寒温针叶林土壤呼吸作用的时空特征
2013-12-20贾丙瑞周广胜蒋延玲殷晓洁胡天宇
贾丙瑞,周广胜,2,* ,蒋延玲,王 宇,殷晓洁,胡天宇
(1.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;2.中国气象科学研究院,北京 100081)
北方林又称泰加林,分布于45°—70°N之间的寒温带地区,横贯欧亚和北美大陆的北部。北方林向南延伸到我国大兴安岭北部,即我国的寒温针叶林。作为我国寒温针叶林的主要植被类型,兴安落叶松林面积占全国森林面积的30%,被认为在碳汇中占据重要份额[1]。北方林是地球上第二大陆地生物群区,来自地面植被观测、大气CO2和O2浓度监测、卫星遥感信息应用、生态和大气模型模拟等方面的研究均表明,北半球中高纬度的森林生态系统是一个巨大的碳汇,固定了全球碳循环中大部分“未知碳汇”[2-3]。
彭少麟等[4]研究表明,土壤碳大量积累并不是来自大的输入量,而是其它因素(如低温)对分解者的限制起主导作用。北方林生态系统地处寒温带气候区,因此,北方林生态系统拥有全球最大的土壤碳库,约占全球土壤碳库总量的23.4%[5]。已有研究表明,温度是寒温针叶林生态系统土壤呼吸作用的主要调控因子[6-11]。王庆丰等[12]对移栽自不同纬度的兴安落叶松研究表明,土壤呼吸作用对温度的敏感性随纬度升高而增加;其他研究也发现,寒冷气候区土壤呼吸作用对温度升高更加敏感[13-15]。因此,北方林生态系统因其巨大的土壤碳库和对温度变化的响应极为敏感而备受关注。我国大兴安岭林区近50年来气温呈显著增加趋势[16-18]。如果全球持续变暖,寒温针叶林生态系统潜在的巨大碳库将可能成为大气CO2的重要来源。因此,寒温针叶林在气候变化与全球碳收支中具有十分重要的作用[19]。
土壤呼吸作用作为土壤碳主要输出途径和重要的大气CO2源,对其精确测定已成为全球变化研究中的关键问题之一。目前,土壤呼吸作用野外原位测定通常依据观测样方大小不同随机选取3—5个观测样点[7,12,20-24],少数研究设置更多的观测样点数[9,25]。已有研究表明,寒温针叶林生态系统土壤呼吸作用空间变动性很大[8,10,20,26-28]。土壤呼吸作用的空间变化增加了其测定的不确定性,为准确估算土壤呼吸作用,必须解决土壤呼吸作用小尺度上的空间变异性[29]。同时,土壤呼吸作用时空动态变化特征及其驱动因子研究对于预测生态系统对气候变化的响应至关重要[8,22]。本研究对大兴安岭原始寒温针叶林生态系统土壤呼吸作用的日、季动态特征及其影响因子进行了分析,并比较同一区域不同观测样方个数土壤呼吸作用的变异性及其取样代表性,为深入研究寒温针叶林生态系统碳动态特征和准确估算土壤碳排放量提供科学依据与基础资料。
1 材料和方法
1.1 区域概况
