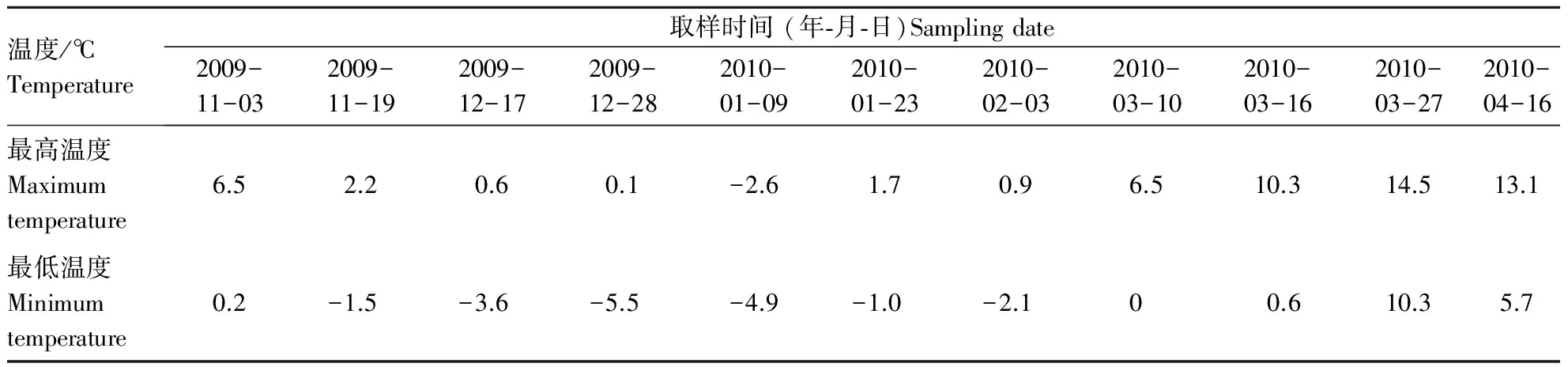
表2 采样时间及日最高和最低温度
2 结果与分析
2.1相对电导率的变化
越冬期间,5种樟树叶片的质膜透性整体呈先升后降再升的变化趋势(表3).I,Ⅲ树种的电导率峰值出现在2010-01-09,表明2010-01-05─2010-01-08持续-8 ℃左右的低温对其造成的影响较大,与CK相比分别增加了223.00%和241.00 %;而II,IV,V的峰值均出现2010-01-23,表明连续的低温对其造成的影响较小,这可能与其保护酶活性及渗透调节物质含量的增加相关,较CK分别增加了116.00%,167.00%和180.00%,均明显小于树种Ⅲ的增加量.除II外,其余树种电导率在4月份均有不同程度的增加,表明其忍受“倒春寒”的能力不尽相同,与2010-03-27电导率相比,I,III,IV,V分别增加了25.00%,67.26%,15.38%及21.76%,增加顺序为III >I>V>IV,由此表明“倒春寒”对III的影响最大,其次是I,V,然后是IV,II在“倒春寒”期间受到的影响相对较小.
