冻融作用下寒温带针叶林土壤碳氮矿化过程研究
2014-02-27罗亚晨吕瑜良杨浩何念鹏李胜功高文龙
罗亚晨,吕瑜良,杨浩,何念鹏,李胜功,高文龙,3
1. 西南大学地理科学学院,重庆 400715;2. 中国科学院地理科学与资源研究所,生态系统观测与模拟重点实验室,北京 100101;3. 中国科学院大学,北京 10049
冻融作用下寒温带针叶林土壤碳氮矿化过程研究
罗亚晨1,2,吕瑜良1,杨浩2*,何念鹏2,李胜功2,高文龙2,3
1. 西南大学地理科学学院,重庆 400715;2. 中国科学院地理科学与资源研究所,生态系统观测与模拟重点实验室,北京 100101;3. 中国科学院大学,北京 10049
以大兴安岭落叶松林土壤为研究对象,设置8 ℃恒温和-5~8 ℃冻融循环(1个冻融循环为在-5 ℃培养24 h,后在8 ℃培养24 h)2个处理,进行30 d的室内培养实验,探讨了寒温带针叶林土壤在冻融交替时期的碳氮矿化过程及其相互关系。结果表明,培养温度和培养时间对土壤碳矿化速率和碳矿化累积量均有显著影响。第1次和第5次冻融循环后,冻融处理土壤的碳矿化速率显著高于恒温培养下土壤的碳矿化速率;第7次和第15次冻融循环后,冻融土壤碳矿化累积量显著低于恒温土壤的碳矿化累积量。土壤氮矿化速率没有受到培养温度、培养时间以及二者交互作用的影响,但培养时间和培养温度对土壤净氮矿化累积量有显著的影响。第5、7、15次冻融循环后,冻融处理的土壤无机氮净矿化累积量低于恒温培养的土壤无机氮净矿化累积量。经过30 d的培养,恒温处理下的土壤碳、氮矿化累积量(碳累积量:92.82 μg·g-1,氮累积量73.76 mg·kg-1)是冻融处理下(碳累积量:65.51 μg·g-1,氮累积量33.45 mg·kg-1)的1.42倍和2.21倍。土壤碳矿化累积量与土壤净氮矿化累积量均为正相关关系,但在相同的碳释放量下冻融循环处理土壤累积的无机氮较少。以上结果表明,冻融循环减少了大兴安岭寒温带落叶松林土壤碳排放和无机氮的累积,有利于土壤碳的固持和减少养分的流失。
北方森林;冻融循环;土壤碳释放;铵态氮;硝态氮;微生物生物量
冻融现象是由于季节性或昼夜温度的变化使土壤反复出现冻结-融化的过程,是高纬度地区和山区普遍存在的自然现象(伍星和沈珍瑶,2010)。冻融循环的频率和强度主要取决于气候气温和冬季积雪的厚度。在全球变化的背景下,变暖可能会降低冻融循环的强度、频率和持续时间;而减少的积雪厚度可能会加强冻结的过程(Matzner和Borken,2008)。
冻融循环通过改变土壤的物理性质、水热条件、微生物种类组成和活性,改变土壤的碳氮循环,进而影响土壤可利用碳氮的流失和植物对土壤养分的吸收(Schimel和Clein,1996;Schimel等,2004;Matzner和Borken,2008)。冻融循环对土壤碳氮矿化过程影响的研究是近年来生态学研究的热点问题之一。冻融对土壤碳氮矿化的影响结果是不一致的,有正效应、负效应和没有效应,主要取决于土壤本身的特性和冻融的温度、频率和持续时间。通常认为,在较低的冻结温度下,冻融循环破坏了土壤团聚体的组成结构,促进了土壤可溶性有机碳(DOC)和可溶性有机氮(DON)的释放、土壤胶体解吸附NH4+离子;使部分微生物细胞膜破裂释放有机碳和有机氮;促使根系死亡(Fitzhugh等,2001;Herrmann和Witter,2002;Sharma等,2006)。这些微生物呼吸底物的增加使得土壤碳氮矿化速率增加。而在较高的冻结温度下,冻融循环对碳氮矿化过程的影响取决于不同的土壤特性、植被类型和冻结温度。例如,-5 ℃左右的冻结温度降低了耕地土壤微生物的活性(Herrmann和Witter,2002;Dörsch等,2004),但是没有改变高山和极地土壤的微生物活性(Lipson等,2000;Lipson等,2002;Grogan等,2004)。