入侵藤本薇甘菊的耐盐能力
2014-02-07
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 中山大学有害生物控制与资源利用国家重点实验室,广东 广州 510275
入侵藤本薇甘菊的耐盐能力
胡 亮1,2,李鸣光2*,韦萍萍2
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 中山大学有害生物控制与资源利用国家重点实验室,广东 广州 510275
薇甘菊Mikania micrantha Kunth为原产中南美洲的入侵藤本植物,现已在全球热带和亚热带地区照成严重的生态问题。野外观察发现薇甘菊已在珠三角沿海植被中滋生。为揭示薇甘菊的耐盐能力及其对滨海盐生植被的潜在危害性,分别对薇甘菊种子、根系和茎干在不同盐度胁迫下的响应进行了测定。在珠三角地区采集了薇甘菊3个批次的种子,并选取珠江三角洲6种常见植物(菊科假臭草Eupatorium catarium、藿香蓟Ageratum conyzoides、野茼蒿Crassocephalum crepidioides、鬼针草Bidens pilosa、翅果菊Pterocypsela indica和旋花科小花假番薯Ipomoea triloba)作为对照,测定了它们在不同NaCl盐度(质量分数)胁迫下的萌发率和幼苗生长;为揭示薇甘菊种子是否可由海水传播,将薇甘菊和对照植物假臭草、藿香蓟和鬼针草在不同盐度条件下浸泡不同时间后解除胁迫再观测其种子萌发率;为确定薇甘菊能否在滨海盐土中无性繁殖,分别对薇甘菊的根系和茎干在盐水浸泡下的植株生长进行了测定。研究结果表明:1)总体而言,薇甘菊3个种群在≤0.6%盐度胁迫下的种子萌发率均与对照组无显著差异,个别种群的这一阈值可高达1.5%,明显高于其他对照植物;同一盐度胁迫条件下,薇甘菊幼苗根系和苗高生长受抑制程度明显低于对照植物。2)即使经高达3.0%盐度胁迫浸泡长达12 d后,薇甘菊、藿香蓟和假臭草种子的萌发率也没有受到显著影响,而鬼针草种子的萌发率有明显下降;四种植物受浸泡后萌芽的幼苗生长均正常。3)薇甘菊根系受0.5%盐度胁迫时没有植株死亡,但苗高增长仅为对照组的53.3%;根系在1.0%盐度胁迫下有95%的植株能存活至少25 d,但苗高增长仅为对照的18.9%;根系在≥1.5%盐度胁迫下薇甘菊无法存活。4)被清水浸泡的薇甘菊茎茎叶均能存活且在全部被浸泡的节上萌芽。当薇甘菊茎干受≥1.0%盐水浸泡时,受浸泡的叶片全部死亡,在1.0%、2.0%和3.0%盐度胁迫下受试茎第21天的死亡率分别为42.9%、40.9%和86.4%。以上结果表明薇甘菊对盐生生境具一定的适应能力,可能通过种子萌发或无性繁殖扩张的方式对滨海盐生植被构成威胁,应当引起重视。
薇甘菊;入侵植物;NaCl胁迫;种子萌发;耐盐性
生物入侵是除生境破坏之外造成生物多样性丧失的第二主导因素(Wilcove等, 1998)。生物入侵在进化史上无处不在,但自然发生的生物入侵在次数和规模上都有限;随着国际贸易和发展和人类活动的频繁,生态系统被大幅度改变和简化,生物传播、入侵和扩散的种类和数量在全球范围内呈增加的趋势(Elton, 1958; 徐汝梅和叶万辉, 2004)。许多入侵生物对被入侵地的生物多样性、生态系统稳定性和经济等造成了巨大的威胁和长期的负面影响(徐汝梅和叶万辉, 2004; 全国明等, 2001)。有效的管理战略应该包括准确预测到已建立的入侵杂草的可能范围蔓延(Rejmánek, 2000; Ficetola等, 2007)。区域尺度上,入侵生物对非生物环境因子的适应性将为其分布预测提供重要的信息(Zalba等, 2000; 胡亮, 2009)。
薇甘菊(Mikania micrantha Kunth)是受人类活动显著影响而造成严重危害的入侵物种之一,现已在热带、亚热带亚太地区广泛分布(Holm等, 1977; Zhang等, 2004; 张海娟等, 2011)。目前我国广东、广西、云南、海南、江西、台湾等省份均有薇甘菊的分布(王伯荪等, 2003; 黄东光等, 2007;杜凡等, 2006; 范志伟等, 2010; 曾宪锋, 2013)。薇甘菊在新的分布地区能迅速占领被干扰生境(Waterhouse, 1994)。薇甘菊的生长速度迅速,茎节可随处生根,有性生殖能力强,具有缠绕习性等特点使其能迅速蔓延和覆盖大面积的林木,抑制本地植物的生长并降低群落多样性(Waterhouse, 1994; 周先叶等, 2003; 周先叶等, 2005)。