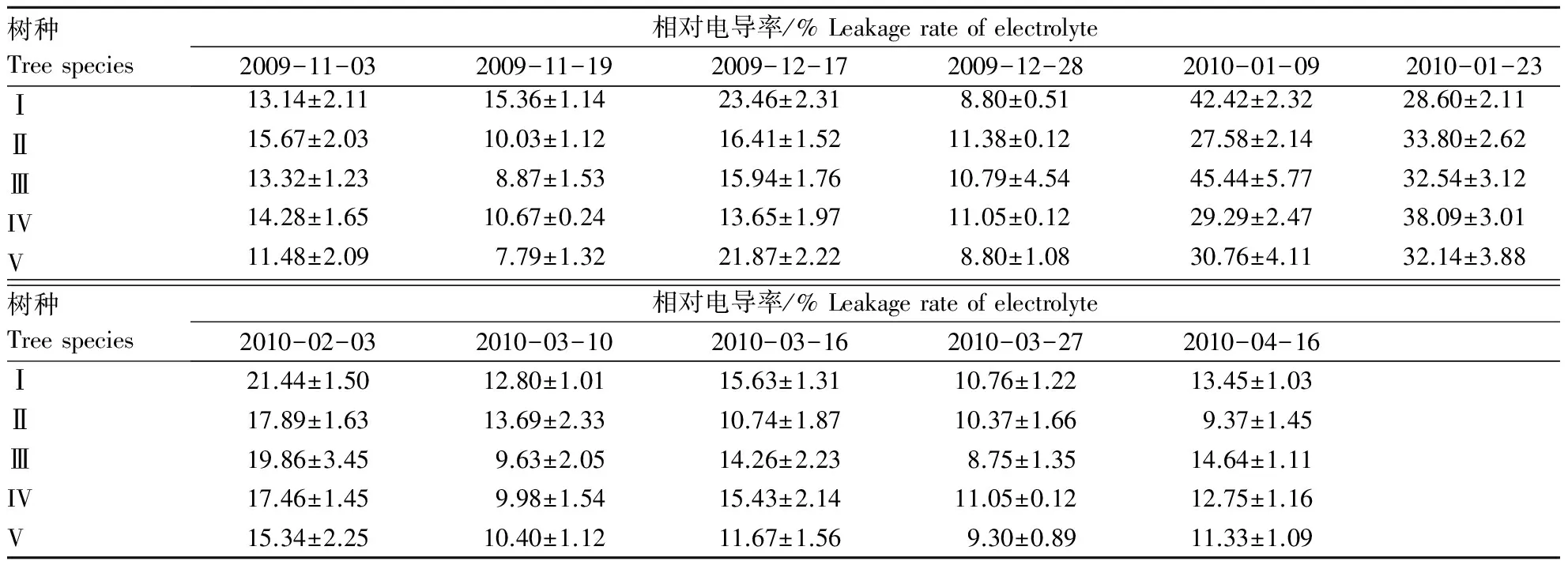
表3 自然降温对5种樟树叶片相对电导率的影响
2.2MDA含量的变化
5种樟树叶片细胞膜脂过氧化程度随着自然界温度的降低而逐渐增加,春季则随着温度的升高而逐渐降低(表4).自然降温过程中,前期降温过程中5种樟树的膜脂过氧化程度增加较为缓慢,其中以III增加最大;而2010-01-05─2010-01-08 持续-8 ℃左右的低温出现后,MDA含量增加较为明显,2010-01-09较2009-12-28分别增加了89.73 %,11.10 %,93.17 %,19.75 %和24.32 %,增加顺序为III>I>V>IV>II,表明持续的低温胁迫对III造成的伤害最大;与CK相比,I,Ⅲ的MDA峰值出现在2010-01-09,分别增加了116.00%和154.00%,而II,IV,V的峰值均出现2010-01-23,分别增加了63.00%,97.94%和121.00%,可以看出以III增加最为明显,且从2009-12-17─2010-03-16,III的MDA含量明显高于同时期其它树种.4月份的再次降温胁迫中,除II外,其余樟树MDA含量均有不同程度的增加,表明不同树种对“倒春寒”的适应性不同.

表4 自然降温对5种樟树叶片MDA含量的影响
2.3SOD活性的变化
越冬期间,随着气温的下降,几种樟树叶片内SOD活性均出现迅速增加(表5).I,III峰值出现在2009-12-28,而II,IV,V峰值出现在2010-01-09,随后均出现明显的下降,可能是持续低温增加了膜脂过氧化程度,导致植物体内的酶促防御系统受到危害,造成未得到及时清除活性氧的积累,从而降低了SOD活性.与2009-11-03相比,5种樟树SOD值分别增加了88.17 %,71.30 %,113.00 %,103.00 %和95.50 %,增加顺序为II>IV>V>I>III,表明II对低温胁迫的适应能力最强,其次IV,V,最弱的为III;此后,随着春季温度的逐渐回升,酶促防御系统逐渐得到恢复.至2010-03-27,与2010-01-09相比,I,III的SOD活性分别增加了44.94 %和38.67 %,II,IV,V的SOD活性与2010-01-23相比分别增加了52.75 %,50.56 %和46.13 %,可以看出,以II增幅最大,表明其酶促防御系统受害较轻,恢复也相对较快,其次为IV,V,而增加最少的为III,表明自然降温过程中其受伤害相对较重.4月份的再次降温造成了SOD活性小幅上升,与2010-03-27相比,其中以II增幅最大,为14.57 %,而III增幅最小,仅为4.50 %,表明III对“倒春寒”的适应性最弱,而II的适应性最强.