有可能是高山和极地土壤微生物适应了低温环境,从而对-5 ℃的冻结温度并不敏感。Neilsen等(2001)对糖槭林(Acer saccharum Marsh)和黄桦林(Betula alleghaniensis Britton)土壤培养实验研究表明,相比于对照处理来说,-3 ℃的冻融过程轻微地促进了土壤CO2的排放,没有明显改变氮矿化,并且黄桦林土壤的CO2释放量明显高于糖戚林土壤。对挪威云杉林(picea abies L. Karst)土壤的研究表明,-8 ℃的冻融循环降低了土壤氮矿化量(Hentschel等,2008),增加了CO2排放(Goldberg等,2008)。此外,野外的雪移除实验也表明,雪移除改变的冻融循环有的是通过影响土壤的物理结构促进了土壤无机氮的增加(Freppaz等,2008),有的是通过降低微生物的活性而抑制了无机氮的产生(Tan等,2014)。因此,对于较高冻结温度的冻融循环过程对土壤碳氮矿化过程的影响没有一致性的结论,仍然需要针对不同的土壤进行进一步的研究。
大多数关于冻融循环和土壤碳氮矿化的关系的研究仅仅分别关注了冻融对碳矿化和氮矿化的影响,而对于冻融作用下碳氮矿化关系的分析很少。氮矿化和碳矿化之间的关系很大程度上取决于分解底物的C∶N比和微生物碳利用效率(Murphy等,2003)。添加可利用的碳源或者氮源会改变碳氮矿化之间的关系(Song等,2011)。那么,冻融循环作用下土壤DOC和DON等土壤微生物可利用的碳氮底物的改变,是否会改变土壤碳氮矿化之间的关系,不得而知。本文采用室内培养实验,以大兴安岭兴安落叶松林土壤为研究对象,分析-5~8 ℃冻融和8 ℃恒温条件下土壤碳氮矿化的变化过程以及碳氮矿化的相互关系,有利于深入理解秋末春初冻融作用下的土壤碳氮循环过程。
1 材料和方法
1.1 样地概况
实验样地属于内蒙古根河国家野外科学观测研究站以东的开拉气林场。该林场位于大兴安岭西北坡(50°49′~50°51′N,121°30′~121°31′E),海拔826 m,地势平坦,坡度小于3°。该区域属于寒温带半湿润气候,年均气温-5.4 ℃,年降水量450~550 mm,其中60%集中在6─8月。秋季日最高温月平均值为10.6 ℃,日最低温月平均值为-4.7 ℃;春季日最高温月平均值为6.6 ℃,日最低温月平均值为-11.1 ℃(刘丽和王晓红,2008)。9月末到翌年5月初为降雪期,降雪厚度20~40 cm,降雪量占全年降水总量的12%。全年地表蒸发量800~1200 mm,年均日照2594 h,无霜期80 d。该区为大片连续多年冻土带的南缘,土壤为发育于花岗岩残积物上的暗棕壤,土层厚度30~40 cm,其中矿质层石砾较多。土壤主要理化性质如下:腐殖质含量10%~30%,全氮2.9~4.7 g·kg-1,全磷0.5~1.1 g·kg-1,pH值为4.5~6.5,土壤容重0.15~0.74 g·cm-3(Gao等,2013)。植被类型为杜香-落叶松林,林龄约为200年,主要物种为兴安落叶松(Larix gmelini)、白桦(Betula platyphylla)、杜香(Ledum palustre)、杜鹃(Rhododendron simsii)、红豆越橘(Vaccinium Vitisidaea)等。
1.2 样品采集与处理
我们选择了9个10 m×20 m的样方,每个样方间隔10 m。2013年10月,土壤秋季冻融之前进行样品采集。在每个样方内,去除地表凋落物后,用土钻取0~10 cm土壤样品,6点形成1个混合样品。总计9个土壤样品。取样后过4 mm筛,手工挑除根系和杂质。放入冻干机中进行冻干,以便于保持微生物活性和室内培养过程中调节土壤含水量。