目前国内外相关研究主要涉及薇甘菊的生物学特征、生态学、分布与危害、防治方法等方面。其中对影响薇甘菊的非生物环境研究主要集中在光照(胡玉佳和毕培曦, 1994; 李鸣光等, 2002; Deng等, 2004)、温度(李鸣光等, 2002; Yang等, 2005)和水分(杨期和等, 2003; 杨逢建等, 2005)3个方面。盐度是影响植物生长与分布的重要生态因子之一(Crain等, 2004; 任丽娟等, 2010),但目前未见有关薇甘菊对盐生生境适应性的报道。野外调查发现,广东深圳、珠海和香港特别行政区的滨海盐生植被区均已有薇甘菊的分布,对薇甘菊耐盐能力的对比研究,可揭示薇甘菊的环境适应能力和逆境生存能力,有助于对薇甘菊的危害进行风险评估。
本文将通过一系列实验揭示薇甘菊对华南滨海植被区的潜在入侵能力和危害。为揭示薇甘菊对盐生生境的适应性,本研究将对比薇甘菊和其他6种常见植物在不同程度的盐胁迫下的种子萌发和幼苗生长情况。为揭示是否高渗透势引起的水分胁迫抑制了种子的萌发,本研究将薇甘菊和对照植物在不同盐度条件下浸泡不同时间后解除胁迫再观测其种子萌发率。为进一步研究确定薇甘菊对滨海盐水植被的潜在危害及危害模式,本文分别对薇甘菊的根系和茎干在盐水浸泡下的植株生长情况进行了测定。
1 材料和方法
薇甘菊种子包括2007年4月采自深圳内伶仃(MMN)、2008年11月采自深圳百果园(MMB)和2008年11月采自珠海淇澳岛(MMQ)的3个批次。选取珠江三角洲6种常见植物为对照:菊科草本植物假臭草Eupatorium catarium(EC)、藿香蓟Ageratum conyzoides(AC)、野茼蒿Crassocephalum crepidioides(CC)、鬼针草Bidens pilosa(BP)、翅果菊Pterocypsela indica(PI)及旋花科草质藤本小花假番薯Ipomoea triloba(IT)。所有对照植物的种子采集自2007年10月至2008年11月。薇甘菊3个批次的种子采集前后相差不足2年,而根据我们多年的实验经验表明,在密封干燥条件下保存的薇甘菊种子能在5年后仍保持较高的萌发率。因薇甘菊和对照植物的结实和采集时间不同,所有种子萌发实验均在采集当年或下一年的生长期内完成。用蒸馏水配置不同盐度(质量分数)的NaCl溶液模拟盐度胁迫。
1.1 盐胁迫对植物种子萌发与幼苗生长的影响
于2008年和2009年生长期,选取饱满、无破损的受测种子各1批。在放有2层定性滤纸的9 cm培养皿中分别加入5.0 g盐度分别为0%、0.3%、0.6%、0.9%、1.2%、1.5%、1.8%、2.1%的NaCl溶液(因BP和IT生长迅速,用12 cm培养皿并加入10.0 g溶液),每皿放置30粒种子,每个梯度3个重复。培养皿置于RXZ-300B型人工气候箱中(白天12 h,30 ℃,光照2500~3500 lx;黑夜12 h,28 ℃,无光照)。以胚根突破种皮作为萌发标准,每24 h记录1次萌发数并用称重法补充损失的水分。实验第21天(IT种子的萌发和幼苗生长极为迅速,于第7天结束实验)测量叶片已突破种皮的全部幼苗之苗高和根长。以相对萌发率、相对苗高和相对根长作为种间比较指标。
1.2 盐水浸泡对种子活力的影响
于2008年5月选取将受试种子MMN、AC、EC和BP,在室温、无光照条件下分别在0%、2.0%和3.0%的NaCl溶液中浸泡3、6和12 d后,将种子置于纱布上用蒸馏水反复冲洗。将冲洗过的种子放置于培养皿中定性滤纸上,加入5 mL蒸馏水,每皿30粒受试种子,每个梯度3个重复。将培养皿放置于与2.1相同设置的RXZ-300B型人工气候箱中。以胚根突破种皮作为萌发标准,每24 h记录1次萌发数并补充损失水分。以第7天的萌发率作为种间比较指标。
1.3 薇甘菊根部的耐盐性
在底部具透水孔的小方盆中铺满沙粒,于2008年6月选取MMN种子1批播于沙基上,将小方盆放置在稍高的容器中。在容器中加入0.2倍Hogland宏量元素配方营养液使营养液表面与沙基表面持平,以保证薇甘菊根部全部浸没在稳定的溶液环境中,并避免因毛细作用使盐分在沙基表面聚集。每3 d更新1次营养液,至第21天,留取长势均匀的幼苗,并选取其中10株测量其苗高、叶数、根长、根数。将容器中的营养液换成以0.2倍Hogland宏量元素营养液配置的0%、0.5%、1.5%和2.0%的NaCl溶液,每个梯度幼苗40株。每48 h更新容器中溶液,每5 d记录1次幼苗死亡数。当2.0%盐度下植株全部死亡时结束实验,测量所有存活幼苗的苗高、叶数、根长、根数。计算盐胁迫期间各指标的增长速度。
1.4 薇甘菊茎干的耐盐性
