光和不同打破种子休眠方法对紫茎泽兰种子萌发及幼苗状态的影响
2013-12-25李艳红王文杰包松莲祖元刚王慧梅
姜 勇,李艳红,王文杰,*,包松莲,祖元刚,王慧梅
(1. 东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040; 2. 中国林科院资源昆虫研究所,昆明 650224)
紫茎泽兰(Eupatoriumadenophorum),属菊科多年生草本植物或亚灌木,与其它植物竞争水分、光照和生存空间等,是一种世界性的恶性杂草[1]。对紫茎泽兰的研究表明,其具有强阳性植物典型特征,但也能够在阴暗环境下生长,光能够影响紫茎泽兰种子萌发与成苗过程,但不同研究之间、室内和野外研究结果之间尚存在较大差异[2-6],很难定量这一入侵植物种子萌发需要的光照强度、其它打破休眠方法是否可以替代光以及其对幼苗初期生长状态如何影响等问题。对包括紫茎泽兰在内的多种入侵植物种子研究发现,有光条件下发芽率显著高于无光对照[6],本研究拟在此基础上,通过类比的方法对紫茎泽兰种子需光萌发的机制进行深入探讨:即在不同光照条件下,看常规打破休眠的方法(低温处理[7]、水杨酸(SA)处理[8]、干旱聚乙二醇(PEG)处理[9-10]、硝酸钾[11]处理)是否代替光照处理,如果能够,则说明光照打破休眠的机理与此类似,否则说明其存在不同的机制,如种子静态等[12-13];同时对光照及不同处理影响幼苗生长(根长、芽长和幼苗鲜重)进行研究。预期光照对种子萌发和幼苗生长影响的研究结果将有助于理解外来植物紫茎泽兰在入侵定植阶段的深层原因。
1 材料和方法
1.1 实验材料
供试紫茎泽兰种子于2010年3月中旬采自于云南省昆明市世博园附近,千粒重48 mg,室温干燥、储藏备用,挑选籽粒饱满,大小均匀的种子用于实验。
1.2 实验设计与发芽处理方法
对紫茎泽兰种子进行不同光照和不同打破种子休眠方法的双因素控制实验,观察二因素对种子的萌发、根长、芽长以及幼苗鲜重的影响及是否存在交互作用。其中光照处理包括黑暗(生长箱光照不打开)、铝箔纸覆盖、全光照3个水平:以全光照为100%,可见光范围内,铝箔纸覆盖的透光率为0.23%,黑暗处理透光率为0%(图1)。
不同打破种子休眠的处理方法包括低温处理、水杨酸处理(SA处理)、聚乙二醇处理(PEG处理)、硝酸钾处理。每个处理的水平数见表1。具体处理方法:低温处理是将种子放入垫有两层滤纸的培养皿中,滤纸用蒸馏水浸湿,之后将装有种子的培养皿放入5℃和-20℃冰箱培养1周,低温打破休眠期间没有光照;水杨酸处理是指种子在0.01、0.05、0.1 mmol/L 水杨酸溶液中浸泡20—21 h;聚乙二醇处理是指种子在5%、10%、15% 聚乙二醇(PEG-6000)溶液中浸泡20—21 h;硝酸钾处理是指种子在10、25、50 mmol/L硝酸钾溶液中浸泡22—23 h(表1)。