本研究在中国科学院植物研究所中国北方林森林生态系统定位研究站(51°47'N,123°01'E,海拔773 m)进行。该站位于我国最大的寒温针叶林保护区——黑龙江呼中国家级自然保护区内,属寒温带大陆性季风气候。年均气温-4.4℃,在-2.7— -5.3℃之间波动。降水量为481.6 mm,主要集中在6—8月,无霜期80—100 d。棕色针叶林土是该地区最具代表性的土壤类型,地貌类型为大兴安岭北部石质中低山山地,坡度平缓,一般坡度在15°以下。地带性植被类型为寒温针叶林,以兴安落叶松(Larix gmelinii)为单优势树种,并混有少量白桦(Betula platyphylla),平均树高15 m,郁闭度0.52,林下灌木主要有杜香(Ledum palustre)、越桔(Vaccinium vitis-idaea)等。
1.2 研究方法
采用便携式光合作用测量系统Li-6400(LI-COR Inc.,USA)配合土壤气室(Li-6400-09)测定土壤呼吸作用,土壤呼吸作用测定前一天将10 cm直径PVC环插入土壤表面下2 cm,保持不动,并剪掉植被地上部分,共设置9个重复,每个重复连续测定3次,取其平均值。2006年和2007年6月至10月,每月进行1次土壤呼吸作用测定,其中2006年7月5—6日和8月10—11日、2007年7月22—23日进行了土壤呼吸作用全天日动态测定,其他为白天(6:00—18:00)日动态测定,每隔2 h测定1次。为了解土壤呼吸作用的空间异质性及其取样代表性,选择平坦的样地布设40个观测样点,分为5列8行,列和行间距均为2 m,2009年7月4日分别于3:00、7:00和11:00测定土壤呼吸作用。测定完成后收集40个观测点的地表凋落物,并挖取每个样点0—30 cm土壤,用0.5 mm孔径筛冲洗分别拣出土层中的粗根(>2 mm)和细根(≤2 mm),带回实验室烘干(75℃)称重。气象观测塔距观测样地约100 m,每半小时自动记录1次,包括1.5 m气温(HMP45C,Vaisala,Finland);表层、5 cm 和 10 cm 土壤温度(107 L,Campbell Scientific Inc.,USA)等。
1.3 统计分析
运用SPSS统计软件双变量相关分析中的Pearson相关分析,进行土壤呼吸作用与不同层次温度在日、季尺度的相关性比较。为求取土壤呼吸作用、根系和凋落物40个观测样点不同取样个数的变异性,分别以1个样点观测值以及相邻2个、3个,…,10个样点平均值计算变异系数,用于描述样地空间异质性造成的取样变异性,变异系数=标准差/平均值×100。
2 结果和分析
2.1 气温与地温的日、季动态变化
寒温针叶林生态系统6月至10月1.5 m气温日动态呈单峰型曲线(图1),最高值出现在午后14:00左右,最低值通常出现在 4:30 左右;6—10 月月均温分别为 14.47、15.81、16.15、5.29、0.19 ℃,6 月、7 月和 8 月气温逐渐升高,9月和10月气温则大幅降低且夜间气温降到0℃以下。图1给出了寒温针叶林生态系统6—10月5 cm土壤温度日动态变化,其中6—9月呈单峰型曲线,10月变幅很小,基本在0℃上下波动。6—9月5 cm土壤温度最高值出现在16:00左右,滞后于气温2 h;最低值出现在6:00左右,滞后于气温约1.5h。6—8月1.5 m月平均气温比5 cm土壤温度高约5℃,9月和10月两者月均温则接近。