表5 自然降温对5种樟树叶片SOD活性的影响
2.4脯氨酸含量的变化
越冬期间,随着温度的下降,5种樟树叶片内脯氨酸含量均出现持续的增加,之后随着春季温度的回升又呈逐渐降低(表6).5种樟树的峰值均出现在2010-01-09,与2009-11-03相比,分别增加了115.00 %,329.00 %,71.44 %、200.00 %和139.00 %,其中以II增加最大,III增加最小,表明II对低温的适应能力最强,而III最弱.前期降温过程中,与2009-12-17相比,至2009-12-28,5个树种脯氨酸含量分别增加了15.15 %,18.37 %,12.74 %,17.94 %和13.35 %,增加顺序为II > IV > V > I > III,而与2009-12-28相比,至2010-01-09,持续-8 ℃左右低温后,脯氨酸含量分别增加了13.18 %,56.63 %,11.50 %,21.51 %和13.60 %,其中以II增加最为明显,与前面的增加情况相比,除II,IV增幅明显增加外,I,III虽有增加,但小于前面的增幅,V大致持平,由此说明,持续-8 ℃左右的低温胁迫后,II,IV依然通过脯氨酸的大幅增加提高其对低温的适应能力,而I,III的适应能力却逐渐减弱.4月份的“倒春寒”胁迫中,5个树种的脯氨酸含量均有不同程度的增加,与2010-03-27相比,以II增加最为明显,达24.41 %,而III增幅最小,仅为8.82 %.
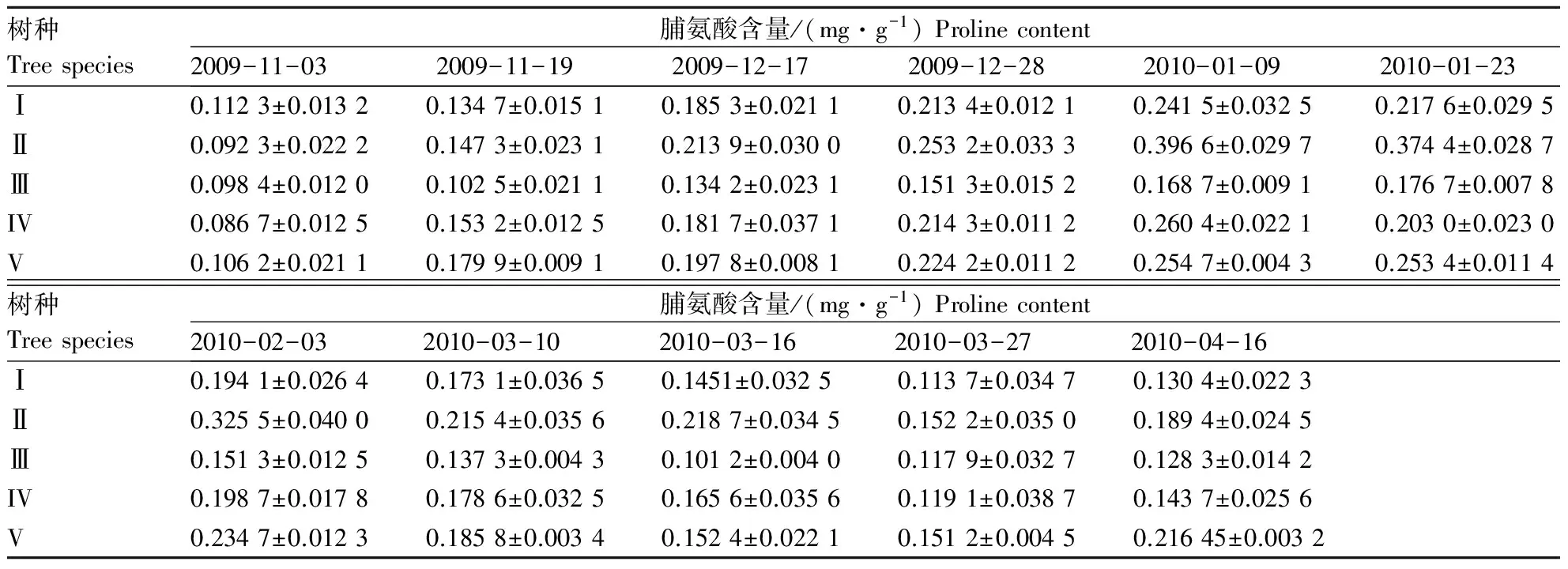