1.3 室内培养及测定
培养实验设置-5~8 ℃冻融循环和8 ℃恒温对照2个温度处理。其中,-5~8 ℃冻融循环处理是先将样品放入-5 ℃冰箱中冻结24 h,后将样品放入8 ℃恒温培养箱中融化24 h,如此为1个冻融循环。8 ℃对照处理设计为将样品放入恒温培养箱中8 ℃恒温培养。土壤含水量统一调节为60%土壤饱和含水量,即含水量为35.5%。土壤饱和含水量采用简易法(代景忠等,2012)测定,3次重复。
培养实验开始前,称取冻干的土壤样品,用小喷壶将去离子水均匀喷至土上,将土壤含水量调节至60%土壤饱和含水量。随后,在150 mL塑料圆瓶中装入33 g左右(干重25 g左右)调节好含水量的土壤样品,用于测定土壤碳矿化速率;另将250 g左右调节好含水量的土壤样品装于塑料方瓶内,用于分次测定土壤NH4+-N和NO3--N含量。所有瓶子盖盖(留有通气孔)在8 ℃培养箱中预培养2 d后,测定土壤碳矿化速率和土壤NH4+-N和NO3--N含量。随后,冻融循环处理的培养样品放入-5 ℃的冰箱中,恒温处理部分的培养样品继续留在8 ℃培养箱中培养。此后在30 d的培养期内,分别在第1、3、5、7和15次冻融循环完成后(即培养的第2、6、10、14和30天)测定土壤碳矿化速率和土壤NH4+-N和NO3--N含量。培养过程中,每隔5 d采用称重法给培养样品补水一次,将去离子水均匀喷洒在土壤表面。
土壤碳矿化速率采用土壤微生物呼吸速率测定系统进行测定,详见代景忠等(2012)。土壤无机氮(NH4+-N,NO3--N)采用0.5 mol·L-1硫酸钾(K2SO4)浸提,含量采用流动化学分析仪(Bran Luebbe,Germany)测定。土壤微生物生物量碳(MBC)采用氯仿熏蒸法(chloroform fumigation-extraction method,FE)测定。称取10 g左右新鲜土样放入50 ml的玻璃烧杯中。烧杯放入真空干燥器中,在真空干燥器底部放入少量用去离子水湿润过的滤纸以保持湿度。另外,放置一个装有30~50 mL氯仿的烧杯,烧杯中放入少量沸石以防止暴沸。真空干燥器密封后用真空泵抽至烧杯中的氯仿沸腾1~2 min,将活塞关闭,将干燥器放入一恒温培养箱中,在25~28 ℃下暗室培养24 h。同时称取10 g的同一份土样,置于50 mL广口塑料瓶中,加入100 mL 0.5 mol·L-1K2SO4,放在振荡器上以180 r·min-1速度振荡1 h,停止振荡后静置30 min,将样品过滤至小塑料瓶中放入冰柜冷冻。熏蒸培养结束后取出真空干燥器,用真空泵反复抽去残余氯仿,土样同上提取保存提取液。浸提液采用TOC仪测定,土壤微生物生物量碳(MBC)计算公式如下:
MBC=(熏蒸后土壤提取液中的有机C-未熏蒸土壤提取液中的有机C)×2.2,单位mg·kg-1(林先贵等,2010)。
1.4 数据处理及统计分析
土壤碳矿化速率采用如下公式进行计算(代景忠等,2012):

式中:R为土壤微生物呼吸速率(µg·g-1·d-1,以C计),C为测试时间内CO2浓度变化的直线斜率,V是培养瓶和管线的总体积,m是培养瓶内土壤干重,α是CO2气体质量转化系数,β是时间转化系数。
同时,我们对第一个冻融循环过程中土壤呼吸进行了详细测定,在48 h中每隔20 min测量一次。经过测定发现,在一次冻融循环中,升温过程和降温过程各持续4个小时。一次冻融循环的全过程分为8~-5 ℃降温(4 h)、-5 ℃恒温(20 h)、-5~8 ℃升温(4 h)和8 ℃恒温(20 h)4个过程。据降温和升温过程中的土壤呼吸速率与土壤温度进行拟合,两者呈指数关系,分别表示为如下公式:
升温:Y = 2.5987e0.0783X