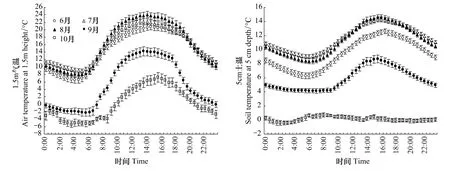
图1 6—10月1.5 m气温和5 cm土温月平均日动态Fig.1 Mean diurnal dynamics of air temperature at 1.5 m height and soil temperature at 5 cm depth from June to October.Dots are monthly means with standard error bars
2.2 土壤呼吸作用的日、季动态及其影响因子
图2给出了2006年7月5—6日和8月10—11日、2007年7月22—23日土壤呼吸作用日动态变化,一天内土壤呼吸作用均呈单峰型曲线变化,高峰值出现在16:00左右,最低值出现在6:00左右,日变动幅度分别为 2.88、5.31 μmol CO2m-2s-1和 4.42 μmol CO2m-2s-1,与 5 cm 土壤温度日动态变化趋势相近,滞后于气温日动态变化(图1)。10:00和22:00的土壤呼吸作用最接近一天的平均值。因此,可以选择10:00左右测定的土壤呼吸作用平均值作为日平均值。姚玉刚等[30]对热带雨林研究也表明,9:00—11:00土壤呼吸作用接近日平均值。
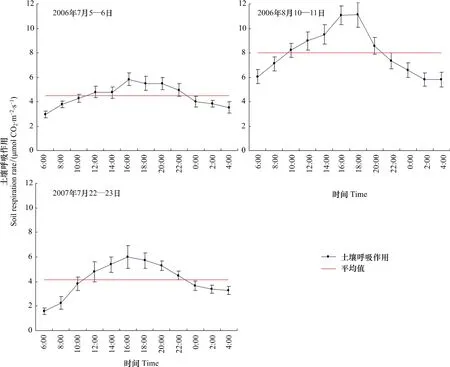
图2 2006年和2007年土壤呼吸作用日动态变化Fig.2 Diurnal dynamics of soil respiration rate on Jul.5—6,2006,Aug.10—11,2006 and Jul.22—23,2007
图3给出了2006年和2007年生长季节6—10月土壤呼吸作用动态变化,6月份随着天气转暖和植被生长土壤呼吸作用逐渐增大,8月份气温最高时土壤呼吸作用也达到最大值,2006年和2007年分别为8.19μmol CO2m-2s-1和6.89 μmol CO2m-2s-1,随后 9 月份气温迅速下降土壤呼吸作用也降低,到10月份气温波动于0℃左右,寒温针叶林生态系统只维持微弱的土壤呼吸作用,约为 0.61 μmol CO2m-2s-1。2006 年和 2007 年 6—10月土壤呼吸作用季节动态变化幅度分别为7.38 μmol CO2m-2s-1和 6.48 μmol CO2m-2s-1。兴安落叶松人工林土壤呼吸作用与本研究结果相近,6—10月分别为 4.30、4.88、5.88、3.47、2.76 μmol CO2m-2s-1,最大值出现在8月份[31]。小兴安岭兴安落叶松沼泽林[24]和寒温带落叶林[21]也表现出相似的季节变化特征。
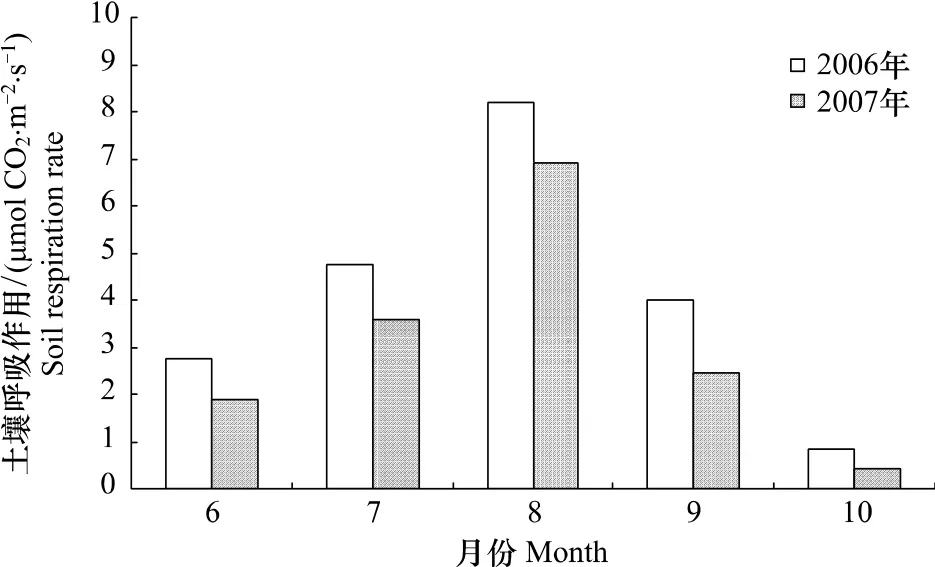
图3 2006年和2007年土壤呼吸作用季节动态变化Fig.3 Seasonal dynamics of soil respiration rate in 2006 and 2007
