DNA中甲基、羟甲基、醛基与羧基胞嘧啶的化学检测方法进展
2014-02-23肖珩,田沺,周翔
肖 珩,田 沺,周 翔
(武汉大学 化学与分子科学学院,湖北 武汉430072)
在表观遗传学上,胞嘧啶转化为5-甲基胞嘧啶(5-methylcytosine,5mC)形式的 DNA 甲基化作为一种特殊的表观遗传修饰,在包括基因表达调控、组蛋白修饰、染色体重组、发育调节和疾病发病机制在内的一系列生命活动中扮演着十分关键的调控作用[1]。维持DNA甲基化和去甲基化过程中DNA甲基化程度的动态平衡对哺乳动物的生长发育至关重要[2]。目前,研究人员在包括癌症在内的许多疾病中均有发现不同程度的DNA甲基化失衡[3]。在细胞分裂过程中,DNA甲基化转移酶家族(DNA methyltransfereases,DNMTs)能够催化未甲基化的胞嘧啶进行甲基化,形成5-甲基胞嘧啶。因此,DNA甲基化转移酶能够影响DNA甲基化的程度。然而,DNA甲基化的状态在细胞中并不是恒定不变的,而是在不同的发展阶段与去甲基化过程保持一个可逆变化。去甲基化现象并非仅仅局限在DNA复制时期[4,5],去甲基化的方式在多年来也存在较多争议。2009年,N.Heintz课题组报道了5-羟甲基胞嘧啶(5-hydroxymethylcytosine,5hmC)存在于哺乳动物的基因组DNA中[6]。与此同时,A.Rao课题组报道了 TET(ten-eleven-translocation,10-11易位)家族的氧化酶能够催化5-甲基胞嘧啶形成5-羟甲基胞嘧啶[7]。羟甲基胞嘧啶作为去甲基化过程中的重要因素开始引起了越来越多的关注。
2011 年,5-醛 基 胞 嘧 啶 (5-Formylcytosine,5fC)和5-羧基胞嘧啶(5-Carboxylcytosine,5caC)作为TET蛋白酶氧化5-甲基胞嘧啶形成的更高级别的氧化产物开始被发现和逐渐了解[8-10]。研究人员发现,TET蛋白酶不仅能够催化甲基胞嘧啶形成羟甲基胞嘧啶,还能将甲基胞嘧啶氧化为醛基胞嘧啶和羧基胞嘧啶的形式,而醛基胞嘧啶和羧基胞嘧啶可以被胸腺嘧啶DNA糖苷酶(thymine DNA glycosylase,TDG)特异性地识别并切割。研究人员不仅在小鼠胚胎干细胞和小鼠器官中发现了醛基胞嘧啶和羧基胞嘧啶,还通过TET蛋白表达调控和TDG酶敲除等实验证实了体内主动去甲基化进程的存在。至此,整个DNA的主动去甲基化机制开始建立并逐步完善(图1)。

图1 DNA甲基化与去甲基化的全过程Process of DNA methylation and demethylation
甲基胞嘧啶、羟甲基胞嘧啶、醛基胞嘧啶和羧基胞嘧啶作为整个DNA甲基化和去甲基化的过程中的重要中间体形式,在整个生命活动中发挥着十分关键的作用。因此,无论是对于基础表观遗传学研究,还是对于包括癌症在内的许多疾病的前期诊断,检测实际样品中这些特殊碱基的分布和含量都具有极为重要的意义,我们曾对甲基化DNA检测的方法作过综述[11]。本文将结合本课题组近些年来在DNA甲基胞嘧啶、羟甲基胞嘧啶、醛基胞嘧啶和羧基胞嘧啶检测方面的工作,小结和综述化学方法检测这些特殊胞嘧啶的最新研究进展。
1 5-甲基胞嘧啶(5mC)的化学检测
在DNA甲基化的检测工作中,亚硫酸氢钠法发挥着举足轻重的作用。研究人员发现[12,13],在经过亚硫酸氢钠磺化、水解脱去氨基和碱化去除亚硫酸根离子等一系列过程之后,正常的胞嘧啶会很快转化为尿嘧啶(uridine,U),而5-甲基胞嘧啶虽然也能够相应地转化为胸腺嘧啶(thymidine,T),但反应速率却远远低于前者(图2)。亚硫酸氢钠法测序(bisulfite sequencing,BS-Seq)(图3)正是利用两者在反应速率上的差异,通过对亚硫酸氢钠法处理后的样品进行PCR扩增,达到准确区分胞嘧啶和甲基胞嘧啶的目的。时至今日,亚硫酸氢钠法测序已成为一种十分普及的检测5-甲基胞嘧啶的方法,并已达到全基因组测序的水平[14,15]。

尽管亚硫酸氢钠法测序已作为一种检测甲基胞嘧啶的有效方法被广泛使用,但其本身依然存在着操作复杂和成本高昂的缺点。因此,越来越多的研究人员开始寻求利用更方便快捷的化学方法来解决甲基胞嘧啶检测的问题。为了解决DNA序列中5-甲基胞嘧啶的位点检测以及定量检测的问题,我们设计并合成了两种N-卤代-硝基苯磺酰胺钠盐的化合物(化合物1和2),用于DNA序列中甲基胞嘧啶的检测[17]。
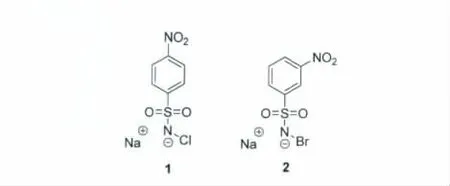
图4 N-卤代-硝基苯磺酰胺钠盐的化合物1和2的结构式[17]Structures of the compound 1 and 2
由于5-甲基胞嘧啶和胞嘧啶的嘧啶环上双键的亲核性以及空间位阻上存在着差异,本课题组选取了两种亲核试剂:N-氯代-硝基苯磺酰胺钠盐和N-溴代-硝基苯磺酰胺钠盐(化合物1和2)。这两种化合物均可以进攻胞嘧啶嘧啶环上的双键,形成卤代胞嘧啶。但是只有N-溴代-硝基苯磺酰胺钠盐(化合物2)可以进攻5-甲基胞嘧啶,形成溴代甲基胞嘧啶(图5)。