表6 自然降温对5种樟树叶片脯氨酸含量的影响
2.5隶属函数法综合分析抗寒性
2.5.1 自然降温过程中各抗寒指标隶属值 以Ⅱ为例,运用隶属函数法求出其加权平均值D,值越大表示抗寒性越强(表7).从表7可以看出,综合评价值呈“升-降-升”的变化趋势,由此表明,越冬期间猴樟的抗寒性呈现增强、减弱再增强的变化.依次可将其抗寒性分为2个时期,分别为:(1)增强期.分2个阶段,2009-11-03─2009-12-17,随着气温的降低,通过渗透调节物质含量和保护酶活性的增加,从而增强其对低温胁迫的抵御能力;2010-01-09─2010-04-16,随着气温的逐渐回升,受低温胁迫伤害的细胞膜逐渐得到恢复,樟树的抗寒性逐渐得到恢复.(2)减弱期.2010-01,随着2010-01-05─2010-01-08持续 -8 ℃左右的低温出现,抗寒性迅速降低,这可能与低温的过渡胁迫所致.
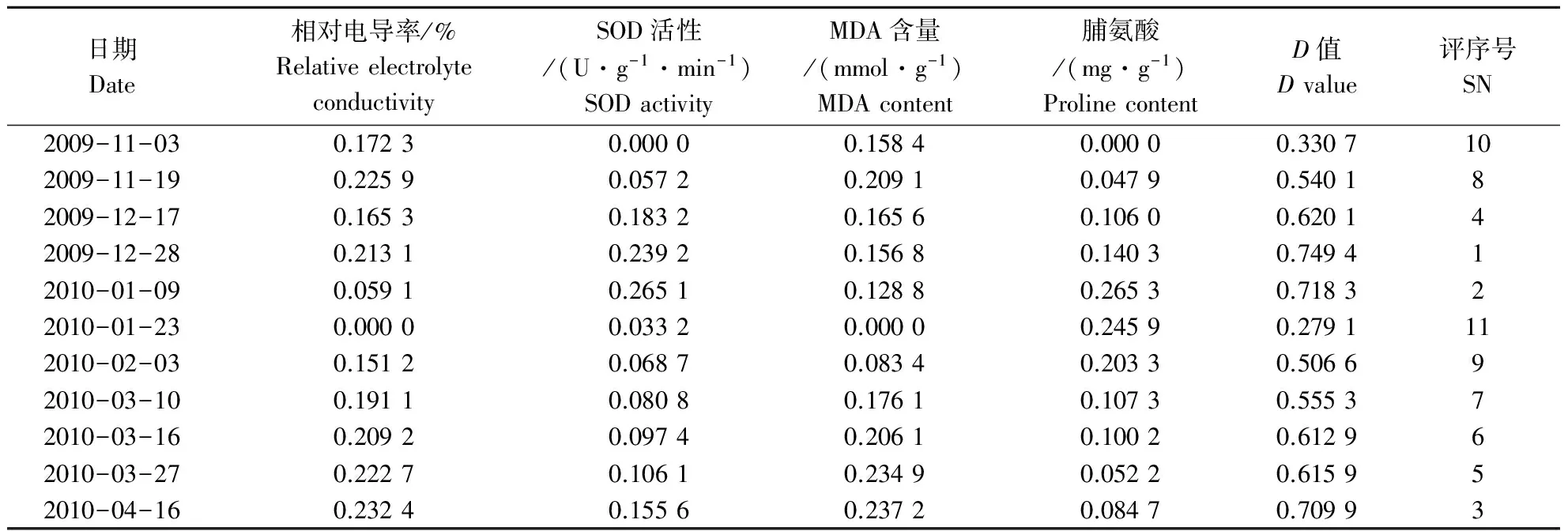
表7 自然降温过程中各抗寒指标参数的D值
2.5.2 几种樟树的抗寒性综合评价 低温胁迫过程中,由于受到多重因素的影响,植物的生理变化相对较为复杂,因此,孤立的应用某一种指标较难真实反映植物的抗寒性[18,19].因此,为了较为全面地反映5种或类型樟树的抗寒性强弱,结合隶属函数法的运用,求出几种樟树各指标参数的隶属函数加权平均值D,该值越大表明抗寒性越强(表8).从表8可以看出,抗寒性强弱顺序为:II>IV>V>I>III.其中以猴樟的D值最大,表明其耐寒性最强,而沉水樟耐寒性最差.