土壤呼吸作用平均值与气温和3个层次土壤温度相关性分析表明(表1),在日尺度上,土壤呼吸作用与3个层次土壤温度的相关性均达到显著和极显著水平,Pearson相关系数(R)在0.737以上;土壤呼吸作用与气温的相关系数为0.502—0.868,土壤呼吸作用与土壤温度的相关性好于气温。在季节尺度上,2006年土壤呼吸作用与土壤温度和气温的相关性均达到极显著水平,其中与3个层次土壤温度的相关系数在0.846以上,而与气温的相关系数为0.687;2007年土壤呼吸作用与3个层次土壤温度相关性均达到极显著水平且相关系数在0.615以上,与气温的相关性达到显著水平,相关系数为0.513。因此,不论在日尺度上还是季节尺度上,寒温针叶林生态系统土壤呼吸作用主要受土壤温度调控,尤其是5 cm土壤温度。对兴安落叶松人工林[32]和兴安落叶松沼泽林[24]研究均表明,土壤温度比气温更能准确地反映土壤呼吸作用的动态变化。
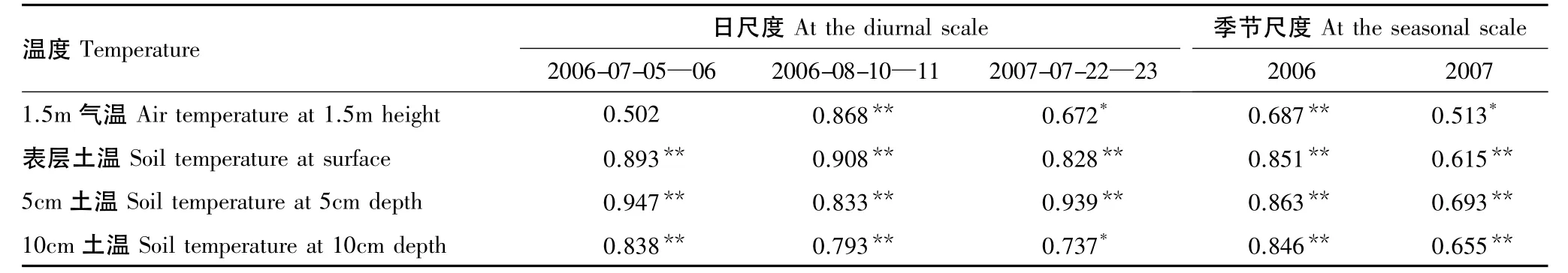
表1 土壤呼吸作用与不同层次温度的Pearson相关系数(R)Table 1 Pearson correlation coefficients(R)of soil respiration rate with temperature at different layers
2.3 土壤呼吸作用的空间分布特征
基于3:00、7:00和11:00以2 m间距(5列8行)测定的40个观测样点土壤呼吸作用,分别以1个样点观测值以及相邻2个、3个、…、10个样点平均值计算的土壤呼吸作用变异系数如图4。1个观测样点变异系数即40个样方在2 m间距(5列8行)范围内的空间变异性,3:00、7:00和11:00分别为35.5%、27.6%和23.0%,说明该研究地点以10 cm直径圆形样方测定土壤呼吸作用时,样方间存在较大的空间变异性。3:00、7:00 和11:00 变动幅度分别为5.97、4.96 μmol CO2m-2s-1和5.84 μmol CO2m-2s-1。随着观测样点个数增加,土壤呼吸作用变异系数呈逐渐递减趋势,3:00由35.5%减少到19.8%,7:00由27.6%减少到11.1%,11:00由23.0%减少到9.1%。3个观测样点时土壤呼吸作用变异系数迅速降低,3:00、7:00和11:00变异系数分别为27.0%、16.8%和 13.0%;6 个观测样点时,3:00、7:00 和 11:00 变异系数分别为 22.8%、11.1%和 11.8%,基本接近7—10个观测样点时的最小变异系数。因此,以10 cm直径圆形样方测定时,6个观测样点基本能够反映寒温针叶林的土壤呼吸作用变化情况。
对于40个样方的根系和凋落物来说,与土壤呼吸作用表现出相似的空间变异性(图4)。其中粗根的空间变异性最大,达到103.8%;总根系和凋落物次之,分别为60.3%和47.1%;细根空间变异性最小,为32.1%。随着观测样点个数增加,变异系数均逐渐降低,且降低幅度按粗根、总根、凋落物和细根顺序依次递减,3个观测样点时变异系数分别为 53.1%、33.4%、34.1%和 18.2%,6个观测样点时变异系数分别为 37.8%、27.3%、25.7%和16.7%,与7—10个观测样点时的变异系数基本持平。