利用这两种化合物对5-甲基胞嘧啶和胞嘧啶亲核进攻的差异性,通过结合聚丙烯酰胺电泳实验,便可以准确地检测DNA序列中5-甲基胞嘧啶的数量和分布。相比于传统的亚硫酸氢钠法,该方法操作简便快捷、成本低廉、毒性低。同时,该方法克服了许多化学检测方法存在的不能同时检测DNA序列中多个5-甲基胞嘧啶残基分布的问题,为5-甲基胞嘧啶的检测提供了一种简单可靠的方法。此研究对于寻找更有效检测5-甲基胞嘧啶的化学方法具有重要的参考价值。

图5 5-甲基胞嘧啶检测的路线图Process of 5mC detection
除此之外,通过利用β-糖基转移酶(β-glucosyl transferase,βGT)将糖环转移到DNA序列中的5-羟甲基胞嘧啶的羟基上,进而增加5-羟甲基胞嘧啶的空间位阻,可以成功地区分5-羟甲基胞嘧啶和5-甲基胞嘧啶。这也从另一方面验证了先前对实验中反应机理的推断,在糖基转移酶对5-羟甲基胞嘧啶进行作用之后,5位上的羟基由于被糖环所保护,空间位阻增大导致在随后的反应中无法形成溴鎓离子中间体。因此,该方法还解决了包括亚硫酸氢钠法在内的许多传统方法对DNA链上5-羟甲基胞嘧啶和5-甲基胞嘧啶难以辨别的问题,有效地排除了5-甲基胞嘧啶检测中5-羟甲基胞嘧啶的干扰。为人们深入了解胞嘧啶与5-甲基胞嘧啶在化学结构和性质上的差异,进而通过化学手段检测5-甲基胞嘧啶提供了一种新的思路。
尿嘧 啶-DNA 糖 基 化 酶(Uracil-DNA glycosylase,UDG)能够选择性地去除DNA上的尿嘧啶,进而阻止突变发生。整个过程包括N-糖苷键的切割和碱基修复的启动(base-excision repair,BER)[18]。本课题组的研究人员利用UDG酶的这一特点,再结合亚硫酸氢钠法,设计出了一种能够有效识别DNA链上5-甲基胞嘧啶位点的方法[19]。首先,通过亚硫酸氢钠法将未甲基化的胞嘧啶C脱氨基化,转化为尿嘧啶U,再通过UDG酶将DNA链上的尿嘧啶切除,形成嘧啶缺失的位点(apyrimidinic site,AP位点),然后在碱性条件下处理,DNA链上碱基缺失的位点会被切割(图6)。与此同时,5-甲基胞嘧啶所在的位点依然维持不变,不能被UDG酶切割。因此,通过聚丙烯酰胺电泳实验对比即可检测出5-甲基胞嘧啶的位点。
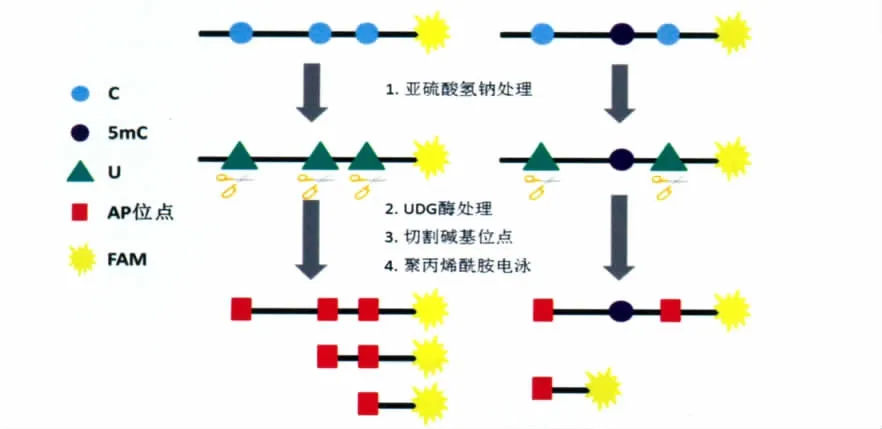
图6 利用亚硫酸氢钠法和UDG酶检测5-甲基胞嘧啶的路线图Combined bisulfite UDG assay for 5mC detection
该方法将UDG酶与亚硫酸氢钠法结合起来,将UDG酶运用到DNA链上5-甲基胞嘧啶分布位点的检测中,取得了准确可靠的结果。该研究为表观遗传学上5-甲基胞嘧啶的检测提供了新的视角,并有望在将来运用到基因组DNA中5-甲基胞嘧啶的分布位置及含量的检测中。
5-甲基胞嘧啶作为哺乳动物基因组中除腺嘌呤(adenine)、胸腺嘧啶(thymine)、胞嘧啶(cytosine)及鸟嘌呤(guanine)之外的第五种碱基,在基因表达调控中发挥着非常重要的作用。由于5-甲基胞嘧啶与胞嘧啶在结构和性质上高度相似,对于基因组中5-甲基胞嘧啶的检测工作存在着许多困难。化学方法由于可控性和选择性强的特点,能够通过合适的化学反应有效地区分5-甲基胞嘧啶和胞嘧啶。相信在未来的研究中,会有越来越多的化学方法被运用到5-甲基胞嘧啶的检测工作中。
2 5-羟甲基胞嘧啶(5hmC)的化学检测
在整个TET系列蛋白酶调控的5-甲基胞嘧啶的氧化过程中,5-羟甲基胞嘧啶(5hmC)扮演着重要的角色。近年,已经陆续有报道在神经元细胞和胚胎干细胞中发现了5-羟甲基胞嘧啶[8-10]。越来越多的资料显示,5-羟甲基胞嘧啶在表观遗传学上具有十分重要的作用,5-羟甲基胞嘧啶的分布和含量已经开始被作为许多癌症疾病诊断的重要标志之一[20,21]。因此,5-羟甲基胞嘧啶的检测逐渐成为表观遗传学上的一大热点。
然而,伴随着5-羟甲基胞嘧啶被发现和认知,一个新的问题开始显现出来:传统的亚硫酸氢钠法测序无法区分5-甲基胞嘧啶和5-羟甲基胞嘧啶[22,23]。Hayatsu课题组曾 经报道[24],在 经过亚硫酸氢钠处理之后,5-羟甲基胞嘧啶(5hmC)会转变为5-亚甲基磺酸盐形式的胞嘧啶(cytosine 5-methylenesulfonate,CMS)(图7),这种磺酸盐形式的胞嘧啶的脱氨基速率比甲基胞嘧啶更慢。在经过PCR信号扩增之后,5-羟甲基胞嘧啶位置的测序结果也会显示为胞嘧啶。因此,传统的亚硫酸氢钠法测序并不能完整地反映基因组中各种碱基的真实分布情况,人们迫切需要寻找新的方法来解决5-羟甲基胞嘧啶检测的问题。