降温:Y = 1.5075e0.0747X
式中:Y为土壤呼吸速率(µg·g-1·d-1),X为土壤温度。
我们假设-5 ℃恒温(20 h)和8 ℃恒温(20 h)过程中土壤呼吸速率呈线性变化。按照如下方法计算每个冻融循环过程中土壤呼吸累积量:利用第1、3、5、7、15冻融循环后实际测量的土壤呼吸速率,分别通过升温和降温公式计算出-5 ℃和8 ℃恒温时的土壤呼吸速率,采用线性内插法计算土壤呼吸累积量。土壤无机氮净矿化累积量(mg·kg-1·d-1,以N计)同样采用线性内插法计算。
利用SPSS16.0统计软件进行统计分析。采用单因变量多因素方差分析(Univariate ANOVAs)方法分析培养温度、培养时间及其交互作用对大兴安岭寒温带针叶林0~10 cm表层土壤碳、氮矿化的影响,采用配对样本T检验方法分析对比-5~8 ℃冻融处理和8 ℃恒温处理土壤碳氮矿化的差异。利用SigmaPlot 10.0软件进行绘图。除特殊说明外,显著性水平ɑ=0.05。
2 结果
2.1 冻融作用下土壤碳矿化的变化
培养温度和培养时间对土壤碳矿化速率均具有显著的影响(P=0.001和P=0.000,表1),且两因素间存在显著的交互效应(P=0.025,表1)。从培养温度上看,总体来说,经过-5~8 ℃冻融循环处理后,土壤在融化阶段的碳矿化速率要高于在8 ℃下恒温培养的土壤,其中在第1次和第5次冻融循环后表现最为明显(P=0.003和P=0.000,图1a)。经过-5~8 ℃冻融循环处理的土壤,在融化阶段总体上表现为随着冻融循环次数的增加,土壤碳矿化速率降低,在5次冻融循环后,土壤碳矿化速率最低(图1a)。在8 ℃恒温培养条件下,随着培养时间的增加,土壤碳矿化速率没有明显的降低或增加趋势。
土壤碳矿化累积量受到培养温度、培养时间以及二者交互作用影响极其显著(P=0.000,表1)。随着培养时间的增加,在第7次和第15次冻融循环后两种处理间土壤碳矿化累积量表现出明显差异(P=0.049和P=0.003,图1b)。8 ℃恒温培养条件下的土壤碳矿化累积量是-5~8 ℃冻融处理下的土壤的1.42倍(图1b)。
2.2 冻融作用下土壤氮矿化的变化
土壤氮矿化速率没有受到培养温度、培养时间以及二者交互作用的显著影响(P=0.281,P=0.236和P=0.186,表1)。在第5次和第15次冻融循环,-5~8 °C冻融处理的土壤氮矿化速率显著低于8 °C恒温土壤(P=0.000和P=0.001)。

表1 温度、时间及其交互作用对不同培养条件下土壤碳氮矿化速率及碳氮矿化累积量影响的单因变量多因素方差分析Table 1 Results of Univariate ANOVAs of the effects of temperature and time on the Soil C and N mineralization rate and accumulated C and N mineralization

图1 土壤碳矿化速率和土壤碳矿化累积量的动态变化Fig. 1 Dynamics of soil C mineralization rate and accumulated C mineralization
培养温度和培养时间则对土壤无机氮的净矿化累积量产生了极其显著的影响(P=0.001和P=0.000,表1)。在8 ℃恒温条件下培养2 d后,土壤氮矿化累积量为-2.04 mg·kg-1,说明在这一阶段土壤中的微生物以对氮的固持为主(图2b);经过6 d的恒温培养,土壤无机氮矿化累积量增加到13.56 mg·kg-1,且在之后的培养过程中随着时间的延长而逐渐增加,经过30 d的培养,土壤无机氮净矿化累积量显著增加(图2b)。冻融循环处理的土壤,经过1次冻融循环后同样出现氮固持现象,其土壤氮矿化累积量为-0.83 mg·kg-1(图2b);随培养时间的增加,土壤氮矿化累积量呈增加趋势。对比8 ℃恒温和-5~8 ℃冻融2种处理,冻融循环处理的土壤在5次、7次和15次循环后,其土壤氮矿化累积量显著低于恒温部分(P=0.004,P=0.001和P=0.000,图2b),可见冻融交替会减少土壤中有效氮的累积。经过15次的冻融循环后,冻融处理土壤的净氮矿化累积量为恒温土壤的45.3%。