3:00、7:00和11:00土壤呼吸作用与细根、粗根、总根及凋落物的相关性分析表明(图5),3个时段土壤呼吸作用均与细根的相关性最好,相关系数(R2)分别为0.351、0.396和0.322,说明该区域土壤呼吸作用的空间变异性主要由于细根差异引起,粗根和凋落物的影响并不大。
3 讨论
本研究土壤呼吸作用最大值出现在夏季生长旺盛期8月份气温最高时,2006年和2007年分别为8.19 μmol CO2m-2s-1和 6.89 μmol CO2m-2s-1。兴安落叶松林 4 大类常见微生物(细菌、放线菌、真菌、自生固氮菌)最大数量也同样出现在夏季[33]。10月份气温波动于0℃左右,寒温针叶林生态系统只维持微弱的土壤呼吸作用,约为 0.61 μmol CO2m-2s-1,接近于解冻前兴安落叶松林的土壤呼吸作用平均值(0.5 μmol CO2m-2s-1)[12]。分布于其他纬度的北方林生态系统土壤呼吸作用变动范围为 0.4—6.9 μmol CO2m-2s-1(48°13'N)[10]、0.6—9.3 μmol CO2m-2s-1(53°42'N)[6]、0.1—7 μmol CO2m-2s-1(53°59'N)[9],本研究结果基本接近这些范围。
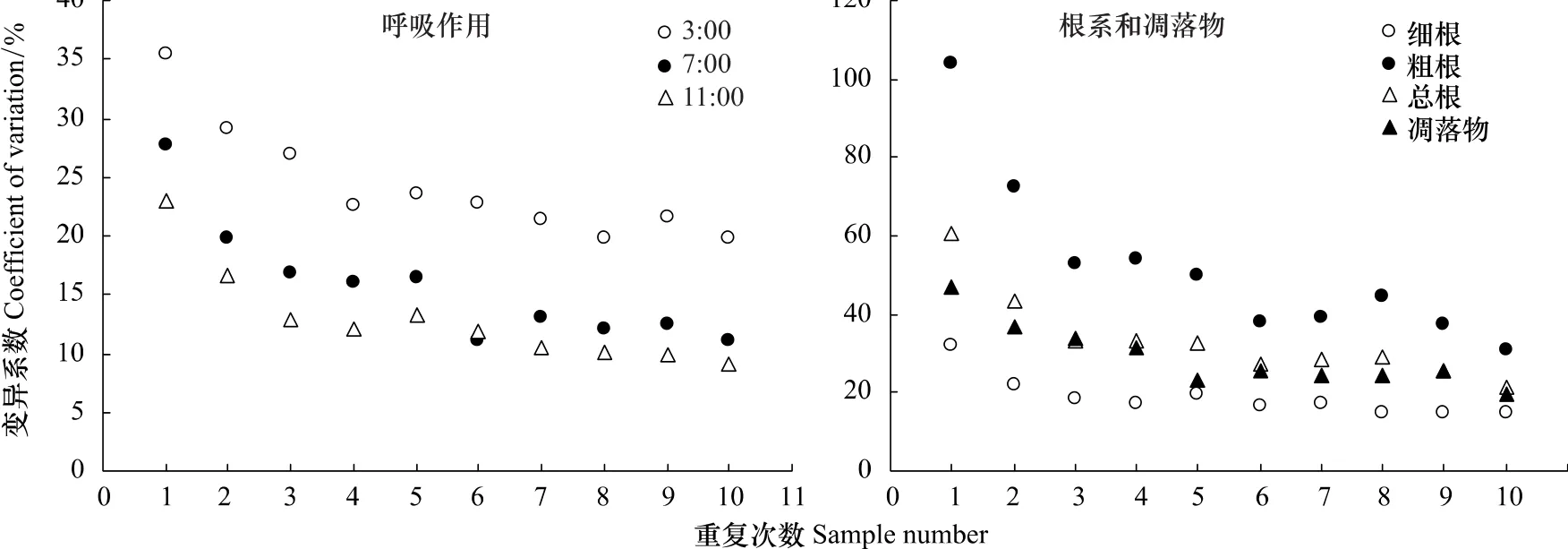
图4 土壤呼吸作用变异系数、根系和凋落物变异系数与重复次数的关系Fig.4 Relationship of sample number with coefficients of variation for soil respiration rate,root and litter

图5 土壤呼吸作用与细根、粗根、总根及凋落物的关系Fig.5 Relationships of soil respiration rate with fine root,coarse root,total root and litter
本研究区域40个样方土壤呼吸作用的空间变异幅度为4.96—5.97μmol CO2m-2s-1,大于或接近土壤呼吸作用日动态变化幅度(2.88—5.31 μmol CO2m-2s-1),而小于土壤呼吸作用季节动态变化幅度(6.48—7.38 μmol CO2m-2s-1),与Buchmann[20]研究结果相近。以10 cm直径圆形样方测定土壤呼吸作用时,40个样点在3:00、7:00和11:00 3个时间段的变异系数分别为35.5%、27.6%和23.0%,说明寒温针叶林生境下用10 cm直径圆形样方取样时各样方间的空间变异性较大,通过增加观测样点个数可使土壤呼吸作用变异系数迅速降低,6个观测样点基本能够反映寒温针叶林的土壤呼吸作用变化情况。Fang等[34]佛罗里达湿地松人工林研究表明,在25×25 m样地内12个样方(直径7.1cm)的土壤呼吸作用变异系数高达55%。