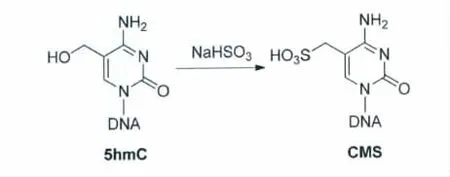
图7 亚硫酸氢钠法处理5-羟甲基胞嘧啶[24]Bisulfite treatment of 5hmC
He Chuan课题 组 发 现[25],和5-甲 基 胞嘧 啶一样,5-羧基胞嘧啶(5-Carboxylcytosine,5caC)在亚硫酸氢钠法测序中会显示为胸腺嘧啶T。因此,研究人员通过利用β-糖基转移酶(β-glucosyl transferase,βGT)将糖环转移到5-羟甲基胞嘧啶的5位羟甲基上,将5-羟甲基胞嘧啶保护起来,然后利用过量的TET氧化酶将5-甲基胞嘧啶氧化为5-羧基胞嘧啶。在经过β-糖基转移酶和TET氧化酶处理之后,在最终的亚硫酸氢钠法测序结果中,5-甲基胞嘧啶处被显示为胸腺嘧啶T,而5-羟甲基胞嘧啶被显示为胞嘧啶C。这样,通过与原先的亚硫酸氢钠法测序做对比,即可识别出5-甲基胞嘧啶和5-羟甲基胞嘧啶残基的分布位点。这种TET协同的亚硫酸氢钠法测序(Tet-assisted bisulfite sequencing,TAB-Seq)有效解决了传统方法无法区分5-甲基胞嘧啶和5-羟甲基胞嘧啶的问题,并达到了基因组中5-羟甲基胞嘧啶测序的水平,为实际样本中5-羟甲基胞嘧啶的检测提供了一种非常可靠的方法。
虽然酶具有高选择性和高反应活性等众多优点,但其本身存在的难以表达及成本过高的问题限制了其使用范围,许多研究人员开始寻求合适的化学方法来检测5-羟甲基胞嘧 啶 。2 0 1 2 年 ,Balasubramanian课 题 组 发 现KRuO4可以选择性地将5-羟甲基胞嘧啶氧化为5-醛 基 胞 嘧 啶(5fC),且 具 有 很 高 的 产率[26]。经 过 KRuO4氧 化 处 理 后,5-羟 甲 基 胞嘧啶会转化为5-醛基胞嘧啶,在最后的测序结果中显示为胸腺嘧啶T,而5-甲基胞嘧啶则显示为胞嘧啶C。这种氧化亚硫酸氢钠测序 (oxidative bisulfite sequencing,oxBS-Seq)不仅成功地解决了5-羟甲基胞嘧啶检测的问题,还为化学手段检测DNA甲基化分布提供了新的思路。KRuO4选择性氧化5-羟甲基胞嘧啶成为5-醛基胞嘧啶这一特性被广泛运用到许多检测方法中。
近些年来,研究人员发明了许多检测5-羟甲基胞嘧啶的方法,包括糖基转移酶实验[27-29]、过氧钨酸盐氧化[30]、基于TET蛋白的亚硫酸氢钠测序[25]和基于高钌酸钾氧化的亚硫酸氢钠测序[26]等。然而,这些方法均需要酶催化或者重金属离子作用,难以避免会引起成本过高、操作复杂以及重金属带来的高毒性的问题,人们开始寻求一些低成本和低毒性的检测方法。
我们设计并合成了一类带有苯甲醛结构的探针,用于检测5-羟甲基胞嘧啶[31]。考虑到5-羟甲基胞嘧啶在嘧啶环上同时拥有羟甲基(C5-CH2OH)和氨基(C4-NH2),研究人员发现拥有苯甲醛的化合物能够与5-羟甲基胞嘧啶反应生成带有1,3-O,N-杂环结构的化合物。因此,通过设计同时带有苯甲醛结构和荧光基团的探针(化合物3和4),可以选择性地与5-羟甲基胞嘧啶反应(图8),并伴随有荧光信号的相应,从而达到特异性检测5-羟甲基胞嘧啶的目的。该方法为化学手段解决5-羟甲基胞嘧啶检测的问题提供了一种崭新的思路,且具有廉价易得、操作方便、毒性低和选择性高等优点,在未来5-羟甲基胞嘧啶的检测工作中具有潜在的应用。但是,令人遗憾的是,研究人员并没有将上述方法进一步运用到对DNA链上的5-羟甲基胞嘧啶以及实际样品中的5-羟甲基胞嘧啶的检测中。

图8 带有苯甲醛结构的探针与5-羟甲基胞嘧啶反应的路线图Reaction of 5hmC with benzaldehyde derivatives probes
由于5-羟甲基胞嘧啶在表观遗传学上的关键作用,基因组DNA中的5-羟甲基胞嘧啶分布的定量检测工作显得尤为重要。Balasubramanian课题组曾发现KRuO4能选择性地氧化5-羟甲基胞嘧啶为5-醛基胞嘧啶[26]。相对于5-羟甲基胞嘧啶,5-醛基胞嘧啶具有更强的反应活性。因此,本课题组研究人员利用KRuO4将5-羟甲基胞嘧啶定量地氧化为5-醛基胞嘧啶,再通过加入羟胺类荧光探针对氧化形成的5-醛基胞嘧啶进行荧光标记。最后,在整个反应体系中加入阳离子共轭聚合物(CCP),使羟胺荧光基团与阳离子共轭聚合物(CCP)之间形成FRET效应(荧光能量共振转移效应)。由于阳离子共轭聚合物(CCP)具有光捕获效应(light harvesting)强、Stokes位移大和视觉信号增强等优点,羟胺荧光基团与阳离子共轭聚合物(CCP)之间形成的FRET效应能够使荧光基团产生的荧光信号大大增强。因此,本课题组通过结合KRuO4氧化、荧光探针标记、阳离子共轭聚合物的FRET效应,成功设计出一种能够在基因组DNA中定量地检测5-羟甲基胞嘧啶含量的方法(图9)。该方法能够成功检测出实际样品中基因组DNA中的5-羟甲基胞嘧啶的含量,为基因组DNA中的5-羟甲基胞嘧啶分布的定量检测工作提供了一种简单有效的新方法,为实际样品中5-羟甲基胞嘧啶的检测提供了更多的选择。同时,该方法还为许多相关工作的实验设计提供了重要的参考依据。