于2008年9月将薇甘菊水培苗移栽于以山地红壤(采自广州龙眼洞)和营养土按1∶1混合的花盆中。待测植株藤茎长度>1.5 m后,选取长势一致、无病虫害、无机械损伤的植株,随机分成4组,每组21~22株。如图1所示,将薇甘菊茎干中端2~3个节浸泡在0%、1.0%、2.0%和3.0%的NaCl溶液中,模拟受海水淹没的匍匐茎;摘除前端顶芽和极少数已长出叶片的叶芽,保留8片成熟叶片以使受测藤茎前端有较一致的叶面蒸腾(图1)。每天补充盐水容器中液体至指定刻度以保持盐度恒定。每天记录各株薇甘菊前端成熟叶片的存活情况,每3 d记录1次前端新生叶的增长和叶芽的活动。实验持续21 d。以成熟叶片和茎的死亡率、整体平均新生叶数和存活茎平均新生叶数等作为比较指标。
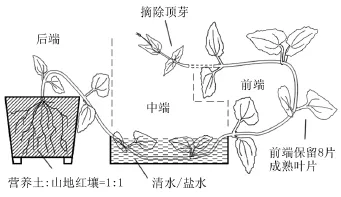
图1 薇甘菊茎干耐盐性实验设置示意图Fig. 1 Schematic diagram of the salt-stress test of the stem of Mikania micrantha
2 实验结果
2.1 薇甘菊在盐胁迫下的种子萌发与生长
2.1.1 种子萌发
盐度显著影响了所有受试植物的种子萌发率。藤本植物MM和IT对盐度的耐受性高,EC、PI和BP次之,AC和CC最低。以相对萌发率80%为盐度耐受性的判断标准,9个受试种群对盐度的耐受性由高到低为MMB、MMQ(1.8%~2.1%)>IT(1.5%~1.8%)>MMN(0.9%~1.2%)>PI、EC(0.6%~0.9%)>BP(0.3%~0.6%)>AC、CC(0%~0.3%)(图2a)。
随着盐度的上升,薇甘菊种子开始出现萌发的时间依次滞后,但其滞后时间要短于其他植物(图3)。在各种盐度胁迫下,受试种子的萌发时间都明显滞后,表现为高盐度组种子开始出现萌发的时间以及萌发率达到最大值的时间都晚于低盐度组。种子开始出现萌发的时间滞后于其对照组的盐度分别为:BP、PI、CC为0.3%,AC、IT为0.6%,MMN为0.9%,MMB、EC为1.2%,而MMQ为1.5%。在2.1%盐度下仍有萌发的5批种子中,薇甘菊3个种群和EC均比对照晚6 d才出现萌发;IT虽没有出现滞后,但前4 d的萌发率仅为2.2%,而对照组已高达97.8%(图3i)。
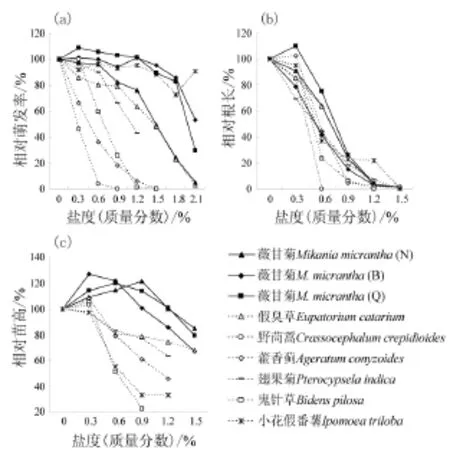
图2 不同盐度胁迫下7种植物第21天的种子相对萌发率与相对生长Fig. 2 Relative germination and growth rates of seven species in the 21st day under different salt stresses
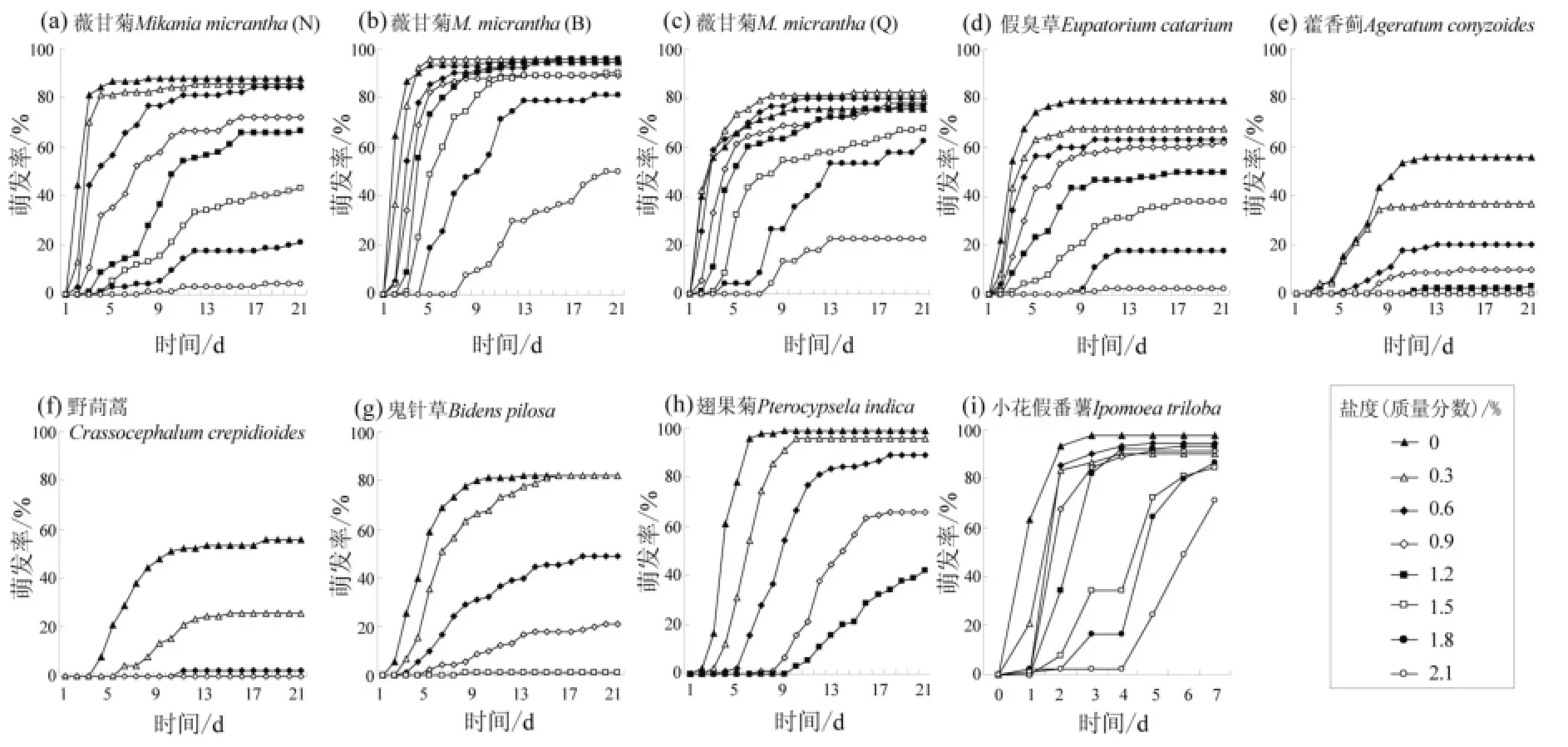
图3 不同盐度下7种植物种子21 d的累计萌发曲线Fig. 3 Germination curves of seven species in 21 days under different salt stresses
总体而言,薇甘菊种子在不高于0.6%盐度下的萌发率与其对照无显著差异,个别种群的这一阈值可高达1.5%。薇甘菊不同种群的种子萌发对盐度的敏感性差异显著,MMB和MMQ2个种群种子敏感性相对较低,而MMN种群对盐度敏感性相对较高(图2a)。MMB和MMQ种群即使在1.5%盐度下的萌发率也都与各自对照无显著差异,而MMN种群仅在0.3%和0.6%盐度下的萌发率与其对照无显著差异(表1)。在0.9%、1.2%、1.5%和1.8%盐度下MMB和MMQ种群间无显著差异,而MMN种群的相对萌发率则显著低于其余2个种群;MMB和MMQ种群的相对萌发率在1.8%盐度条件下都超过80%,而MMN种群的相对萌发率仅为24%。
2.1.2 幼苗生长
总体而言,薇甘菊根系受盐胁迫的抑制程度低于其他植物(表2,图2b)。盐度显著影响了所有受试植物的根系生长。除CC外其余植物在0.3%盐度下的相对根长都高于75%;在0.6%盐度下仅MMN,MMQ和EC的相对根长高于50%;而在0.9%盐度下仅MMQ的相对根长高于25%;1.2%盐度下除IT外,其余种群的相对根长都低于10%。以相对根长50%为盐度敏感性的判断标准,9个受试种群对盐度的耐受性由高到低为:MMN、MMQ、EC(0.6%~0.9%)>MMB、IT、PI、BP、AC、CC(0.3%~0.6%)(图2b)。
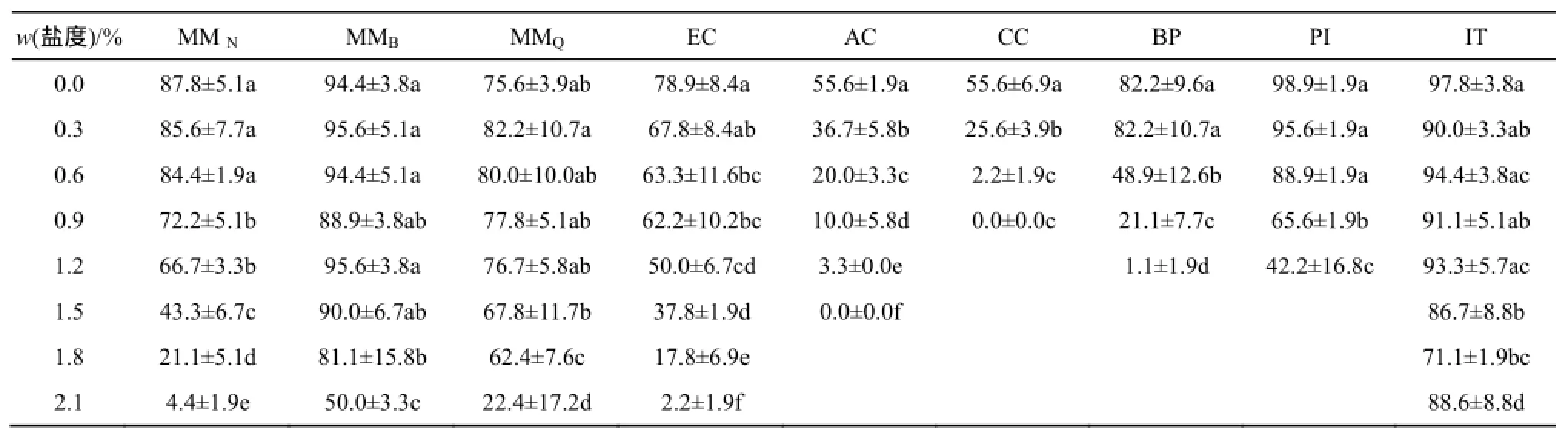
表1 不同盐度(质量分数)胁迫下7种植物21 d的种子萌发率(%)1)Table1 Germination rates of seven species in 21 days under salinity stresses
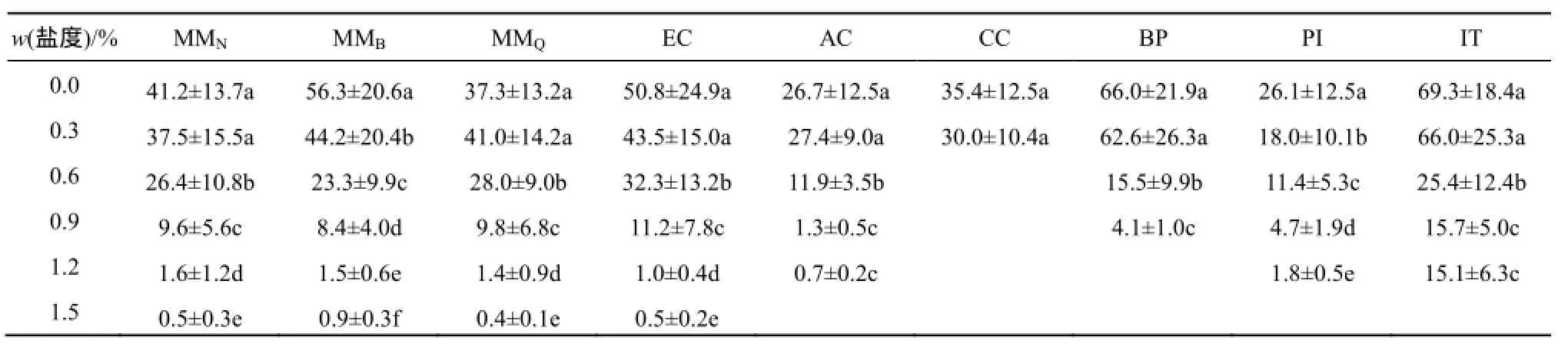