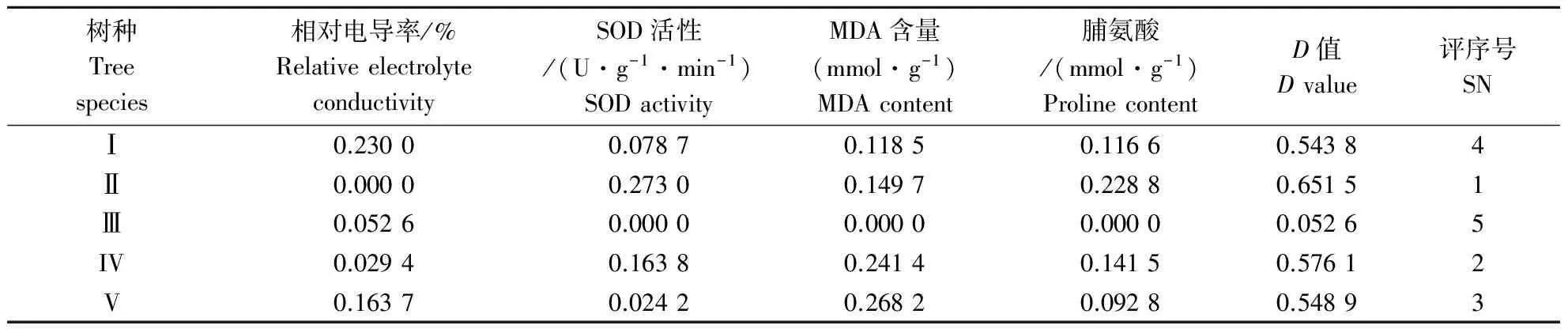
表8 5种樟树的D值及抗寒性综合评价
3 小结与讨论
3.1细胞膜透性与樟树的抗寒性关系
大量研究认为,低温造成植物发生寒害的根本原因是其对细胞膜系统造成的损害,因此细胞膜系统的流动性和稳定性与植物抗寒性之间关系密切[20~22].早在1930年,DEXTER等[23]发现植物电导率的大小与其组织受低温伤害的程度呈正相关,并首次利用电导法测定离体植物材料的抗寒强弱.随后,电导法作为测定植物抗寒性快速可靠的方法之一被广泛的采用.杨家胭等[24]、刘祖祺等[25]先后采用电导法测定了柑桔的抗寒性,发现柑桔叶片及枝条的电导率随着胁迫温度的降低而增加,并发现电导具有较好的灵敏度和精确度;BIGRAS等[26]、HAWKINS等[27]采用电导法对黑云杉及黄扁柏的耐寒性进行了研究,并得出了理想的结果.本研究结果表明,自然降温过程中,尽管几种樟树细胞内相对电导率的峰值出现日期不尽相同,但整体变化趋于一致.降温初期电导率的上升较为平缓,然后,随着2010-01温度的持续降低,相对电导率出现急剧升高,这与向邓云等[28]对自然降温过程中曼地亚红豆杉和鲍思伟[29]对云锦杜鹃的研究结果一致.与CK相比,其中,以III增幅最大,II最小,表明III电解质外渗严重,细胞膜受到了伤害,其抗寒性也最弱,而以II的抗寒性为最强.2010-04再次降温中,除II外,其余树种电导率均有不同程度的增加,而其中又以III增幅最大,表明其对“倒春寒”的适应能力最弱.