以直径为9 cm的培养皿为发芽床,内垫2层滤纸,将上述每个处理的种子分别装入3个培养皿中,均匀地排布,分别以蒸馏水处理种子为对照,每个培养皿内装入40粒种子。发芽培养皿放入生长箱,铝箔纸遮光处理和全光照处理设置为白天16 h光照、温度25℃和晚上8 h黑暗、温度15℃交替处理。所用生长箱为:黑龙江省东扩仪器制造有限公司生产ZPW-400智能植物培养箱,全光照是指打开24个荧光灯管,光照强度11640lx。

表1 实验设计

图1 不同波长下铝箔纸的透光性能
1.3 发芽率、幼苗状态测定与数据处理方法
以子叶展开为正常发芽标准,对种子发芽进行统计[6]。当连续3d无新发芽种子出现,即视为发芽过程结束。芽重、芽长和根长在发芽率测定结束时进行测定,重量测定天平精度为0.001g,发芽率(%)=发芽种子数/供试种子数×100%。
在使用可重复双因素方差分析对光照和各种打破休眠方法对发芽指标的影响进行统计分析的基础上,使用Duncan′s多重比较方法,对同一因素内不同水平的差异显著性进行检验(SPSS 17.0)。
2 结果与分析
2.1光照与低温处理对紫茎泽兰种子的萌发和幼苗生长的影响
在全光照、加铝箔纸和黑暗条件下,-20℃处理使种子丧失活力,不萌发;全光照对室温和5℃处理条件下发芽特征影响类似,均表现黑暗条件下发芽率显著低于光照,相差30%左右(P<0.05);芽长在光照条件下仅2 mm左右,而黑暗和加铝箔纸处理则接近14 mm;全光照处理显著增加根系生长,达到18—20 mm左右,而黑暗和加铝箔纸处理均2 mm左右;幼苗鲜重在全光照条件下明显高于黑暗和加铝箔纸,后二者差异不明显(图2)。
与全光照对各个指标均存显著影响相比(P<0.0005),温度处理对发芽率(P=0.54)、芽长(P=0.44)、根长(P=0.43)和幼苗鲜重(P=0.81)均不存在明显影响。即温度处理并不能影响光照对种子萌发特征的影响(表2)。
2.2 光照与水杨酸处理对紫茎泽兰种子的萌发和幼苗生长的影响
黑暗条件下种子的萌发率显著低于全光照 (P<0.05) ,全光照与加铝箔纸处理差异不显著(图3)。与此不同,芽长在加铝箔和黑暗处理下,2个(对照和0.1 mmol/L)差异达到显著,另外2个不显著,但二者都显著高于全光处理。总体来看,芽长在全光照条件下为2 mm左右,黑暗和加铝箔纸处理为13—15 mm。根长与芽长相反,全光照促进根系生长(12—14 mm左右),而黑暗和加铝箔纸下不同水杨酸处理下均无差异(2 mm左右)(图3)。
双因素方差分析表明(表2),全光照对各个指标均存在显著影响(P<0.0005),而水杨酸处理对发芽率(P=0.59)、芽长(P=0.83)、根长(P=0.79)不存在明显影响,但对幼苗鲜重(P=0.03)影响显著,并与全光照存在交互作用(表2)。全光照下,随着水杨酸浓度提高,幼苗鲜重从0.8 mg增加到1.3 mg,而黑暗下,从0.7 mg增加到1.0 mg;但在铝箔纸下,普遍低于全光照和黑暗,变化范围在0.4 mg—0.7 mg之间(图3)。线性相关分析发现,全光照和加铝箔条件下,幼苗鲜重与水杨酸浓度不存在显著相关性,而在黑暗下,则相关性达到显著水平(表3)。
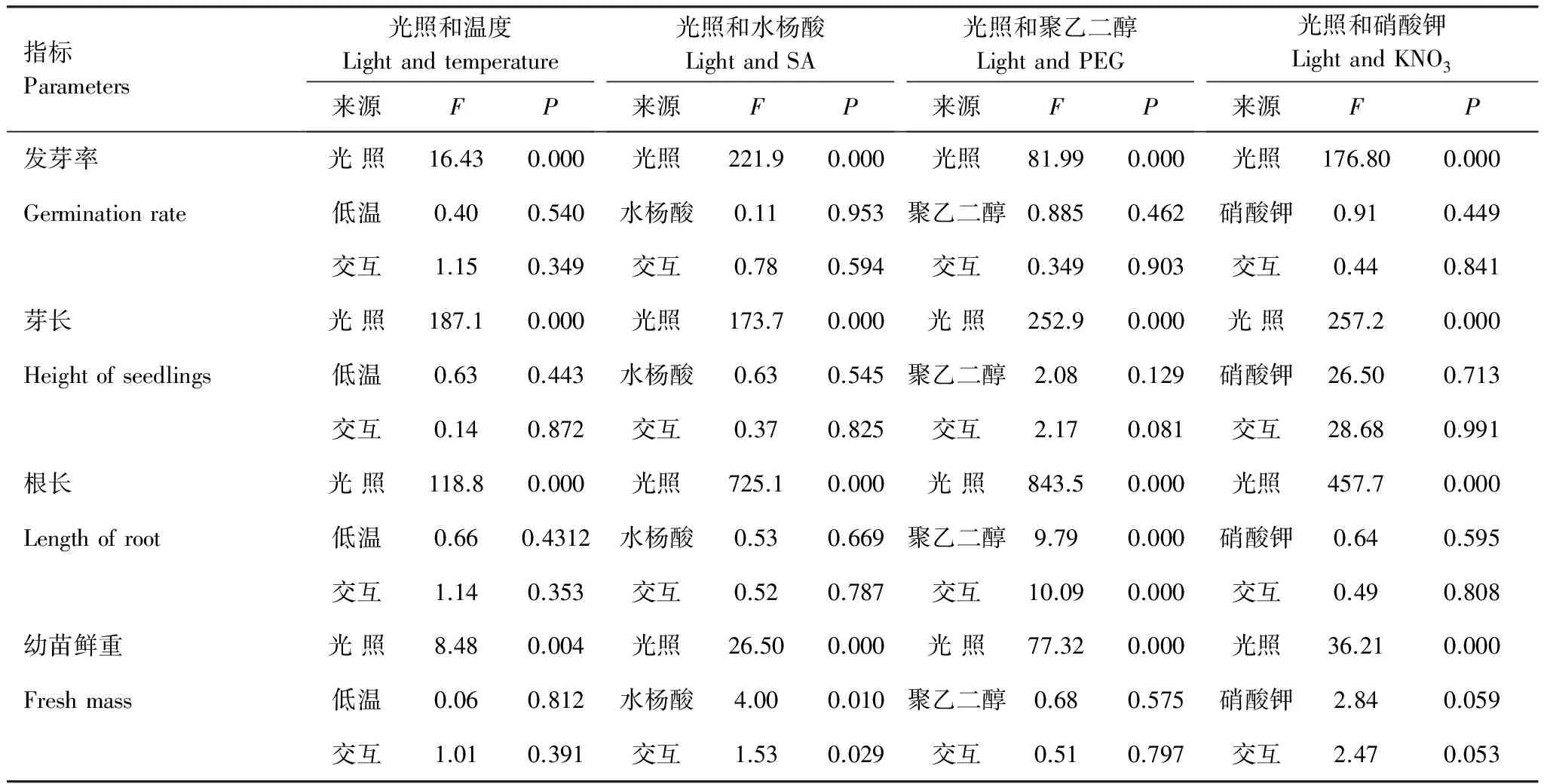
表2 光照和不同打破休眠方法对紫茎泽兰种子发芽特征的双因素方差分析