2.3 冻融作用下土壤微生物生物量碳的变化
土壤微生物生物量碳在冻融和恒温培养条件下,培养前后没有出现显著差异(P=0.906和P=0.964)。-5~8 ℃冻融处理的土壤微生物生物量碳稍有降低,而8 ℃恒温对照土壤的微生物生物量碳则稍有增加(图3)。
2.4 冻融作用下土壤碳氮矿化的关系
8 ℃恒温培养条件下的土壤碳矿化量与土壤氮矿化量呈正相关关系(图4),即土壤释放的CO2量越多,土壤中的无机氮含量也越高。经过-5~8 ℃冻融循环处理,土壤的碳矿化量与氮矿化量也同样表现出正相关关系(图4)。与8 ℃恒温培养相比,在具有相同的土壤碳排放的情况下,经过冻融的土壤中无机氮的累积量更少。
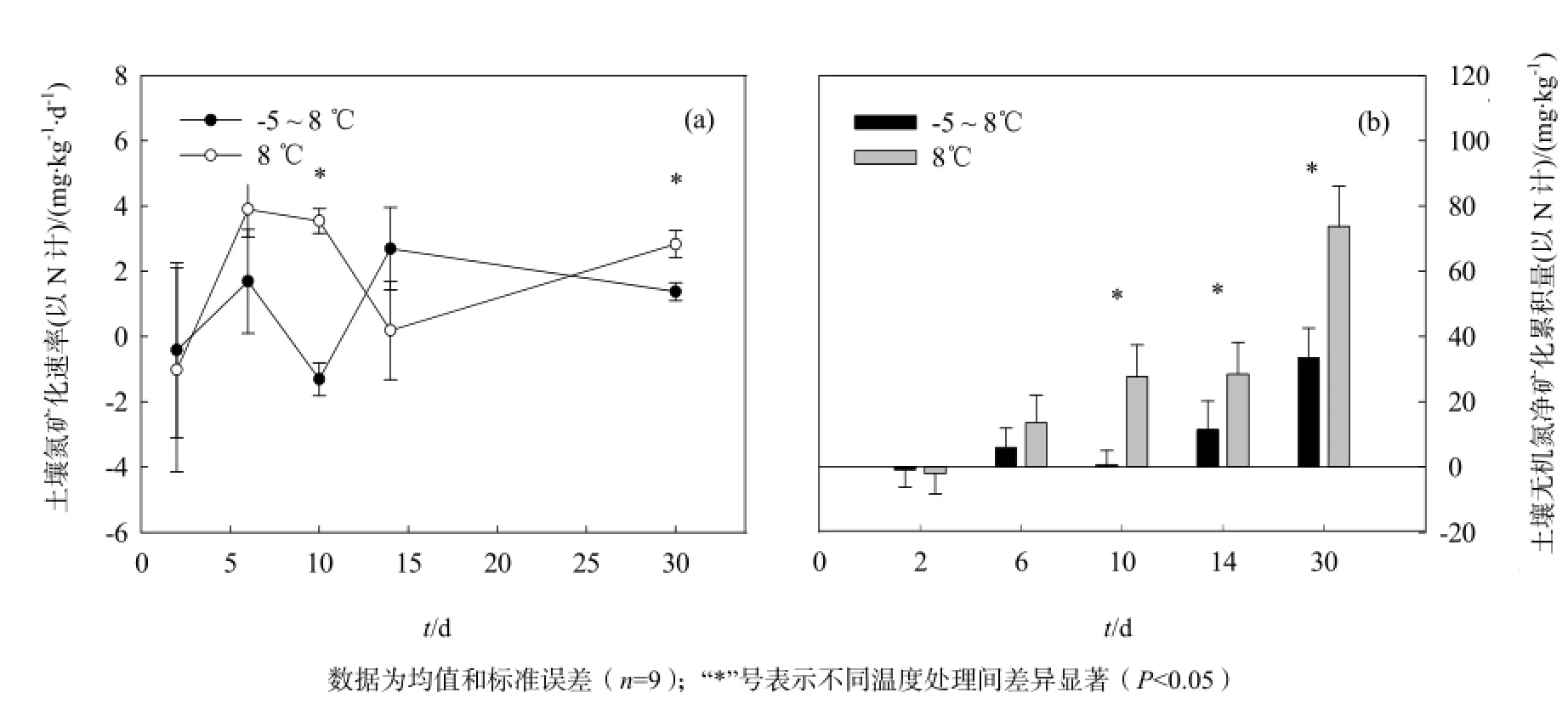
图2 土壤氮矿化速率和氮矿化累积量的动态变化Fig. 2 Dynamics of Soil N mineralization rate and accumulated N mineralization

图3 土壤微生物生物量碳的培养前后的变化Fig. 3 Dynamics of Soil MBC in the 30 d incubation

图4 土壤碳矿化累积量和氮矿化累积量的关系Fig. 4 Relationship of accumulated C mineralization and accumulated N mineralization
3 讨论
3.1 冻融对土壤碳矿化的影响
土壤碳矿化的过程是由微生物主导的。微生物可利用的底物和微生物生长过程都受到温度的影响。冻结促使微生物活性降低,而融化促使微生物活性恢复或增加。我们的研究结果表明,相比于恒温处理,1个冻融循环处理后,微生物呼吸显著增加;随着冻融循环次数的增加,冻融作用对微生物呼吸的刺激效果逐渐消失,2种处理土壤的碳矿化速率均逐渐降低(图1)。我们的这一结果,与前人的研究结果是一致的(贾国晶等,2012)。在冻融初期,连续的冻融循环破坏了土壤团聚体结构,使大团聚体破碎成小团聚体,有利于被土壤团聚体吸附的小分子物质释放(李忠佩和张桃林,2004),增加了土壤可利用碳氮。同时,尽管微生物在低温下仍有活性,-5 ℃的冻结温度也杀死了部分微生物。在融化阶段,死亡微生物可以作为存活微生物的分解底物(Zhou等,2011;Neilsen等,2001)。微生物可利用底物的增加是冻融初期土壤碳矿化速率较高的主要原因。随着冻融循环次数的增加,分解底物逐渐被耗尽,同时,微生物死亡的绝对数量也在减少,使得土壤碳矿化速率逐渐下降。因此,短期来看,冻融促进了碳矿化;而长期来看,由于受到分解底物的限制,两者的碳矿化速率并无显著差异。在土壤碳矿化的累积量上,温度是主要决定因素。相比于恒温处理,冻融循环过程中土壤微生物有一半左右的时间处于低温低活性的状态下。尽管融化的过程中微生物活性得以恢复,冻融处理土壤的总碳释放量仍然低于恒温对照。
3.2 冻融对土壤氮矿化的影响