3.2膜脂过氧化与樟树的抗寒性关系
低温胁迫条件下,植物细胞内产生大量的自由基,增加的自由基首先攻击膜系统,同时氧化膜质脂肪酸的不饱和键,造成MDA含量的增加[30].因此,通常以MDA含量作为膜脂过氧化反应的主要指标,用来反映植物细胞膜脂过氧化的程度,以及植物对低温胁迫的适应性强弱[31,32].
本研究中,自然降温过程中,随着温度的降低,5种樟树的MDA含量均出现不同程度的增加,这与庄炳莉[33]对自然降温过程中阔瓣含笑、亮叶含笑等5个常绿阔叶树种的研究一致.5种樟树MDA含量在2009-12的增幅明显小于2010-01,表明1月份的持续低温胁迫加剧了5种樟树膜脂过氧化程度,而其中又均以III增幅最大,II增幅最小,表明自然降温对III造成的伤害最大.4月份的再次降温中,除II外,其余樟树MDA含量均有不同程度的增加,表明不同树种对“倒春寒”的适应性不同,其中又以III增幅最大,表明其对“倒春寒”的适应能力最弱.
3.3保护酶活性与樟树的抗寒性关系
在一定的低温胁迫中,保护酶活性的升高有利于保持植物体内自由基的产生和清除之间的平衡,随着低温的持续降低,此动态平衡被打破,细胞的清除系统遭到损伤,活性氧对细胞产生氧化胁迫[34].大量研究表明,植物体内SOD活性的高低直接反映了活性氧的清除能力大小, 与植物的抗逆性关系密切[35,36].
本研究结果表明,在自然降温过程中,各种樟树叶片内SOD活性变化趋势基本一致,降温过程中,各种樟树通过SOD活性持续增加以适应外界的低温胁迫,而后随着春季温度的逐渐回升,细胞酶促防御系统逐渐得到恢复,SOD活性出现小幅上升,这与应叶青等[37]对自然降温过程中毛竹中的SOD变化的研究结果一致.与CK相比,至SOD活性峰值,降温胁迫中各种樟树SOD活性的增幅大小顺序为II > IV > V > I > III,表明II的活性氧清除能力相对较大,其抗寒性也最强,而以III最弱.而后随着春季温度的回升,各种樟树SOD活性均出现小幅上升,其中又以II增幅最大,III的增幅最小,说明II细胞内酶促防御系统在前期的降温胁迫中受到伤害相对较小,因此恢复也相对较快,而III则相反.
3.4渗透调节物质含量与樟树抗寒性关系
冬季来临之前,植物发生一系列形态和生理生化的变化以适应低温胁迫,即所谓的抗寒锻炼.低温条件下,植物细胞通过渗透调节以维持细胞内外的渗透平衡,因此必然使机体内如糖、蛋白质、脯氨酸等参与渗透调节的物质含量发生变化,从而提高植物的耐寒力,避免冻害的发生[38].
脯氨酸是水合力较强的氨基酸,其含量的增加是植物对逆境胁迫的一种生理生化反应,是植物对逆境的适应表现.脯氨酸具有多种生理功能,如作为细胞质渗透调节物质、稳定生物大分子结构等[39].DELAUMEY等[40]研究认为,作为细胞质内的渗透调节物质,脯氨酸积累的多少与植物的抗逆性相关,可作为抗逆性筛选的指标之一.本研究结果表明,随着气温的降低,5种樟树的脯氨酸含量均出现不同程度的增加,与2009-12的增幅相比,2010-01的持续低温出现后,II,IV增幅依然明显增加,而I,III的增幅却出现减少现象,其中又以III的增幅减少最多,由此说明随着低温的持续胁迫,II,IV依然具有较强的适应性,而III适应性则相对最弱.春季随着温度的逐渐回升,各种樟树的脯氨酸含量出现不同程度的减少,这与苏敬[41]对浙江樟、乐昌含笑等5种常绿阔叶树种的研究结果一致.