Buchmann[20]对挪威云杉林同样研究表明,夏季生长旺盛期土壤呼吸作用最大时,同一研究地点不同观测样方(直径10 cm)间土壤呼吸作用的变异系数最高,达40%。Yim等[27]对落叶松人工林研究表明,在30 m×30 m样地内50个样方(直径12.6 cm)两个观测日期的土壤呼吸作用变异系数平均值为28%。Rodeghiero和Cescatti[28]对两个寒温针叶林生态系统研究表明,挪威云杉林和银冷杉林观测样方(直径10 cm)间土壤呼吸作用变异系数平均值分别为41.5%和28.9%。另外,土壤呼吸作用空间变异性与其观测样方的大小有关[27,35]。
对于相对均一的生态系统来说,温度和水分通常是影响土壤呼吸作用时间动态变化的主导因素,而根系、凋落物、土壤有机质和孔隙度等反映了土壤呼吸作用的空间变化特征[27,34]。土壤呼吸作用主要由根系呼吸和微生物呼吸组成,森林生态系统根系呼吸所占比例平均为45%—50%[36]。细根分布不均是该区域土壤呼吸作用空间变动的主要因素之一,能解释土壤呼吸作用32.2%—39.6%变化。究其原因,根系呼吸速率随直径增加而呈明显下降趋势[37-38],细根呼吸速率大于粗根。Cook等[39]研究也表明,随着与树干距离的增加土壤呼吸作用呈指数形式递减,主要与细根密度的减少有关。
[1]Gower ST,Richards J H.Larches:deciduous conifers in an evergreen world.Bioscience,1990,40(11):818-826.
[2]Fang J Y,Piao SL,Zhao SQ.The carbon sink:the role of the middle and high latitudes terrestrial ecosystems in the northern hemisphere.Acta Phytoecologica Sinica,2001,25(5):594-602.
[3]Yu G R,Li H T,Wang SQ.Global Change,Carbon Cycle and Storage in Terrestrial Ecosystem.Beijing:China Meteorological Press,2003.
[4]Peng SL,Li Y L,Ren H,Zhao P.Progress in research on soil respiration under the global change.Advances in Earth Sciences,2002,17(5):705-713.
[5]IPCC.Climate change 2001:The Scientific Basis.Intergovernmental Panel on Climate Change.2001.http://www.grida.no/climate/ipcc_tar/wg1/index.htm.