表2 不同盐度(质量分数)胁迫下7种植物生长21 d的平均根长(单位:mm)1)Table2 Average root length (mm) of seven species grew in different salinity stresses
3个受试薇甘菊种群中,MMQ种群的受抑制程度相对较低,MMN种群次之,而MMB种群最高。盐胁迫显著抑制了薇甘菊幼苗根系的生长,随盐度上升,其受抑制程度越高(表2,图2b)。各盐度梯度下3个种群根长受抑制程度的平均值分别为:0.3%盐度下抑制6.9%,0.6%盐度下抑制39.9%,0.9%盐度下抑制78.5%,1.2%盐度下抑制96.6%。
薇甘菊在不高于0.9%的盐度胁迫下相对苗高均高于对照,部分种群在1.2%盐度下也与对照无显著差异,在1.5%盐度胁迫下苗高显著降低,但其受抑制程度显著低于其他植物(表3,图2c)。在0.3%盐度下,所有受试植物的苗高都高于对照或与对照无显著差异。除薇甘菊外,其余受试植物在0.6%盐度下的苗高均显著受到抑制,EC、AC和PI苗高降低约17%~20%;BP和IT降低约45%~49%;而该盐度下CC已萌发幼苗在实验期间全部死亡。
2.2 盐水浸泡后的种子活力
经不同盐胁迫浸泡3、6和12 d后,MMN、AC和EC种子萌发率均没有受到显著影响,2.0%和3.0%盐水浸泡后的种子均与各自对照组无显著差异或不低于对照组(图4a, b, d)。BP种子经2.0%盐水浸泡3~12 d后种子萌发率与对照组没有显著差异,但在3.0%盐水中随浸泡时间增加,萌发率呈下降趋势,浸泡12 d后的种子萌发率显著低于对照组(图4c)。所有4种受试植物的种子在蒸馏水、2.0%和3.0%盐水中浸泡3~12 d后萌发幼苗的胚根、胚芽生长均正常,没有出现死亡。
2.3 薇甘菊根部在盐胁迫下的生长
薇甘菊21 d实生苗根系在0.5%NaCl胁迫下没有植株死亡(图5a)。该盐度下,薇甘菊苗高显著受到抑制,仅为对照处理的53.3%;而叶数、根系长、根数、地上部分干质量等指标的增长受抑制程度相对较低(图5b)。另外,该盐度处理下薇甘菊小根显著变粗。
根系受1.0%的NaCl胁迫时对薇甘菊的各种生长指标均已造成显著抑制,部分植株死亡(图5a)。该盐度下,薇甘菊苗高受抑制程度最高,相对增长仅为18.9%;根系相对增长为51.0%;新生叶数相对增长为53.3%;根数相对增长为40.1%(图5b)。实验第25天,该盐度处理下有5%的植株死亡。
根系在1.5%和2.0%盐度胁迫下的薇甘菊幼苗无法存活(图5a, b)。2.0%盐度下第3天即出现叶片和茎干变黄、植株倒伏,第5天就开始出现植株死亡;1.5%盐度比2.0%盐度晚5天才出现植株死亡。实验第25天,2.0%幼苗全部死亡时,此时1.5%盐度下幼苗死亡率达到95%;而余下的2株薇甘菊生长不良,植株倒伏,苗高和根系相对增长分别为8%和14.1%。
2.4 茎干在盐胁迫下的薇甘菊生长
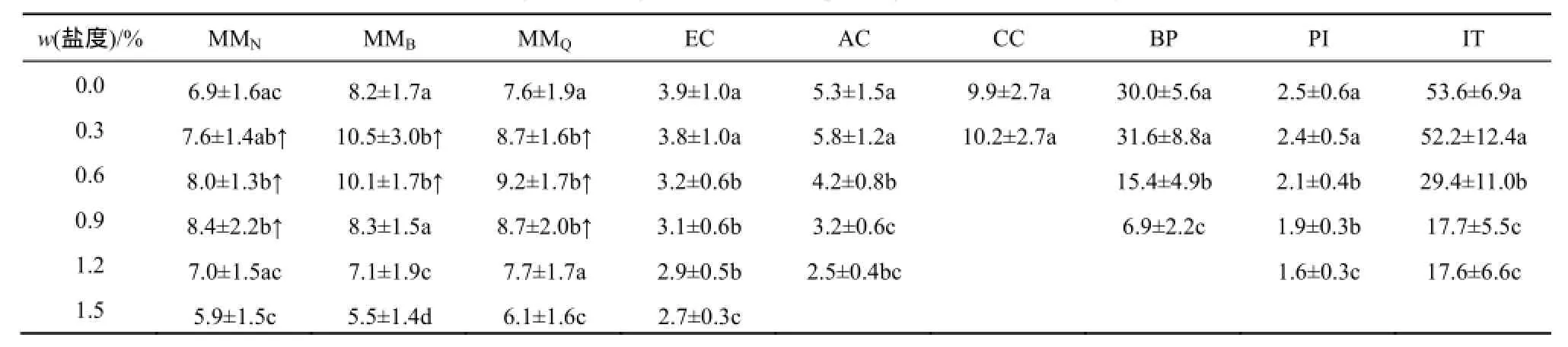
表3 不同盐度(质量分数)胁迫下7种植物生长21 d的平均苗高(单位:mm)1)Table3 Average shoot length (mm) of seven species grew in different salinity stresses
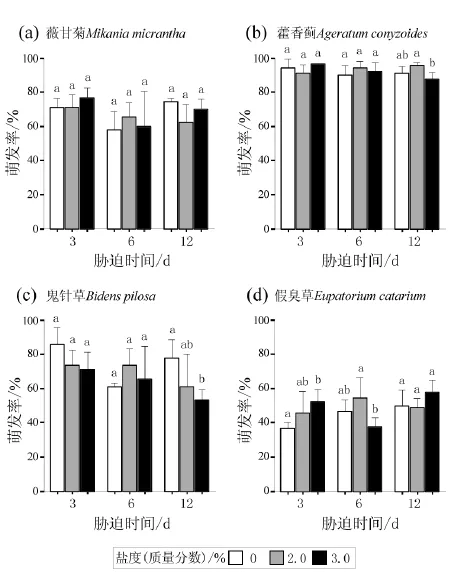
图4 四种植物种子在不同盐度条件下浸泡3、6和12 d后的萌发率Fig. 4 Germination rates of four species dealt with 3, 6 and 12 days immersion in different salt stresses
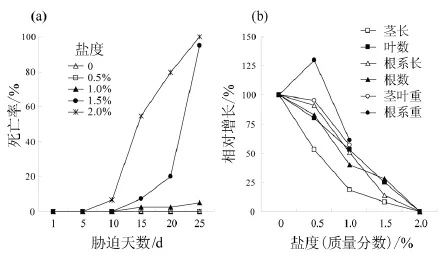
图5 薇甘菊根部在盐胁迫条件下的幼苗死亡率与相对增长速率Fig. 5 Mortalities and relative growth of Mikania micrantha grew in different salt stresses
1)成熟叶片与茎干的死亡。所有受试薇甘菊植株成熟叶片的凋亡只出现在被浸泡中端和前端,植株后端均没有出现死亡。被浸泡的中端叶片除清水对照组外,其余梯度的叶片均全部死亡,且盐度越高,被浸叶片死亡速度越快。实验第3天开始,各梯度薇甘菊的植株前端均开始出现叶片死亡,其中3.0%盐度下的死亡最快,而1.0%和2.0% 2种盐度下薇甘菊叶片死亡情况没有显著差异。至第21天,清水、1.0%、2.0%和3.0%盐度中薇甘菊累计死亡率分别为0.6%、48.7%、44.5%和91.6%(图6a)。