目前关于冻融交替对土壤氮矿化的影响的研究表明,冻融循环有利于土壤中无机氮的增加(Grogan P等,2003),土壤微生物也是影响土壤氮矿化的重要因素。Marumoto等(1982)研究发现土壤中大部分矿化的N并不是直接来自土壤有机物质,而是由死亡微生物所释放的。冻融过程中部分微生物死亡,使得细胞内物质渗出,成为土壤中矿质氮的直接来源。同时死亡的微生物为土壤中残留微生物提供了有效碳源,刺激了残留微生物的活性,有利于融化阶段土壤氮矿化效率的提高(Herrmann A和Witter E,2002)。此外,冻融会改变土壤的物理结构,温度的变化使得土壤交替收缩和膨胀,导致土壤晶格开放,释放出固定的NH4+(Ferris J C,2000)。
我们的研究结果表明,经过10 d(5次冻融循环)的培养,2种培养条件下的土壤无机氮净矿化累积量差异显著,冻融处理土壤的氮矿化累积量明显低于恒温培养下的土壤,这主要是受到不同培养温度的影响(图2)。在-5 °C的温度处理下,可能微生物没有出现大量地死亡,一是影响了由死亡微生物所释放的矿质氮,同时也影响了能够增加无机氮产生的底物。而在低温下,微生物的活性也有所降低,虽然冻融循环可能使得土壤氮矿化速率在短期内得到提高,但其氮矿化累积量仍要低于在8 ℃恒温培养下的土壤。我们的研究结果与相似冬季温度的其他地区研究结果相近。例如,Neilsen等(2001)对美国新罕布什尔州糖戚林和黄桦林的土壤进行冻融培养实验研究,发现-3 ℃的冻融过程没有明显改变氮矿化,而在更低的温度条件──-13 ℃下,冻融过程促进了氮矿化。
3.3 土壤碳氮矿化的关系
微生物氮矿化的过程,是微生物利用可利用的碳源分解底物获取氮的过程。在分解有机物的同时释放的无机氮一部分被微生物自身固持,一部分被释放到土壤中。土壤碳矿化和氮矿化的关系大部分为正相关关系(Scott等,1998;Parfitt等,2003),即微生物释放CO2的同时也释放无机氮。而在可利用碳氮都十分匮乏的土壤中,如青藏高原的高山草甸(Song等,2011),微生物分解有机物释放的无机氮首先被微生物利用,在满足微生物需求后才释放到土壤中,土壤碳矿化和氮矿化的关系为负相关关系。添加碳源或者氮源,或者改变土壤温度和湿度,都可以使土壤碳氮矿化的关系发生改变(Gao等,2009;Song等,2011)。在冻融条件下,土壤可利用性碳氮源由于土壤物理(团聚体破碎、解吸附等)和生物(温度对微生物和酶的刺激作用)性质的改变而变化,这可能会影响土壤碳氮矿化的关系。我们的研究结果表明,大兴安岭落叶松土壤的碳氮矿化之间是正相关关系,冻融并没有转变这种正相关关系(图4)。但是,在恒温培养和冻融条件下,正相关关系的斜率不同。在相同的碳释放量下,相比于恒温培养,冻融循环减少了土壤无机氮的累积。可能的原因,一是冻融增加了微生物的呼吸;二是冻融促进了微生物氮固持,增加了微生物的氮利用效率;三是微生物种群可能发生了改变。具体原因还需要对冻融过程中土壤总氮矿化的过程、微生物种类的变化以及添加碳源或氮源后土壤碳氮矿化关系的改变等做进一步的细致研究。
4 结论
(1)相比于8 ℃恒温培养,短期的-5~8 ℃冻融循环增加了大兴安岭寒温带针叶林土壤碳矿化速率。但是,长期来看,-5~8 ℃冻融循环下土壤碳释放累积量较8 ℃恒温培养低。
(2)相比于8 ℃恒温培养,冻融循环处理的土壤在5次、7次和15次循环后,其土壤氮矿化累积量显著低于8 ℃恒温培养。冻融交替会减少土壤中的无机氮累积。
(3)8 ℃恒温培养和-5~8 ℃冻融循环处理的土壤,其碳矿化量与土壤氮矿化量均呈正相关关系。但在相同的土壤碳排放的情况下,冻融不利于的土壤中有效氮的累积。
DORSCH P, PALOJARVIA A, MOMMERTZ S. 2004. Overwinter greenhouse gas fluxes in two contrasting agricultural habitats[J]. Nutrient Cycling in Agroecosystems, 70: 117-133.