[6]Russell C A,Voroney R P.Carbon dioxide efflux from the floor of a boreal aspen forest.I.Relationship to environmental variables and estimates of C respired.Canadian Journal of Soil Science,1998,78(2):301-310.
[7]Morén A S,Lindroth A.CO2exchange at the floor of a boreal forest.Agricultural and Forest Meteorology,2000,101(1):1-14.
[8]Rayment M B,Jarvis P G.Temporal and spatial variation of soil CO2efflux in a Canadian boreal forest.Soil Biology and Biochemistry,2000,32(1):35-45.
[9]Swanson R V,Flanagan L B.Environmental regulation of carbon dioxide exchange at the forest floor in a boreal black spruce ecosystem.Agricultural and Forest Meteorology,2001,108(3):165-181.
[10]Khomik M,Arain M A,McCaughey J H.Temporal and spatial variability of soil respiration in a boreal mixedwood forest.Agricultural and Forest Meteorology,2006,140(1/4):244-256.
[11]Karhu K,Fritze H,Tuomi M,Vanhala P,Spetz P,Kitunen V,Liski J.Temperature sensitivity of organic matter decomposition in two boreal forest soil profiles.Soil Biology and Biochemistry,2010,42(1):72-82.
[12]Wang Q F,Wang C K,Tan L H.Vernal soil respiration of Larix gmelinii Rupr.forests transplanted from a latitudinal transect.Acta Ecologica Sinica,2008,28(5):1883-1892.
[13]Lloyd J,Taylor JA.On the temperature dependence of soil respiration.Functional Ecology,1994,8(3):315-323.
[14]Peng SS,Piao SL,Wang T,Sun J Y,Shen Z H.Temperature sensitivity of soil respiration in different ecosystems in China.Soil Biology and Biochemistry,2009,41(5):1008-1014.
[15]Liao Y,Yang Z F,Xia X Q,Jiang H C.Research on temperature sensitivity of soil respiration and different active organic carbon fractions of Qinghai-Tibet Plateau permafrost.Earth Science Frontiers,2011,18(6):85-93.
[16]Zhou M,Yu X X,Feng L.Analysis on the driving force of the permafrost degeneration in Daxinganling.Journal of Arid Land Resources and Environment,2002,16(4):44-47.
[17]Sun F H,Yang X Q,Lu S,Yang S Y.The contrast analysis on the average and extremum temperature trend in northeast China.Scientia Meteorologica Sinica,2006,26(2):157-163.
[18]Wang Y,Zhou G S,Jia B R,Na JH.Climate change characteristics in Chinese boreal forest region from 1954 to 2005.Chinese Journal of Applied Ecology,2008,19(5):942-948.
[19]Jia B R,Zhou G S.Advance in the studies of the response of boreal forest to climate change.Advances in Earth Sciences,2009,24(6):668-674.
[20]Buchmann N.Biotic and abiotic factors controlling soil respiration rates in Picea abies stands.Soil Biology and Biochemistry,2000,32(11/12):1625-1635.
[21]Lee M S,Nakane K,Nakatsubo T,Koizumi H.Seasonal changes in the contribution of root respiration to total soil respiration in a cool-temperate deciduous forest.Plant and Soil,2003,255(1):311-318.
[22]Li H J,Yan J X,Yue X F,Wang M B.Significance of soil temperature and moisture for soil respiration in a Chinese mountain area.Agricultural and Forest Meteorology,2008,148(3):490-503.
[23]Yan JX,Qin Z D,Zhang Y H,Li H J.Effect of soil temperature and moisture on soil CO2efflux in a Pinus tabulaeformis forest.Acta Ecologica Sinica,2009,29(12):6366-6376.
[24]Mu C C,Cheng W,Sun X X,Wu Y X.Seasonal variation of emission fluxes of CO2,N2Oand CH4from Larix gemlinii swamps soils in Xiaoxing'an Mountains of China.Scientia Silvae Sinicae,2010,46(7):7-15.
[25]Zhang J B,Shangguan T L,Meng Z Q.Changes in soil carbon flux and carbon stock over a rotation of poplar plantations in northwest China.Ecological Research,2011,26(1):153-161.