受试茎的死亡也仅出现在被浸泡中端和前端,被浸泡后端均能生长正常。在3.0%梯度下第4天即开始出现枝条死亡,而在1.0%和2.0% 2种盐度下均是在第10天才开始出现茎干死亡。受试茎的死亡率情况与叶片死亡率情况基本一致,第21天清水、1.0%、2.0%和3.0%盐度中薇甘菊受试茎前端的死亡率分别为0、42.9%、40.9%和86.4%(图6a)。
2)新叶与根的生长。受不同盐度胁迫的薇甘菊茎前端叶腋的芽均有一定的活力,能生长出新的枝条和叶片;随盐度增加,有新叶长出的芽所占的比例逐渐降低。各梯度有新叶生长活动芽的比例分别为对照组62.9%,1.0%盐度53.2%,2.0%盐度57.8%,3.0%盐度36.7%。
盐胁迫初期(0~6 d),盐度越高,新叶增长速度越快;随胁迫时间的延长,各盐度胁迫下的新叶增长速度逐渐降低至对照水平以下(图6b, c)。各梯度的整体平均新叶增长速率分别为对照组1.20叶/d、1.0%盐度0.75叶/d、2.0%盐度0.83叶/d、3.0%盐度0.15叶/d。对照组在整个实验过程中的新叶增长速率保持相对稳定(图6b, c)。3.0%盐度的整体增长速度降低最快,而1.0%和2.0%梯度之间的整体增长变化没有显著差异。
由于没有茎干死亡,对照组的存活茎平均值与整体平均值完全一致;各盐度梯度下的存活茎平均值都低于其整体水平,表明死亡植株上的新叶增长在死亡前是高于整体平均值的。
对照组被清水浸泡部分的茎叶全部存活,且能在被浸泡的节上萌生新的枝叶;其余各组被浸泡部分的叶片均全部死亡。对照组被浸节上100%能生根;而在1.0%盐度下的21株薇甘菊仅有1株在被浸节上的全部3个节生根,其余植株没有根的生成;其余高盐度胁迫组的被浸节部均没有根的生成。
3 讨论与分析
薇甘菊种子能耐受低含量的盐胁迫,可在滨海盐生生境下萌发生长。在0.9%盐度条件下的薇甘菊种子相对萌发率在第7天时均高于90%,比沙漠地肤(Kochia americana)、碱蓬(Suaeda salsa)、碱茅(Puccinellia tenuiflora)、野麦草(Hordeum jubatum)等耐盐植物在相同盐度下的相对萌发率还要高(Clarke和West, 1969; 李昀等, 1997; 赵可夫和范海, 2005)。非盐生植物对盐度的低敏感性还在黄瓜(Cucumis sativus)、西葫芦(Cucurbita pepo)、甜瓜(Cucumis melo)、西瓜(Citrullus lanatus)等藤本植物中被发现(陈国雄等, 1996; 王广印等, 2004; 王春林等, 2006; 王吉明和马双武, 2007; 张秀玲, 2008; 高美玲等, 2008),推测藤本植物对盐度的敏感性可能低于自支持植物。在盐胁迫下薇甘菊种子萌发和幼苗生长受抑制程度显著低于其它植物,表明薇甘菊吸取水分的能力强于其它受试植物。在淇澳岛滨海红树林的观察发现,薇甘菊能在一定的盐度环境下完成生活史,受周期性海水淹没的薇甘菊种群(MMQ)所产生的种子萌发率可达到80%以上。
短时间的淡水和海水浸泡对薇甘菊、藿香蓟、假臭草和鬼针草种子的萌发率都没有显著影响,因此这些种子不但能沿淡水水系传播,还能通过海流沿海岸传播。高盐度导致的水分胁迫虽然抑制种子萌发但不致死,解除胁迫后种子能正常萌发和生长。
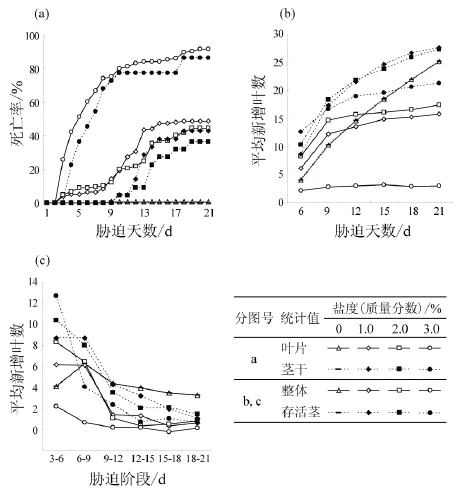
图6 薇甘菊茎干在不同盐胁迫条件下的茎干、叶片死亡率和新叶增长速率Fig. 6 Mortality of shoots and leaves, and growth of new leaves of Mikania micrantha grew in different salt stresses
薇甘菊的根系和茎干均具一定的耐盐能力,且根系耐盐性高于茎干;因此薇甘菊可能由非盐生环境下生长的种群通过枝条蔓延扩张到临近的盐生生境中。盐胁迫下薇甘菊幼苗根系生长缓慢,苗高增长加快;而根部受胁实验中薇甘菊苗高受抑制程度显著高于根系。根系和茎干在1.0%盐胁迫下20天的死亡率分别为2.5%和48.7%,亦表明根系对盐环境的耐受性高于茎干。茎干受胁实验中,薇甘菊根部不在盐水浸泡中,没有直接受到水势胁迫;而在被浸泡部分茎干处,茎干可能因外界溶液水势较低而失水,同时盐离子也可能渗入茎干。在潮间带,实际的海水浸泡是周期性的,周期性浸泡可能缓解薇甘菊的受胁迫程度,从而提高其存活率。