3.5综合性抗寒评价
对多个指标或因子共存的系统进行综合评价,通常采用模糊数学中的相关理论和方法.基于隶属函数法具有简单及准确等优点,目前已被广泛的应用于生态学籍环境科学的研究领域.
自然降温过程中,植物耐寒性的一系列变化是多个因子综合作用的结果,因此,单一抗寒指标难以判断其对冬季低温的综合适应能力[16].庄炳莉[33]、苏敬[41]及绍文鹏[34]利用隶属函数法,分别对自然降温过程中多种常绿阔叶树种进行了抗寒性综合评价研究,并得出理想的结果.本研究结果表明,通过隶属函数法的应用,对自然降温过程中5种或类型樟树的MDA、SOD等4个生理生化指标进行了综合分析,得出抗寒性强弱顺序为:II>IV>V>I>III.
以猴樟为例,采用隶属函数法,对自然降温过程中4个生理生化指标参数进行综合分析,结果显示,越冬期间,其抗寒性分为增强期、减弱期2个阶段.随着秋季气温的逐渐降低,樟树通过保护酶活性及渗透调节物质含量的增加,提高其抗寒性,为冬季低温胁迫奠定基础,这个时期即所谓的抗寒锻炼期;之后,随着2010-01-05─2010-01-08持续-8 ℃的低温,过渡的低温胁迫导致细胞膜透性变大,电解质外渗随之增加,同时膜脂过氧化程度逐渐增加,共同导致其抗寒性减弱;春季,随着气温的逐渐回升,受低温胁迫的伤害细胞酶促防御系统逐渐得到恢复,细胞内部活性氧自由基的清除能力随之增加,樟树的抗寒性也逐渐得到恢复.
参考文献:
[1]STUSHNOFF C, JUNTTILA O. Seasonal development of cold stress resistance in several plant species at a coastal and a continental location in North Norway [J].Polar Biology, 1986, 15: 129-133.
[2]张亚红,孙存华,蒋继宏,等.自然降温过程中香樟和广玉兰的抗寒性研究[J].安徽农业科学,2010,38(16):8653-8657.
[3]李 凌,袁同印.自然越冬状态下香樟抗寒性生理指标的动态变化[J].江西农业学报,2012,24(12):46-48.
[4]薛建辉,苏 敬,田如男,等.自然降温过程中5个常绿阔叶树种的抗寒性分析[J].植物资源与环境学报,2009, 18(1):52-56.
[5]尤 扬,刘 弘,吴荣升,等.低温胁迫对香樟幼树抗寒性的影响[J].广东农业科学,2008(11):23-25.
[6]毛春英.香樟的引种与驯化研究[J].山东农业大学学报:自然科学版,2004,35(4):534-539.
[7]毛春英,张纪德,王秀梅.樟树引种驯化及抗寒育苗栽培技术[J].林业科技,2001,26(6):10-12.
[8]王 宁,袁美丽,苏金乐.几种樟树叶片结构比较分析及其与抗寒性评价的研究[J].西北林学院学报,2013,28(4):43-49.
[9]王 宁,董莹莹,苏金乐.低温胁迫下2种樟树叶片超微结构的比较[J].西北农林科技大学学报,2013,41(7):106-112.
[10] 王 宁,姚 方,袁美丽,等.自然降温对猴樟和芳樟叶绿素荧光参数的影响[J].河南农业大学学报,2013,47(4):414-419.
[11] 姚 方,吴国新,梅海军.自然降温过程中3种樟树渗透调节物质的动态变化[J].华南农业大学学报,2012,33(3):378-383.
[12] 宋芳林,张苗苗,苏金乐,等.自然降温过程中猴樟和香樟幼苗的抗寒性比较[J].西部林业科学,2012,41(6):48-52.
[13] 谢晓金,郝日明.南京地区12种常绿阔叶树种冬季抗寒性动态变化[J].生态学报,2009,29(4):2149-2154.
[14] 邹 琦.植物生理生化实验指导[M].北京:中国农业出版社,2000.
[15] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[16] 李轶冰,杨顺强,任广鑫,等.低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J].生态学报,2009,29(3):1341-1347.
[17] 田钦谟.模糊综合评价中的若干问题[J].模糊系统与数学,1996,10(2):62-69.