图2 不同温度和光照处理对紫茎泽兰种子发芽率、芽长、根长、幼苗鲜重的影响
2.3 光照与聚乙二醇处理对紫茎泽兰种子的萌发和幼苗生长的影响
在全光照和加铝箔纸条件下,不同聚乙二醇浓度处理对于发芽率的影响均不显著(P>0.05),但二者均显著高于黑暗处理(P<0.05)(图4),显示光照是影响种子萌发的首要因素,而聚乙二醇处理没有显著的影响(表2)。对芽长的影响,也体现在全光照下,明显抑制芽长的生长,多数情况下,加铝箔的芽长较黑暗下稍长,但多数情况下差异不显著(图4),双因素方差分析显示,聚乙二醇处理没有显著影响芽长生长(表2)。幼苗鲜重在全光照条件下明显高于黑暗和加铝箔纸处理,但后二者差异不显著(图4)。统计分析也显示聚乙二醇浓度对鲜重影响没有达到显著水平(表2)。

表3 水杨酸处理、聚乙二醇处理和硝酸钾处理浓度下紫茎泽兰幼苗鲜重、根长的拟合关系受光照的影响

图3 不同浓度水杨酸处理和光照对紫茎泽兰种子发芽率、芽长、根长、幼苗鲜重的影响
根系生长受光的调节,但微弱的光(加铝箔)与黑暗在不同的聚乙二醇浓度处理下种子的根系生长差异均不显著,说明微弱的光对其没有影响(图4)。双因素方差分析显示,全光照和聚乙二醇处理均能够显著影响根系长度,且存在交互作用(表2),全光照下聚乙二醇处理浓度增加,使根长显著增长,但是在加铝箔和黑暗条件下,这一趋势不明显(图4)。进一步线性回归分析发现,全光照下根系长度随聚乙二醇浓度增大而显著增加,但在其它两个处理下没有显著变化(表3)。

图4 不同浓度聚乙二醇和光照对紫茎泽兰种子发芽率、芽长、根长,幼苗鲜重的影响
2.4 光照与硝酸钾处理对紫茎泽兰种子的萌发和幼苗生长的影响
与前面几个处理相似,硝酸钾不同处理下,黑暗条件下发芽率显著低于全光照和加铝箔处理,相差50%左右(P<0.05),但加铝箔纸与全光照处理差异不显著(图 5)。芽长在全光照条件下,长度为1.8—2.4 mm之间,不同浓度硝酸钾下均表现出显著低于黑暗和加铝箔纸处理(14—15 mm)(图5)。根长在光照条件下达到16 mm左右,而黑暗和加铝箔纸处理均2 mm左右,显著低于全光照(P<0.05)(图5)。幼苗鲜重在全光照条件下明显高于黑暗和加铝箔纸处理,但后二者差异不明显(图5),双因素方差分析显示,硝酸钾处理没有显著影响发芽率、芽长、根长,但是对幼苗鲜重影响接近显著(P=0.059),交互作用也接近显著(P=0.053)(表2)。区分不同光处理回归分析发现,加铝箔处理下,随硝酸钾处理浓度增加,鲜重线性增长显著(P<0.05),而全光照和黑暗下则不存在类似关系(表3)。

图5 不同浓度硝酸钾和光照处理对紫茎泽兰种子发芽率、芽长、根长、幼苗鲜重的影响
3 讨论