在DNA甲基化的研究中,5-甲基胞嘧啶和5-羟甲基胞嘧啶的检测引起了广泛的关注。与此同时,5-羟甲基尿嘧啶(5-hmdU)作为 DNA 氧化损伤的形式之一,同样引起了研究人员极大的兴趣[33,34]。Zora课题组曾经发现,血液中5-羟甲基尿嘧啶的含量与乳腺癌密切相关,可以作为乳腺癌检测的标志物[35]。然而,目前对5-羟甲基尿嘧啶检测的研究工作还十分有限[36]。
我们报道了一种检测5-羟甲基尿嘧啶的简单有效的方法[37]。研究人员发现,在三氟乙酸的作用下,5-羟甲基尿嘧啶C-5位上的羟基能够选择性地与叠氮化钠作用,生成5-叠氮甲基尿嘧啶(5-aedU)。利用这一特性,通过加入带炔基的荧光探针,可以与5-叠氮甲基尿嘧啶上的叠氮发生click反应,产生荧光相应,进而达到特异性地检测5-羟甲基尿嘧啶的目的。值得注意的是,其他碱基在三氟乙酸存在的条件下均不与叠氮化钠作用,包括5-羟甲基胞嘧啶。
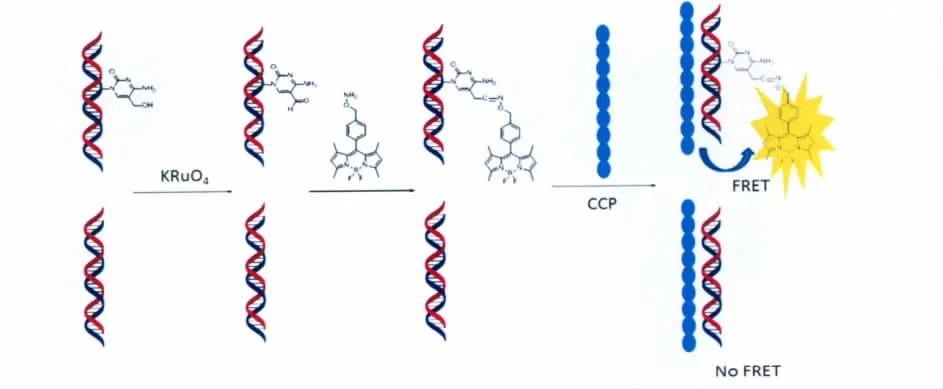
图9 基因组中5-羟甲基胞嘧啶定量检测的路线图Process of genomic 5fC quantitative detection
因此,我们设计了一种仅通过一步取代反应即可以将5-羟甲基尿嘧啶上的羟基替换为叠氮,然后利用带炔基的荧光探针与叠氮发生click反应来对5-羟甲基尿嘧啶进行荧光检测(图10)。该方法相比于传统的检测方法,具有方便快捷、成本低廉和选择性高的优点,对化学手段检测5-羟甲基尿嘧啶的工作具有重要的参考价值,并有望在将来运用到实际样品中5-羟甲基尿嘧啶的检测工作中。
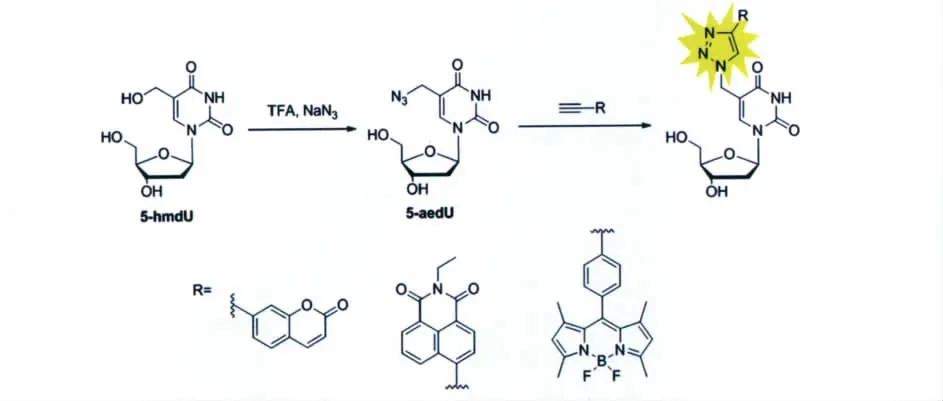
图10 通过click反应对5-羟甲基尿嘧啶(5-hmdU)进行荧光检测的反应路线图Schematic illustration of fluorescence turn-on detection of 5-hmdU based on click reaction
3 5-醛基胞嘧啶(5fC)和5-羧基胞嘧啶(5caC)的化学检测
作为TET酶蛋白调控的5-甲基胞嘧啶氧化过程中的产物,5-醛基胞嘧啶(5fc)和5-羧基胞嘧啶(5caC)在整个DNA的去甲基化过程中扮演着非常关键的角色。研究人员已经在胚胎干细胞中检测到了5-醛基胞嘧啶和5-羧基胞嘧啶的存在,并发现5-醛基胞嘧啶对干细胞分化过程具有潜在的作用[8-10]。因此,5-醛基胞嘧啶和5-羧基胞嘧啶的检测在遗传生物学上具有十分重要的意义。
传统的检测DNA损伤的方法包括聚合酶链式反应(PCR)、基因测序和基因芯片,但以上方法均存在成本高昂和操作复杂的问题。因此,由于化学反应具有高选择性、高灵敏度、操作方便和对DNA低损害等优点,许多课题组开始考虑通过化学的方法设计合成小分子探针来实现对损伤DNA的快速灵敏检测。然而,目前已知的许多检测方法都存在高毒性以及需要特殊目标DNA序列、对检测目标不具有普适性的缺陷[30,38-40]。
Carell课题组经过系统的研究,发现5-醛基胞嘧啶上醛基的氧原子能够与嘧啶环上的氨基形成分子内氢键,致使5-醛基胞嘧啶上的醛基难以反应[41]。该课题组设计了一种带羟胺基团的生物素探针,通过羟胺基团与5-醛基胞嘧啶选择性反应达到检测5-醛基胞嘧啶的目的[10]。Balasubramian课题组利用这一方法,对基因组中DNA的醛基胞嘧啶进行了测序[42]。
众所周知,氨基能够与醛基反应生成西氟碱,因此带有氨基的荧光探针可以用于对5-醛基胞嘧啶进行特异性标记。然而,由于分子内氢键的形成,5-醛基胞嘧啶上的醛基的反应活性较普通醛基要低,许多氨基基团不能与5-醛基胞嘧啶上的醛基反应[41]。
我们通过一系列的研究发现[43],与芳环直接相连的氨基很难与5-醛基胞嘧啶的醛基反应。与此同时,烷基链上的氨基对5-醛基胞嘧啶上的醛基具有很高的反应活性,可以与5-醛基胞嘧啶上的醛基特异性地反应。基于此,研究人员设计并合成了一类高反应活性的氨基荧光化合物5和6,用于5-醛基胞嘧啶上的醛基的检测(图11,图12)。该反应在室温下即可顺利进行,拥有很高的产率,并且不需要任何催化剂。

图11 氨基荧光化合物5和6的结构式Structures of the compound 5 and 6
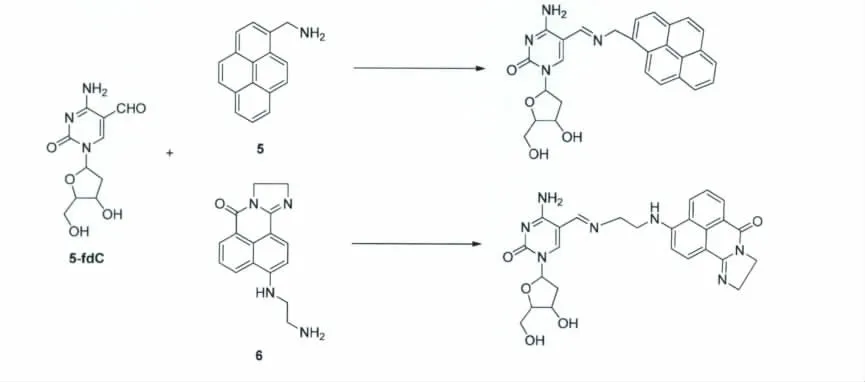
图12 氨基荧光探针与5-醛基胞嘧啶反应的路线图Reaction of 5fC with amine probes