薇甘菊的快速增长可能有利于稀释体内盐分的含量。薇甘菊不具有专门的泌盐结构,也没有体现出喜盐或需盐的特征,在盐胁迫不断增加时因胁迫所造成的胁变单调增加。在薇甘菊茎干受盐水胁迫实验的前端成熟叶片表皮气孔周围可观察到细小的盐晶,推测薇甘菊具有一定的泌盐能力;但当根系环境得不到改善,高盐度胁迫下盐分的输入超出叶片的泌盐能力,盐分在薇甘菊植株内的积累速度高于其排盐能力,最终将导致植株死亡。盐胁迫初期薇甘菊地面部分的快速生长在萌发实验和茎干受胁实验中均有出现。当生物量增长的效应使盐分含量低于阈值时,植株仍可存活;当体内盐分含量超过阈值后,部分叶片因积累盐分较多而死亡。观测结果表明,随着部分叶片的死亡,枝条稀释盐分的总能力也减弱,新生叶芽受到的盐胁迫程度相对增加,因而死亡率也会升高。被浸茎干前端叶片先于被浸茎干的死亡表明存在离子毒害;后端叶片和茎干的正常生长,可能是由于被浸茎干渗入的盐分未输送到根部,或至少输送到根部的量较少,没有达到毒害水平,因此后端成熟叶片和新生叶不共同分担和降低植株前端的盐度。以此推测,当薇甘菊匍匐枝条的某节在地面产生新根后,所吸取的养分和水分也几乎全部用于芽头方向枝叶的生长。一方面,这可能有利于盐生生境中植株整体的安全性,使在盐度较低的地段蔓延部分不受牵连;另一方面,薇甘菊可借助其在非盐土区汲取的养分和水分支持其在滨海植被上的蔓延。另外,在茎干受胁实验中所有盐度梯度中观测到薇甘菊叶片的肉质化,且在新生叶中表现较明显;叶片肉质化也一定程度上会对缓解和稀释体内盐分有所帮助。
据本文的研究结果,薇甘菊可在华南大部分滨海地区生长。广东珠三角地区滨海土壤盐度为0.08%~0.87%,广东东部沿海土壤盐度0.22%~1.69%,西部沿海0.91%~1.44%(刘腾辉和杨萍如, 1990);海南沿海红树林中土壤盐度为0.41% ~1.59%(刘美龄等, 2008);广西为0.23% ~ 2.51%(梁士楚等, 2008; 徐淑庆等, 2010)。薇甘菊可能通过种子扩散或无性繁殖扩张到这些生境。目前在广东滨海盐土地区已发现有薇甘菊的存在,如果不及时治理,有可能在滨海地区进化出耐盐能力更强的新生态型。
研究发现,用11%、14%和17%的盐溶液喷施薇甘菊,在喷施后7 d,11%盐度未能彻底致死薇甘菊,14%盐度能致死全部叶片,但根和茎仍有10%存活,而17%盐度能致死全部的根、茎和叶(Hu和Wu, 2003)。但这种超高盐度远超过薇甘菊和一般植物的耐受能力,而且盐溶液浇灌不仅直接作用于植物植株,还会残留于土壤中,造成次生盐渍化问题,实际情况中无法采用。低盐度的溶液多次喷施可能致死速度会减缓,但有利于土壤中残留盐分的稀释。滨海盐土用盐溶液治理薇甘菊造成的盐分残留和积累可稀释至海水中;但山地和耕地区的稀释不易。耕地本身多属地下水位高,排水不良的集盐区,若再施以盐溶液,容易造成次生盐渍化;山地的氯化盐分集中与山脚地势低洼,排水不良的区域,山地盐浇灌也会导致这些地区土壤的盐度增加,造成次生环境问题,因此,即使是低盐度浇灌或喷施,也不宜在这些地区采用。
4 结论
薇甘菊对盐生生境具一定的适应能力。一方面,其种子在盐生生境中的萌发和生势明显高于其他对照植物;即使受长达12 d的高盐度海水浸泡,仍能保持种子的活性和萌发幼苗的正常生长,因此薇甘菊完全可能通过海流在滨海生境中以种子形式扩散;另一方面,薇甘菊的根系和茎干也具有一定的耐盐能力,可能通过匍匐茎在盐生生境中伸长和扎根的方式扩张。薇甘菊对滨海植被的破坏,可能引发次生的生态问题,应当引起重视。
CLARKE L D, WEST N E. 1969. Germination of Kochia americana in relation to salinity[J]. Journal of Range Management, 22(4): 286-287.
CRAIN C M, SILLIMAN B R, BERTNESS S L et al. 2004. Physical and biotic drivers of plant distribution across estuarine salinity gradients[J]. Ecology, 85: 2539-2549.
DENG Xiong, YE Wanhui, FENG Huiling et al. 2004. Gas exchange characteristics of the invasive species Mikania micrantha and its indigenous congener M. cordata (Asteraceae) in South China[J]. Botanical Bulletin of Academia Sinica, 45: 213-220.
ELTON C S. 1958. The ecology of invasions by animals and plants[M]. London: Methuen.