紫茎泽兰作为对我国造成巨大生态入侵的外来种之一,对其种子萌发与定植过程的研究一直是这一研究领域的热点[6]。刘伦辉等[1]提出紫茎泽兰种子的萌发和幼苗的生长要求比较稳定的光照及高温高湿条件;王文琪等[2]提出全光照对紫茎泽兰种子萌发有抑制作用,而在28%光照条件下萌发率最高。不同光照、干扰强度对紫茎泽兰种子萌发、幼苗定居和生长影响的研究发现,草山(地)上较小的干扰(直径15 cm,与牛、马蹄印大小相似)、较低光照(地表光强6%以下)能使紫茎泽兰种子萌发和幼苗定居[5]。在严格的室内控制条件下,证实了紫茎泽兰种子萌发对光照很敏感,有光与完全黑暗下发芽率相差30%—50%(P<0.05),但很微弱的光照就能够有效打破种子休眠,在透射率为0.23%的铝箔纸下的萌发率几乎与全光下发芽率相同。而且其它多种传统打破休眠的方法均不能代替光照,这些研究结果是对以往研究的补充。
紫茎泽兰种子需光萌发的特性,使得种子在土壤里面时,由于没有光照而形成数量可观的种子库,而一旦表土破坏,外界光照能够刺激种子大量萌发,形成入侵。可见紫茎泽兰种子的需光萌发特性,导致种子库为其在新生境大规模快速入侵打下了关键性的伏笔。此外,紫茎泽兰的种子很小,千粒重仅有48 mg,这使其种子自身储藏能量有限,不利于其抵抗逆境。紫茎泽兰的需光萌发特性,使得其具有利用外界光照的能力,更好的促进幼苗根系和生物量的生长(表3)。因此,这种需光萌发以及幼苗生长受光照与环境因子交互影响的特性,可能在一定程度上弥补其种子小、能量少所带来的入侵劣势[6]。
关于需光萌发的原因,通过类比的方法,发现目前常用的常规打破休眠的方法都不能替代光照的作用(图2—图5,表2)。通过与本研究相似的方法,Hsiao & Quick[14]研究发现,莴苣(LactucaSativa)的需光萌发特性可以被硝酸盐以及聚乙二醇改变。Henig-sever等[15]研究发现火后针叶树种子需光萌发特性可能与硝态氮和氨态氮的配比有关。Mandak & Pysek[16]的研究发现,硝酸盐中的N素很可能是影响草本植物需光萌发的原因。紫茎泽兰种子需光萌发几乎不受温度、聚乙二醇、水杨酸、硝酸钾等的影响,说明这一入侵植物的需光萌发特性可能有别于常见的需光萌发种子。种子休眠具有多样性,处于静态的种子是指处于干燥状态或者不利条件下种子不能萌发的种子,与通常的种子休眠(具有正常活力的种子在适宜条件下——光、温、水和氧气不能萌发的现象),也被称作强迫休眠[12-13]。目前很多学者对紫茎泽兰种子是休眠还是静态颇有争议[1,2,6],结果说明,只要给予光照,种子就能够正常萌发,并与其它环境如干旱、N营养等无关。这一结果支持紫茎泽兰种子属于种子静态这一结论。
虽然不同常规处理方法不能代替光照促进种子萌发,但可促进萌发种子的生长发育,甚至存在与光照的交互作用。一般来讲,干旱处理促进根系伸长而施加N肥,能提高生物量产量。紫茎泽兰种子受光处理直接影响聚乙二醇引发的根系生长的效果(没有光照下不起作用)、以及硝态氮引起的生物量增加等(过高光和没有光均不起作用)(表3)。于洋等[17]和王晓多等[18]已经报道了光照显著影响绒毛番龙眼种子(Pometiatomentosa)、豌豆种子(PisumSativum)光照显著影响幼苗生长特征的结果,结果进一步证实光照可以直接影响某些环境因子(干旱和养分供应)对紫茎泽兰幼苗生物量的调节作用。幼苗初期是生长最为脆弱的时期,对紫茎泽兰这些特征的揭示,有助于生态控制这一入侵植物以及从机理上揭示其入侵原因。紫茎泽兰作为喜光植物,其种子萌发和幼苗生长发育均需要一定的光照条件,而在林冠对光照的郁蔽影响下,紫茎泽兰是难以完成个体发育,因此需要采取相应的措施降低林下的光照度,进而阻滞紫茎泽兰种子的高萌发率,抑制其种群的进一步蔓延。目前较为有效的措施是增加森林高大植被的密度或者防止因毁林开荒造成的植被破坏为紫茎泽兰提供宜生地。