我们通过设计并合成氨基荧光化合物5和6,成功实现了与5-醛基胞嘧啶上醛基的反应,解决了5-醛基胞嘧啶上的醛基具有惰性、难以反应的问题。除此之外,研究人员还通过高效液相色谱、聚丙烯酰胺电泳和荧光检测实验证实了该氨基荧光探针可以对DNA链上的5-醛基胞嘧啶进行了荧光标记,从而达到特异性地检测5-醛基胞嘧啶的目的。此外,该研究对于相关检测探针的设计也具有重要的参考价值。令人遗憾的是,该工作并未选取实际样品进行检测,缺乏能够检测基因组中5-醛基胞嘧啶的足够证据。
为了更充分地证明小分子荧光探针能够用于基因组中5-醛基胞嘧啶的检测,我们设计并合成了一类羟胺荧光化合物7~9(图13)[44],通过对5-醛基胞嘧啶进行荧光标记(图14),进而达到检测5-醛基胞嘧啶的目的。研究人员通过高效液相色谱(HPLC)、聚丙烯酰胺电泳(PAGE)以及 DNA质谱等方法证实了设计合成的羟胺荧光探针特异性地与DNA链上的5-醛基胞嘧啶发生了反应,并且拥有很高的反应产率。除此之外,研究人员还特意选取了小鼠胚胎干细胞作为实际样品,利用所合成的羟胺荧光探针对小鼠胚胎干细胞中的5-醛基胞嘧啶进行了检测,并通过液相色谱-质谱(LC-MS)对这一结果进行了证实。因此,可以推断该羟胺荧光探针可以用于基因组中5-醛基胞嘧啶分布位点和含量的检测。
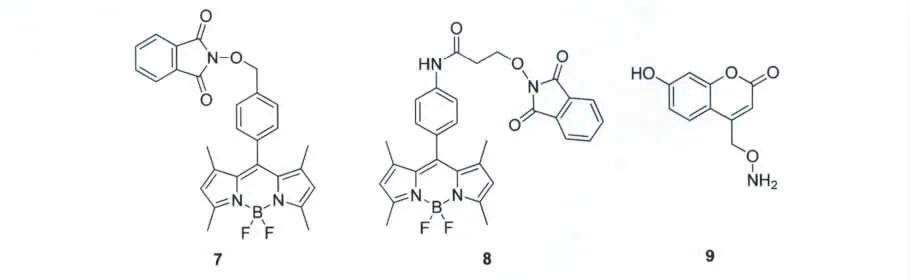
图13 荧光羟胺类化合物7~9的结构式Structures of fluorescent hydroxylamine probes 7-9
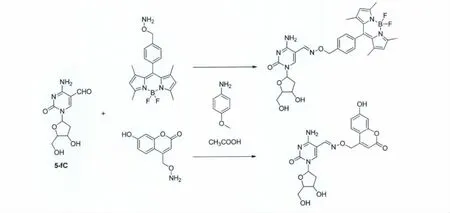
图14 羟胺类荧光探针与5-醛基胞嘧啶反应的路线图Reaction of 5fC with hydroxylamine probes
以上两种方法利用了胺基和羟胺基团与5-醛基胞嘧啶中醛基反应的高活性和高选择性,并且通过标记荧光基团的方法,具有荧光检测的灵敏高效性和简洁性。因此,该方法克服了检测5-醛基胞嘧啶的传统方法操作复杂、花费昂贵的问题,具有化学方法操作简便、成本低廉、高灵敏度和选择性等优点,并且达到了在基因组中检测5-醛基胞嘧啶的水平,为检测DNA序列中5-醛基胞嘧啶的分布位点和含量提供了一种灵敏快捷的新方法,为相关疾病的前期诊断提供了一种可靠的方法。除此之外,对5-醛基胞嘧啶反应活性的系统研究也为将来通过进行化学反应来进行选择性识别5-醛基胞嘧啶的工作提供了许多新的思路,以化学小分子作为探针的研究手段有望在将来直接用于体内5-醛基胞嘧啶含量的检测。
在DNA去甲基化过程中,5-醛基胞嘧啶和5-羧基胞嘧啶作为胞嘧啶被TET酶氧化后更高级别的氧化形式,能够被胸腺嘧啶DNA糖苷酶(thymine DNA glycosylase,TDG)特异性地识别并切割[9]。然而,5-醛基胞嘧啶和5-羧基胞嘧啶在去甲基化过程中的确切功能和机理还存在着广泛的争议[10,42]。
我们发现,经过热哌啶处理,DNA链上的5-醛基胞嘧啶位点会特异性地断裂,且拥有很高的切割效率,而经过相同条件处理的5-甲基胞嘧啶和5-羟甲基胞嘧啶处并不会被切割(图15)[45]。研究人员利用这一特点,设计出了基于热哌啶处理选择性地识别DNA链上5-醛基胞嘧啶分布位点的方法,并通过聚丙烯酰胺电泳和高效液相色谱(HPLC)实验进行了证实。除此之外,基于KRuO4能够选择性地氧化5-羟甲基胞嘧啶为5-醛基胞嘧啶这一特性,研究人员还通过结合KRuO4氧化和热哌啶处理方法来达到检测DNA链上的5-羟甲基胞嘧啶分布的目的(图16)。
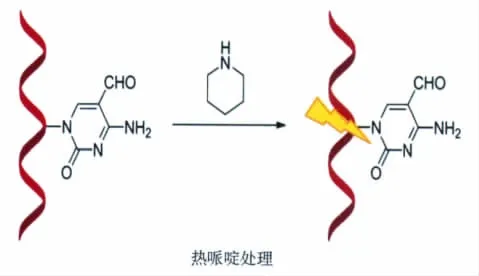
图15 通过热哌啶处理识别DNA序列中的5-醛基胞嘧啶Treatment of the DNA sequences containing 5fC with hot piperidine
该方法能够在没有TDG酶作用的情况下有效地识别DNA链上的5-醛基胞嘧啶和5-羟甲基胞嘧啶,利用化学方法成功检测出DNA序列中5-醛基胞嘧啶和5-羟甲基胞嘧啶分布位点,并且为研究人员深入了解5-醛基胞嘧啶的特有结构和性质,进而找到更多合适的化学方法检测5-醛基胞嘧啶提供了新的思路。
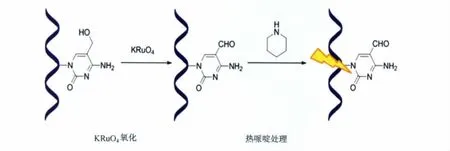
图16 通过KRuO4氧化和热哌啶处理识别DNA序列中的5-羟甲基胞嘧啶Treatment of the DNA sequences containing 5fC with KRuO4and then hot piperidine
随后,本课题组研究人员通过对热碱性条件处理后DNA序列中的5-甲基胞嘧啶、5-羟甲基胞嘧啶、5-醛基胞嘧啶和5-羧基胞嘧啶位点变化的系统研究[46],发现在95℃,250mmol/L NaOH 条件下处理15min,DNA链上5-醛基胞嘧啶位点即可被完全切割(图17)。同时,研究人员还发现5-羧基胞嘧啶位点也会被切割,但切割效率弱于5-醛基胞嘧啶。