FICETOLA G F, THUILLER W, MIAUD C. 2007. Prediction and validation of the potential global distribution of a problematic alien invasive species-the American bullfrog[J]. Diversity and Distributions, 13: 476-485.
HOLM L G, PLUCKNETT D L, PANCHO J V et al. 1977. The world’s worst weeds: distribution and biology[M]. Hawaii: The University Press of Hawaii.
HU Yuja, WU Jianjun. 2003. Allelopathic effects and responses to chemical stresses of Mikania micrantha Kunth[C]//ZHANG Runjie, ZHOU Changqing, PANG Hong et al. Exotic pests and their control. Guangzhou: Sun Yat Sen University Press: 180-186.
REJMÁNEK M. 2000. Invasive plants: approaches and predictions[J]. Austral Ecology, 25: 497-506.
WATERHOUSE D F. 1994. Biological Control of Weeds: Southeast Asian Prospects[M]. Canberra: Australian Centre for International Agricultural Research.
WILCOVE D S, ROTHSTEIN D, DUBOW J, et al. 1998. Quantifying threats to imperiled species in the United States[J]. BioScience, 48(8): 607-615.
YANG Qihe, YE Wanhui, DENG Xiong et al. 2005. Seed germination eco-physiology of Mikania micrantha H.B.K.[J]. Botanical Bulletin of Academia Sinica, 46: 293-299.
ZALBA S M, SONAGLIONI M I, COMPAGNONI C A et al. 2000. Using a habitat model to assess the risk of invasion by an exotic plant[J]. Biological Conservation, 93: 203-208.
ZHANG L Y, YE W H, CAO H L et al. 2004. Mikania micrantha H.B.K. in China-an overview[J]. Weed Research, 44: 42-49.
曾宪锋. 2013. 薇甘菊在赣南的首次详实 记录[J]. 广东农业科学, 1: 181-197.
陈国雄, 李定淑, 张志谦, 等. 1996. 盐胁迫对西葫芦和黄瓜种子萌发影响的对比研究[J]. 中国沙漠, 3: 307-310.
杜凡, 杨宇明, 李俊清, 等. 2006. 云南假泽兰属植物及薇甘菊的危害[J].云南植物研究, 28(5): 505-508.
范志伟, 程汉亭, 沈奕德, 等. 2010. 海南薇甘菊调查监测及其风险评估[J]. 热带作物学报, 31(9): 1596-1601.
高美玲, 袁成志, 赵丽娟, 等. 2008. NaCl胁迫对两个甜瓜品种种子萌发的影响[J]. 北方园艺, 8: 15-17.
胡亮. 2009. 薇甘菊Mikania micrantha H.B.K.对非生物环境改变的响应及其分布预测[D]. 广州: 中山大学.
胡玉佳, 毕培曦. 1994. 微甘菊生活史及其对除莠剂的反映研究[J]. 中山大学学报: 自然科学版, 33(4): 88-95.
黄东光, 周先叶, 昝启杰, 等. 2007. 香港郊野公园薇甘菊的化学防除研究[J]. 华南师范大学学报: 自然科学版, 3: 109-131.
李鸣光, 张炜银, 王伯荪, 等. 2002. 薇甘菊种子萌发特性的初步研究[J].中山大学学报: 自然科学版, 41(6): 57-59.
李昀, 沈禹颖, 阎顺国. 1997. NaCl胁迫下5种牧草种子萌发的比较研究[J]. 草业科学, 14(2): 50-53.