4 结论
本文通过对紫茎泽兰种子进行全光照、铝箔纸遮光及黑暗处理与4种不同打破种子休眠方法,即低温处理、水杨酸、聚乙二醇、硝酸钾处理的交互作用观察紫茎泽兰种子萌发特性及与光照影响的关系。结果表明:铝箔纸覆盖与全光照条件下种子的萌发率差异不显著(P>0.05),证明种子具有需光萌发的特性。而不同处理代替光照打破种子萌发的可能性很小,但其显著影响了某些发芽幼苗的生长特征。通过对紫茎泽兰种子进行光处理和不同浓度水杨酸和聚乙二醇处理的交互作用发现:水杨酸处理对种子的幼苗鲜重具有显著影响 (P<0.05)。其中在全光照条件下,0.05 mmol/L处理效果最明显,较对照提高39%;而根长在全光照条件下,经5%、10%和15%的聚乙二醇处理,分别比对照提高48%、45%、64%;方差分析也表明经过不同浓度的聚乙二醇处理,根长差异达到显著水平(P<0.05)。
[1] Liu L H, Xie S C, Zhang J H. Studies on the distribution, harmfulness and control ofEupatoriumadenophorumspreng. Acta Ecologica Sinica, 1985, 5(1): 1-6.
[2] Wang W Q, Wang J J, Zhao Z M. Seed population dynamics and germination characteristics ofEupatoriumadenophorum. Chinese Journal of Applied Ecology, 2006, 17(6): 982-986.
[3] Benech-Arnold R L, Snchez R A, Forcella F, Kruk B C, Ghersa C M. Environmental control of dormancy in weed seed banks in soil. Field Crops Research, 2000, 67(2): 105-122.
[4] Silvertown J W, Charlesworth D, Deborah C. Introduction to Plant Population Biology. Oxford: Blackwell Publishing, 2003: 240-240.
[5] Niu Y F, Feng Y L, Xie J L, Luo F C. Effects of disturbance intensity on seed germination, seedling establishment and growth ofAgeratinaadenophor. Guihaia, 2011, 31(6):795-800.