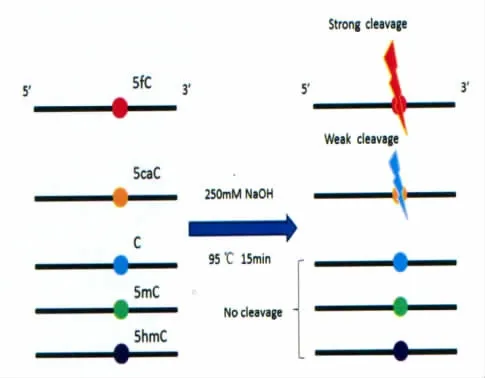
图17 热碱性条件对不同胞嘧啶的选择性切割Illustration of the excision behaviors of DNAs with modified cytosines by hot alkali treatment
通过聚丙烯酰胺电泳实验,针对热碱性条件处理对DNA序列中的5-甲基胞嘧啶、5-羟甲基胞嘧啶、5-醛基胞嘧啶和5-羧基胞嘧啶位点的影响进行了系统研究,发现热碱性条件能选择性切割DNA链上的5-醛基胞嘧啶和5-羧基胞嘧啶位点,且对5-醛基胞嘧啶的切割效率高于5-羧基胞嘧啶。对此特殊现象,研究人员将其归因于5-醛基胞嘧啶和5-羧基胞嘧啶上5位的醛基和羧基的拉电子效应。由于热碱容易配置且处理方便,该方法可以作为Maxam-Gilbert DANN测序法的有效补充,同时能给未来的表观遗传分析工作提供一种非常有效的策略,对DNA去甲基化过程中各种胞嘧啶的性质和机理研究具有重要的参考价值。
除此之外,Bifeng Yuan和我们课题组共同通过将穿梭载体技术与下一代测序技术相结合,系统地研究了胞嘧啶甲基化的衍生物(5hmC、5fC和5caC)对DNA复制过程的影响[47](图18)。统计结果显示,在大肠杆菌中,尽管对DNA复制效率没有影响,所有的胞嘧啶衍生物都能诱导一定程度上(0.17%~1.12%)的胞嘧啶C到胸腺嘧啶T的转录突变,这可能是基因组引导细胞分化的潜在原因之一。这为DNA去甲基化过程中各种甲基胞嘧啶氧化产物的潜在突变特性提供了新的有力证据,为人们深入了解这些特殊碱基的生物学功能提供了新的视角。

图18 对甲基胞嘧啶氧化产物突变和复制性质的系统研究[47]Systematic investigation of mutagenic and cytotoxic properties of oxidation products of 5-methylcytosine
甲基胞嘧啶、羟甲基胞嘧啶、醛基胞嘧啶和羧基胞嘧啶作为DNA甲基化和去甲基化过程中的重要组分,这些特殊碱基在基因组中的分布和含量对整个生命过程具有重要的影响。由于各种碱基的结构和性质方面存在着相似性与差异性,寻找合适的化学方法可以有效地区分这些特殊的碱基,有利于DNA甲基化和去甲基化过程的本质机制研究,并能为甲基化检测的研究带来许多帮助。
4 展望
DNA甲基化作为重要的表观遗传学修饰,在哺乳动物的生长发育过程中起着极为关键的作用。DNA甲基化和去甲基化的动态平衡影响着许多生命活动的正常进行,而这种动态平衡与甲基胞嘧啶、羟甲基胞嘧啶、醛基胞嘧啶和羧基胞嘧啶这些DNA甲基化和去甲基化过程中重要的中间体形式的分布含量密切相关。检测基因组中这些天然修饰的胞嘧啶的分布位置及相对含量对从表观遗传学研究到临床疾病诊断的各种生物医学研究工作都有着非常重要的意义。在对于这些特殊胞嘧啶的检测工作中,研究人员已经取得了许多激动人心的进展。化学方法因其具有方便快捷、成本低廉、可修饰性和可控性强等优点,在整个DNA甲基化的检测过程中发挥着重要的作用。同时,由于这些特殊的胞嘧啶之间存在着密切的联系,化学方法有利于更加深入地认识和了解这些碱基的结构、性质和功能,为表观遗传检测提供了许多帮助和启发。同时,由于生物体的复杂性,还有许多对DNA甲基化检测和与DNA甲基化有关的生命活动机制有待于进一步深入的研究。对于研究DNA甲基化的相关工作,还有赖于包括化学、生物、医学和物理学在内的多学科的共同协作,今后的研究将更加侧重于了解这些特殊碱基确切的角色及其潜在的机理。毫无疑问,化学方法将会在未来的DNA甲基化研究工作中发挥十分重要的作用。致谢:该项目得到国家重点基础研究发展计划(973)(2012CB720600,2012CB720603),教育部“长江学者创新团队”(IRT1030)和“111”引智计划的资助。
[1] Klose R J,Bird A P.Genomic DNA methylation:the mark and its mediators[J].Trends in Biochemical Sciences,2006,31(2):89-97.
[2] Smith Z D,Meissner A.DNA methylation:roles in mammalian development[J].Nature Reviews Genetics,2013,14:204-220.
[3] Baylin S B,Jones P A.A decade of exploring the cancer epigenome-biological and translational implications[J].Nature Reviews Cancer,2011,11(10):726-734.