梁士楚, 蒋潇潇, 李峰. 2008. 广西英罗港红树植物木榄种群年龄结构的研究[J]. 海洋学研究, 26(4): 35-40.
刘美龄, 叶勇, 曹长青, 等. 2008. 海南东寨港红树林土壤粒径分布的分形特征及其影响因素[J]. 生态学杂志, 27(9): 1557-1561.
刘腾辉, 杨萍如. 1990. 广东的海涂土壤[J]. 土壤学报, 27(4): 427-437.
全国明, 章家恩, 谢俊芳, 等. 2011. 薇甘菊入侵对中小型土壤动物群落结构特征的影响[J]. 应用生态学报, 22(7): 1863-1870.
任丽娟, 仇乐, 王国祥, 等. 2010. 盐胁迫对淡水浇灌互花米草种子的萌发及胚生长的效应[J]. 植物研究, 30(3): 305-310.
王伯荪, 廖文波, 昝启杰, 等. 2003. 薇甘菊Mikania micrantha 在中国的传播[J]. 中山大学学报: 自然科学版, 42(4): 47-54.
王春林, 张玉鑫, 陈年来. 2006. NaCl胁迫对甜瓜种子萌发的影响[J]. 中国蔬菜, 5: 7-10.
王广印, 张百俊, 赵一鹏, 等. 2004. NaCl胁迫对黄瓜种子萌发的影响[J].吉林农业大学学报, 26(6): 624-627.
王吉明, 马双武. 2007. NaCl胁迫对西瓜种子发芽的影响[J]. 北方园艺, 3: 20-22.
徐汝梅, 叶万辉. 2004. 生物入侵:理论与实践[M]. 北京: 科学出版社.
徐淑庆, 李家明, 卢世标, 等. 2010. 广西北部湾红树林资源现状及可持续发展对策[J]. 生物学通报, 45(5): 11-14.
杨逢建, 张衷华, 王文杰, 等. 2005. 水分胁迫对入侵植物薇甘菊幼苗生长的影响[J]. 植物学通报, 22(6): 673-679.
杨期和, 冯惠玲, 叶万辉, 等. 2003. 环境因素对薇甘菊开花结实影响初探[J]. 热带亚热带植物学报, 11(2): 123-126.
张海娟, 陈勇, 黄烈健, 等. 2011. 基于生态位模型的薇甘菊在中国适生区的预测[J]. 农业工程学报, (增刊1): 413-418.
张秀玲. 2008. 盐胁迫对绿豆种子萌发的影响[J]. 北方园艺, 4: 52-53.
赵可夫, 范海. 2005. 盐生植物及其对盐渍生境的适应生理[M]. 北京:科学出版社.
周先叶, 王伯荪, 李鸣光, 等. 2005. 薇甘菊对内伶仃岛植被危害的相关分析[J]. 应用生态学报, 16(2): 350-354.
周先叶, 昝启杰, 王勇军, 等. 2003. 薇甘菊在广东的传播及危害状况调查[J]. 生态科学, 22(4): 332-336.
Salt tolerance of the invasive vine Mikania micrantha Kunth
HU Liang1,2, LI Mingguang2*, WEI Pingping2
1. Geography and Planning School, Sun Yat-sen University, Guangzhou 510275, Chian; 2. State Key Laboratory of Biocontrol, Sun Yat-sen University, Guangzhou 510275, China
Mikania micrantha Kunth is a noxious invasive species in tropical and subtropical regions. This weed has been found to grow on the coast of the Pearl River Delta and off-coast islands in China. In the present study, we assessed the salt tolerance of M. micrantha seeds, roots, and stems separately. Seed germination and seedling growth in different concentrations of NaCl were tested for three M. micrantha populations and six common species (Eupatorium catarium, Ageratum conyzoides, Crassocephalum crepidioides, Bidens pilosa, Pterocypsela indica, and Ipomoea triloba) in the Pearl River Delta. Seed germination of M. micrantha, E. catarium, A. conyzoides, and B. pilosa were tested after 3, 6, and 12 days of submergence in different concentrations of NaCl to determine whether seeds can be transferred by seawater. Responses of submerged cultured M. micrantha root and stem in different concentrations of NaCl were also recorded to determine whether the weed can disperse by vegetative propagation in coastal saline soils. Experiments produced the following results. 1) In general, the seed germination rates of M. micrantha did not differ significantly from those of the controls at ≤0.6% salinity, and the highest threshold value of 1.5% was greater than those for all control species. Radicle and seedling height growth rates of M. micrantha were less suppressed than those of control plants under the same salinity stress. 2) In comparison with controls, no significant decline in germination rate was observed in treated (submerged in up to 3.0% NaCl for up to 12 days) M. micrantha, A. conyzoides, or E. catarium, whereas submergence in 3.0% NaCl for 12 days caused a significant decline in B. pilosa. However, seedlings of all four tested species grew well. 3) All tested M. micrantha plants survived 0.5% salinity stress to root systems, with a decline in the average seedling height growth rate to 53.3% those of controls; 95% of tested plants survived up to 1.0% salinity stress to root systems for at least 25 days, with a decline in the average seedling height growth rate to 18.9%; no M. micrantha plant survived 1.5% salinity stress to the root system. 4) All leaves of M. micrantha survived stem submergence in water, with sprouting observed in all submerged nodes, whereas all submerged leaves died when stems were exposed to ≥1.0% salinity stress. Mortality rates of the tested stems at day 21 were 42.9%, 40.9%, and 86.4% for 1.0%, 2.0%, and 3.0% salinity stress, respectively. We conclude that M. micrantha can disperse in coastal saline soils by seed and vegetative propagation. More attention should be paid to the potential threat of M. micrantha populations near coastal halophytic vegetation.
Mikania micrantha Kunth; invasive plant; NaCl stress; seed germination; salt tolerance
Q948
A
1674-5906(2014)01-0007-09
胡亮,李鸣光,韦萍萍. 入侵藤本薇甘菊的耐盐能力[J]. 生态环境学报, 2014, 23(1): 7-15.
HU Liang, LI Mingguang, WEI Pingping. Salt tolerance of the invasive vine Mikania micrantha Kunth [J]. Ecology and Environmental Sciences, 2014, 23(1): 7-15.
国家自然科学基金项目(30370243;41101057)
胡亮(1982年生),男,讲师,博士,研究方向为入侵生物学。E-mail: huliang_hy@163.com;*通信作者:E-mail: lsslmg@mail.sysu.edu.cn
2013-12-19