[18] 杨敏生,裴保华,程志鹏.白杨杂种无性系抗寒性生理指标动态分析[J].生态学报,1997,21(4): 367-375.
[19] 沈 漫,王明庥,黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997,14(2): 1-8.
[20] 简令成.生物膜与植物寒害和抗寒性的关系[J].植物学通报, 1983(1):17-23.
[21] 滕中华,周党卫,师生波,等.青藏高原三种高寒植物的质膜透性变化与抗寒性的关系[J].中国草地,2001, 23(4):37-47.
[22] 严寒静,谈 锋.自然降温过程中栀子叶片膜保护系统的变化与低温半致死温度的关系[J].植物生态学报, 2000,24(1):91-95.
[23] DEXTER S T, TOTTINGHAM W E, GRABER L F. Preliminary results in measuring the hardiness of plants [J].Plant physiology,1930,5: 215-223.
[24] 杨家胭,刘祖祺,刘谷良.电导法测定柑桔耐寒性的灵敏度和精确性的检验[J].南京农业大学学报,1980(1): 87-95.
[25] 刘祖祺,周碧英,王元裕,等.电导法鉴定柑桔耐寒性的试验[J].南京农业大学学报,1981,2(6):32-37.
[26] BIGRAS F J, CALME S. Viability test for estimating root cold tolerance of black spruce seedlings [J].Can Jou For Res,1994,24(5):1039-1048.
[27] HAWKINS B J, RUSSELL J, SHORN R. Effect of population, environment and maturation on the frost hardiness of yellow-ceder [J].Canadian Journal of Forest Research,1994, 24(5):945-953.
[28] 向邓云,谢吉容,谈 锋.自然降温过程中曼地亚红豆杉叶片膜保护系统的变化与低温半致死温度的关系[J].西南师范大学学报:自然科学版, 2001,26(4):452-456.
[29] 鲍思伟.云锦杜鹃低温半致死温度对自然降温的适应[J].西南民族大学学报:自然科学版,2005,31(1): 99-102.
[30] 许凯杨,叶万辉,沈 浩,等.低温胁迫下喜旱莲子草幼苗膜脂过氧化及保护酶活性的变化[J].生态科学,2006,25(2):139-142.
[31] 潘晓云,曹琴东,王根轩.膜脂过氧化作为扁桃品种抗寒性鉴定指标研究[J].生态学报,2002,22(11): 1902-1911.
[32] 高述民,陈朋军,郭惠红,等.日本桃叶珊瑚的冷驯化及抗寒机制研究[J].西北植物学报,2003,23(12): 2113-2119.
[33] 庄炳莉.自然降温过程中5个常绿阔叶树种的耐寒性研究[D].南京:南京林业大学, 2008.
[34] 绍文鹏.几种常绿阔叶植物抗寒性研究[D].泰安:山东农业大学, 2009.
[35] PRASAS T K. Mechanism of chilling-induced oxidative stress injury and tolerance: changes in antioxidant system, oxidation of proteins and lipids and protease activities [J]. Plant J,1996,10:1017-1026.
[36] PRASAD T K, ANDERSON M P, MARTIN B A, et al. Evidence for chilling induced oxidative stress in maize seedling and a regulatory role for hydrogen peroxide [J].Plant Cell,1994,6:65-74.
[37] 应叶青,魏建芬,解楠楠,等.自然低温胁迫对毛竹生理生化特性的影响[J].南京林业大学学报:自然科学版, 2011,35(3):133-136.
[38] 王 瑾,廖 康,王燕凌,等.秋季自然降温对野生樱桃李叶片渗透调节物质的影响[J].新疆农业科学,2010, 47(5):952-957.
[39] 覃光球,严重玲,韦莉莉.秋茄幼苗叶片单宁、可溶性糖和脯氨酸含量对Cd胁迫的响应[J].生态学报,2006,26(10):3366-3370.
[40] DELAUMWY A J, VERMA D P. Proline bilsynthesis and osmoregulation in plants [J].Plant J,1993,4: 215-223.
[41] 苏 敬.自然降过程中五个常绿阔叶树种的抗寒性研究[D].南京:南京林业大学, 2007.