[4] Wu S C,Zhang Y.Active DNA demethylation:many roads lead to rome[J].Nature Reviews Molecular Cell Biology,2010,11(9):607-620.
[5] Bhutani N,Burns D M,Blau H M.DNA demethylation dynamics[J].Cell,2011,146(6):866-872.
[6] Kriaucionis S,Heintz N.The nuclear DNA base 5-hydroxymethylcytosine is present in purkinje neurons and the brain[J].Science,2009,324(5929):929-930.
[7] Tahiliani M,Koh K P,Shen Y,et al.Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1[J].Science,2009,324(5929):930-935.
[8] Ito S,Shen L,Dai Q,et al.Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine[J].Science,2011,333(6047):1300-1303.
[9] He Y F,Li B Z,Li Z,et al.Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA[J].Science,2011,333(6047):1303-1307.
[10] Pfaffeneder T,Hackner B,Truß M,et al.The discovery of 5-formylcytosine in embryonic stem cell DNA[J].Angewandte Chemie International Edition,2011,50:7008-7012.
[11] Tian T,Wang S R,Wu J G,et al.Review:advances in methodology of DNA methylation assay[J].Science China Chemistry,2011,54(8):1233-1243.
[12] R Shapiro,R E Servis,M Welcher,Reactions of uracil and cytosine derivatives with sodium bisulfite[J].Journal of the American Chemical Society,1970,92:422-424.
[13] H Hayatsu,Y Wataya,K.Kai,Addition of sodium bisulfite to uracil and to cytosine[J].Journal of the American Chemical Society,1970,92:724-726.
[14] Lister R,Pelizzola M,Dowen R H,et al.Human DNA methylomes at base resolution show widespread epigenomic differences[J].Nature,2009,462(7271):315-322.
[15] Balasubramanian S.Decoding genomes at high speed:implications for science and medicine[J].Angewandte Chemie International Edition,2011,50(52):12406-12410.
[16] Schüler P,Miller A K.Sequencing the sixth base (5-hydroxymethylcytosine):selective DNA oxidation enables base-pair resolution[J].Angewandte Chemie International Edition,2012,51(43):10704-10707.
[17] Wang T,Hong T,Tang T,et al.Application of N-halogeno-N-sodiobenzenesulfonamide reagents to the selective detection of 5-methylcytosine in DNA sequences[J].Journal of the American Chemical Society,2013,135(4):1240-1243.