FITZHUGH R D, DRISCOLL C T, GROFFMAN P M, et al. 2001. Effects of soil freezing disturbance on soil solution nitrogen, phosphorus, and carbon chemistry in a northern hardwood ecosystem[J]. Biogeochemistry, 56: 215-238.
FREPPAZ M, CELI L, MARCHELLI M, et al. 2008. Snow removal and its influence on temperature and N dynamics in alpine soils (Vallée d'Aoste, northwest Italy)[J]. Journal of Plant Nutrition and Soil Science, 171: 672-680.
GAO J Q, OUYANG H, XU X L, et al. 2009. Effects of temperature and water saturation on CO2production and nitrogen mineralization in alpine wetland soils[J]. Pedosphere, 19: 71-77.
GAO W L, CHENG SH L, FANG H J, et al. 2013. Effects of simulated atmospheric nitrogen deposition on inorganic nitrogen content and acidification in a cold-temperate coniferous forest soil[J]. Acta Ecologica Sinica, 33: 114-121.
GOLDBERG S D, MUHR J, BORKEN W, et al. 2008. Fluxes of climate-relevant trace gases between a Norway spruce forest soil and atmosphere during repeated freeze-thaw cycles in mesocosms[J]. Journal of Plant Nutrition and Soil Science, 171: 729-739.
GROGAN P, MICHELSEN A, AMBUS P, et al. 2004. Freeze-thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms[J]. Soil Biology and Biochemistry, 36: 641-654.
HENTSCHEL K, BORKEN W, MATZNER E. 2008. Repeated freeze-thaw events affect leaching losses of nitrogen and dissolved organic matter in a forest soil[J]. Journal of Plant Nutrition and Soil Science-Zeitschrift Fur Pflanzenernahrung Und Bodenkunde, 171: 699-706.
HERRMANN A, WITTER E. 2002. Sources of C and N contributing to the flush in mineralization upon freeze-thaw cycles in soils[J]. Soil Biology & Biochemistry, 34: 1495-1505.
LIPSON DA, SCHADT CW, SCHMIDT SK. 2002. Changes in soil microbial community structure and function in an alpine dry meadow following spring snow melt[J]. Microbial Ecology, 43: 307-314.
LIPSON DA, SCHMIDT SK, MONSON RK. 2000. Carbon availability and temperature control the post-snowmelt decline in alpine soil microbial biomass[J]. Soil Biology & Biochemistry, 32: 441-448.
MATZNER E, BORKEN W. 2008. Do freeze-thaw events enhance C and N losses from soils of different ecosystems? A review[J]. European Journal of Soil Science, 59: 274-284.
MURPHY DV, RECOUS S, STOCKDALE EA, et al. 2003. Gross nitrogen fluxes in soil: theory, measurement and application of15N pool dilution techniques[J]. In: Advances in Agronomy, 79: 69-118.
EILSON C B, GROFFMAN P M, HAMBURG S P, et al. 2001. Freezing effects on carbon and nitrogen cycling in northern hardwood forest soils[J]. Soil Science Society of America Journal, 65: 1723-1730.
PARFITT R L, SCOTT N A, ROSS D J, et al. 2003. Land-use change effects on soil C and N transformations in soils of high N status: comparisons under indigenous forest, pasture and pine plantation[J]. Biogeochemistry, 66: 203-221.
SCHIMEL J P, BILBROUGH C, WELKER J A. 2004. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities[J]. Soil Biology & Biochemistry, 36: 217-227.
SCHIMEL J P, CLEIN J S. 1996. Microbial response to freeze-thaw cycles in tundra and taiga soils[J]. Soil Biology & Biochemistry, 28: 1061-1066.
SCOTT N A, PARFITT R L, ROSS D J, et al. 1998. Carbon and nitrogen transformations in New Zealand plantation forest soils from sites with different N status[J]. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere, 28: 967-976.
SHARMA S, SZELE Z, SCHILLING R, et al. 2006. Influence ofFreeze-Thaw Stress on the Structure and Function of Microbial Communities and Denitrifying Populations in Soil[J]. Applied and Environmental Microbiology, 72: 2148-2154.