[6] Xu H N, Wang W J, Yu X Y, He H S, Cuan Y, Zu Y G. The difference in light-demanding germination features of seeds between invasive and noninvasive alien plants within composite. Acta Ecologica Sinica, 2010, 30(13): 3433-3440.
[7] Chen H, Zhang S, Cao M. Effects of light and temperature on seed germination ofFicushispidain Xishuangbanna, southwest China. Journal of Plant Ecology, 2008, 32(5): 1084-1090.
[8] Yuan Y B, Cao Z Z. The role of salicylic acid in plants. Chinese Bulletin of Botany, 1994, 11(3): 1-9.
[9] Michael B E, Kaufaman M R. The osmotic potential of polyethylene glycol 6000. Plant Physiology, 1973, 51(5): 914-916.
[10] Stewart R R C, Bewley J D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Physiology, 1980, 65(2): 245-248.
[11] Hilton J R. The Influence of light and potassium nitrate on the dormancy and germination ofAvenafatuaL. (WildOat) Seed and its ecological significance. New Phytologist, 1984, 96(1): 31-34.
[12] Yang Q H, Ye W H, Song S Q, Yin S H. Summarization on causes of seed dormancy and dormancy polymorphism. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(5): 837-843.
[13] Baskin, J M, Baskin. A classification system for seed dormancy. Seed Science Research, 2004, 14(1):1-16.
[14] Hsiao A I, Quick W A. The roles of inorganic nitrogen salts in maintaining phytochrome-and gibberellin A3-mediated germination control inSkotodormantlettuceSeeds. Journal of Plant Growth Regulation, 1996, 15(4): 159-165.
[15] Henig-Severa N, Eshelb A, Ne′emana G. Regulation of the germination ofAleppopine(Pinushalepensis) by nitrate, ammonium, and gibberellin, and its role in post-fire forest regeneration. Physiologia Plantarum, 2000, 108(4): 390-397.
[16] Mandák B, Pyšek P. The effects of light quality, nitrate concentration and presence of bracteoles on germination of different fruit types in the heterocarpousAtriplexsagittata. Journal of Ecology, 2001, 89(2): 149-158.
[17] Yu Y, Cao M, Zheng L, Sheng C Y. Effects of light on seed germination and seedling establishment of a tropical rainforest canopy tree, Pometia tomentosa. Journal of Plant Ecology, 2007, 31(6): 1028-1036.
[18] Wang X D, Lu Y Z, Yang J Y. Effects of Soaking with salicylic acid on seeds germination and seedlings growth of Pea (PisumSativum). Seed, 2007, 26(8): 42-44.
参考文献:
[1] 刘伦辉,谢寿昌, 张建华.紫茎泽兰在我国的分布、危害与防除途径的探讨. 生态学报, 1985, 5(1): 1-6.
[2] 王文琪,王进军, 赵志模.紫茎泽兰种子种群动态及萌发特性. 应用生态学报, 2006, 17(6): 982-986.
[5] 牛燕芬,冯玉龙,谢建磊,罗富成.干扰强度对群落中紫茎泽兰种子萌发、幼苗定居和生长的影响. 广西植物, 2011,31(6): 795-800.
[6] 许慧男,王文杰,于兴洋,贺海升,关宇,祖元刚.菊科几种入侵和非入侵植物种子需光发芽特性差异. 生态学报, 2010, 30(13): 3433-3440.
[7] 陈辉,张霜,曹敏.光和温度对西双版纳地区先锋树种对叶榕种子萌发的影响. 植物生态学报, 2008, 32(5): 1084-1090.
[8] 原永兵,曹宗巽.水杨酸在植物体内的作用. 植物学通报, 1994, 11(3): 1-9.
[12] 杨期和,叶万辉,宋松泉,殷寿华.植物种子休眠的原因及休眠的多形性. 西北植物学报,2003,23(5): 837-843.
[17] 于洋,曹敏,郑丽,盛才余.光对热带雨林冠层树种绒毛番龙眼种子萌发及其幼苗早期建立的影响. 植物生态学报, 2007, 31(6): 1028-1036.
[18] 王晓多,陆远柱,杨九英.水杨酸浸种对豌豆种子萌发及幼苗生长的影响. 种子, 2007, 26(8): 42-44.