[18] Longo M C,Berninger M S,Hartley J L.Use of uracil DNA glycosylase to control carry-over contamination in polymerase chain reactions[J].Gene,1990,93(1):125-128.
[19] Huang R,Wang J,Mao W,et al.A novel combined bisulfite UDG assay for selective 5-methylcytosine detection[J].Talanta,2013,117:445-448.
[20] Jin S G,Jiang Y,Qiu R,et al.5-Hydroxymethylcytosine is strongly depleted in human cancers but its levelsdonot correlate with IDH1mutations[J].Cancer Research,2011,71(24):7360-7365.
[21] Lian C G,Xu Y,Ceol C,et al.Loss of 5-hydroxymethylcytosine is an epigenetic hallmark of melanoma[J].Cell,2012,150(6):1135-1146.
[22] Huang Y,Pastor W A,Shen Y,et al.The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing[J].PLoS One,2010,5(1):e8888.
[23] Jin S G,Kadam S,Pfeifer G P.Examination of the specificity of DNA methylation profiling techniques towards 5-methylcytosine and 5-hydroxymethylcytosine[J].Nucleic Acids Research,2010,38(11):e125-e125.
[24] Hayatsu H,Shiragami M.Reaction of bisulfite with the 5-hydroxymethyl group in pyrimidines and in phage DNAs[J].Biochemistry,1979,18(4):632-637.
[25] Yu M,Hon G C,Szulwach K E,et al.Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome[J].Cell,2012,149(6):1368-1380.
[26] Booth M J,Branco M R,Ficz G,et al.Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution[J].Science,2012,336(6083):934-937.
[27] Song C X,Szulwach K E,Fu Y,et al.Selective chemical labeling reveals the genome-wide distribution of 5-hydroxymethylcytosine[J].Nature Biotechnology,2010,29(1):68-72.
[28] Song C X,Clark T A,Lu X Y,et al.Sensitive and specific single-molecule sequencing of 5-hydroxymethylcytosine[J].Nature methods,2011,9(1):75-77.
[29] Pastor W A,Pape U J,Huang Y,et al.Genome-wide mapping of 5-hydroxymethylcytosine in embryonic stem cells[J].Nature,2011,473(7347):394-397.
[30] Okamoto A,Sugizaki K,Nakamura A,et al.5-Hydroxymethylcytosine-selective oxidation with peroxotungstate[J].Chemical Communications,2011,47(40):11231-11233.
[31] Zhou X,Yan S,Xu X,et al.A turn-on fluorescence strategy by selective one-step reaction with 5-hydroxymethyl-2′-deoxycytidine for the formation of 1,3-O,N-heterocycles[J].RSC Advances,2013,3:12066-12068.
[32] Hong T T,Wang T,Guo P,et al.Fluorescent strategy based on CCP-FRET for the quantification of 5-hydroxymethylcytosine in genomic DNA[J].Analytical Chemistry,2013,85:10797-10802.
[33] Cadet J,Berger M.Radiation-induced decomposition of the purine bases within DNA and related model compounds[J].International Journal of Radiation Oncology,1985,47(2):127-143.
[34] Djuric Z,Heilbrun L K,Simon M S,et al.Levels of 5-hydroxymethyl-2′-deoxyuridine in DNA from blood as a marker of breast cancer[J].Cancer,1996,77(4):691-696.
[35] Bhutani N,Burns D M,Blau H M.DNA demethylation dynamics[J].Cell,2011,146(6):866-872.
[36] Hong I S,Ding H,Greenberg M M.Oxygen independent DNA interstrand cross-link formation by a nucleotide radical[J].Journal of the American Chemical Society,2006,128(2):485-491.
[37] Xu X,Yan S,Hu J,et al.One-step to get 5-azidomethyl-2′-deoxyuridine from 5-hydroxymethyl-2′-deoxyuridine and detection of it through click reaction[J].Tetrahedron,2013,69(46):9870-9874.
[38] Okamoto A,Tainaka K,Saito I.A dielectric-sensitive fluorescent DNA probe for monitoring polarities on the interior of a DNA-binding protein[J].Bioconjugate Chemistry,2005,16(5):1105-1111.
[39] Cohen B E,McAnaney T B,Park E S,et al.Probing protein electrostatics with a synthetic fluorescent amino acid[J].Science,2002,296(5573):1700-1703.
[40] Okamoto A.Chemical approach toward efficient DNA methylation analysis[J].Organic & Biomolecular Chemistry,2009,7(1):21-26.
[41] Münzel M,Lischke U,Stathis D,et al.Improved synthesis and mutagenicity of oligonucleotides containing 5-hydroxymethylcytosine,5-formylcytosine and 5-carboxylcytosine[J].Chemistry-A European Journal,2011,17(49):13782-13788.
[42] Raiber E A,Beraldi D,Ficz G,et al.Genome-wide distribution of 5-formylcytosine in embryonic stem cells is associ-ated with transcription and depends on thymine DNA glycosylase[J].Genome Biology,2012,13(8):R69.
[43] Hu J,Xing X,Xu X,et al.Selective chemical labelling of 5-formylcytosine in DNA by fluorescent dyes[J].Chemistry-A European Journal,2013,19(19):5836-5840.
[44] Guo P,Yan S,Hu J,et al.Selective detection of 5-formyl-2′-deoxycytidine in DNA using a fluorogenic hydroxylamine reagent[J].Organic Letters,2013,15(13),3266-3269.
[45] Mao W,Hu J,Hong T,et al.A convenient method for selective detection of 5-hydroxymethylcytosine and 5-formylcytosine sites in DNA sequences[J].Organic & Biomolecular Chemistry,2013,11:3568-3572.
[46] Tian T,Zhang X,Fu B,et al.Systematic investigation of DNAs with modified cytosines under hot alkali treatment[J].Chemical Communications,2013,49(85):9968-9970.
[47] Xing X W,Liu Y L,Vargas M,et al.Mutagenic and cytotoxic properties of oxidation products of 5-methylcytosine revealed by next-generation sequencing[J].PloS One,2013,8(9):e72993.