SONG M H, JIANG J, XU X L, et al. 2011. Correlation between CO2efflux and net nitrogen mineralization and its response to external C or N supply in an alpine meadow soil[J]. Pedosphere, 21: 666-675.
TAN B, WU F Z, YANG W Q, et al. 2014. Snow removal alters soil microbial biomass and enzyme activity in a Tibetan alpine forest[J]. Applied Soil Ecology, 76: 34-41.
ZHOU W M, CHEN H, ZHOU L, et al. 2011. Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones [J]. Annals of Forest Science, 68: 943-951.
代景忠, 卫智军, 何念鹏, 等. 2012. 封育对羊草草地土壤碳矿化激发效应和温度敏感性的影响[J]. 植物生态学报, 36 (12): 1226-1236.
贾国晶, 周永斌, 代力民, 等. 2012. 冻融对长白山森林土壤碳氮矿化的影响[J]. 生态环境学报, 21(4): 624-628.
李忠佩, 张桃林. 2004. 可溶性有机碳的含量动态及其与土壤有机碳矿化的关系[J]. 土壤学报, 41(4): 544-552.
林先贵, 王一明, 尹睿, 等. 2010. 土壤微生物研究原理与方法[M]. 北京:高等教育出版社: 74-76.
刘丽, 王晓红. 2008. 根河市近47年来气温变化特征[J]. 现代农业科技, 21: 332, 334.
伍星, 沈珍瑶. 2010. 冻融作用对土壤温室气体产生与排放的影响[J]. 生态学杂志, 29: 1432-1439.
Soil Carbon and Nitrogen Mineralization in A Larix gmelinii Forest during Freeze-thaw Cycles
LUO Yachen1,2, LV Yuliang1, YANG Hao2*, HE Nianpeng2, LI Shenggong2, GAO Wenlong2,3
1. School of Geographical Sciences, Southwest University, Chongqing 400715, China; 2. Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China
To investigate the mineralization of soil carbon (C) and nitrogen (N) of cold-temperate coniferous forest soil and their relationships during freeze-thaw cycles, we collected Larix gmelinii forest soil in the Daxinganling region and conducted a 30d laboratory incubation experiment with two treatments of control (constant 8 ℃) and freeze-thaw (each cycle: -5 ℃ in 24 h and then 8 ℃ in 24 h). Our results showed that incubation temperature and time had significant effects on soil C mineralization rates and C mineralization accumulated. After the 1stand 5thfreeze-thaw cycles the soil C mineralization rates were higher in the freeze-thaw treatment than in the control. The amounts of C mineralization accumulated in the freeze-thaw treatment were lower than those in the control after the 7thand 15thfreeze-thaw cycles. Incubation temperature and time had no effect on soil N mineralization rates while the N mineralization accumulated was significantly affected by both two factors. After the 5th, 7th, and 15thfreeze-thaw cycles the amounts of net N mineralization accumulated were higher in the control than in the freeze-thaw treatment. After 30d incubation the amounts of C and N mineralization accumulated were 1.42 and 2.21 times higher respectively in the control (C accumulated:92.82 μg·g-1, N accumulated 73.76 mg·kg-1) as against the freeze-thaw treatment (C accumulated:65.51 μg·g-1, N accumulated 33.45 mg·kg-1). Net N mineralization accumulated was positively correlated with C mineralization accumulated in both control and freeze-thaw treatments. But with the same carbon emission, N mineralization accumulated was less in the freeze-thaw treatment compared with control. Consequently, freeze-thaw cycles can decrease C emission and inorganic N accumulation of the cold-temperate coniferous forest soil in the Daxinganling region, which might improve soil C storage and decrease nutrient loss.
boreal forest; freeze-thaw cycles; soil carbon emission; NH4+; NO3-; microbial biomsss
S718.5
A
1674-5906(2014)11-1769-07
罗亚晨,吕瑜良,杨浩,何念鹏,李胜功,高文龙. 冻融作用下寒温带针叶林土壤碳氮矿化过程研究[J]. 生态环境学报, 2014, 23(11): 1769-1775.
LUO Yachen, LV Yuliang, YANG Hao, HE Nianpeng, LI Shenggong, GAO Wenlong. Soil Carbon and Nitrogen Mineralization in A Larix gmelinii Forest during Freeze-thaw Cycles [J]. Ecology and Environmental Sciences, 2014, 23(11): 1769-1775.
国家自然科学基金项目(31130009;31100336)
罗亚晨(1990年生),女,硕士研究生,从事土地利用与土壤环境研究。Email:xrldxrk1007@163.com
*通信作者:E-mail:yangh@igsnrr.ac.cn
2014-09-18