[26]Thierron V,Laudelout H.Contribution of root respiration to total CO2efflux from the soil of a deciduous forest.Canadian Journal of Forest Research,1996,26(7):1142-1148.
[27]Yim M H,Joo SJ,Shutou K,Nakane K.Spatial variability of soil respiration in a larch plantation:estimation of the number of sampling points required.Forest Ecology and Management,2003,175(1/3):585-588.
[28]Rodeghiero M,Cescatti A.Spatial variability and optimal sampling strategy of soil respiration.Forest Ecology and Management,2008,255(1):106-112.
[29]Han G X,Zhou G S.Review of spatial and temporal variations of soil respiration and driving mechanisms.Chinese Journal of Plant Ecology,2009,33(1):197-205.
[30]Yao Y G,Zhang Y P,Yu G R,Sha L Q,Deng Y,Tan Z H.Representative time selection analysis on daily average value of soil respiration in a tropical rain forest.Journal of Nanjing Forestry University:Natural Science Edition,2011,35(4):74-78.
[31]Wang X W.Study on Key Process of Carbon Cycle in Larix Gmelinii Plantation[D].Harbin:Northeast Forestry University,2006:12-43.
[32]Wang C K,Yang JY.Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests.Global Change Biology,2007,13(1):123-131.
[33]Feng B P,Gao R H,Zhang Q L,Zhao Q M.Study on dynamic variation of soil microorganism under different management modes in Larix Gmelini forest.Journal of Inner Mongolia Agricultural University:Natural Science Edition,2009,30(4):74-79.
[34]Fang C,Moncrieff JB,Gholz H L,Clark K L.Soil CO2efflux and its spatial variation in a Florida slash pine plantation.Plant and Soil,1998,205(2):135-146.
[35]Davidson E A,Savage K,Verchot L V,Navarro R.Minimizing artifacts and biases in chamber-based measurements of soil respiration.Agricultural and Forest Meteorology,2002,113(1/4):21-37.
[36]Hanson P J,Edwards N T,Garten CT,Andrews JA.Separating root and soil microbial contributions to soil respiration:a review of methods and observations.Biogeochemistry,2000,48(1):115-146.
[37]Chapman SB.Some interrelationships between soil and root respiration in lowland Calluna heathland in southern England.The Journal of Ecology,1979,67(1):1-20.
[38]Jia SX,Zhao Y L,Ding G Q,Sun Y,Xu Y,Wang Z Q.Relationship among fine-root morphology,anatomy,tissue nitrogen concentration and respiration in different branch root orders in Larix gmelinii and Fraxinus mandshurica.Chinese Bulletin of Botany,2010,45(2):174-181.
[39]Cook F J,Thomas S M,Kelliher F M,Whitehead D.A model of one-dimensional steady-state carbon dioxide diffusion from soil.Ecological Modelling,1998,109(2):155-164.
参考文献:
[2]方精云,朴世龙,赵淑清.CO2失汇与北半球中高纬度陆地生态系统的碳汇.植物生态学报,2001,25(5):594-602.
[3]于贵瑞,李海涛,王绍强.全球变化与陆地生态系统碳循环和碳蓄积.北京:气象出版社,2003.
[4]彭少麟,李跃林,任海,赵平.全球变化条件下的土壤呼吸效应.地球科学进展,2002,17(5):705-713.
[12]王庆丰,王传宽,谭立何.移栽自不同纬度的落叶松 (Larix gmelinii Rupr.)林的春季土壤呼吸.生态学报,2008,28(5):1883-1892.
[15]廖艳,杨忠芳,夏学齐,蒋宏忱.青藏高原冻土土壤呼吸温度敏感性和不同活性有机碳组分研究.地学前缘,2011,18(6):85-93.
[16]周梅,余新晓,冯林.大兴安岭林区多年冻土退化的驱动力分析.干旱区资源与环境,2002,16(4):44-47.
[17]孙凤华,杨修群,路爽,杨素英.东北地区平均、最高、最低气温时空变化特征及对比分析.气象科学,2006,26(2):157-163.
[18]王宇,周广胜,贾丙瑞,那济海.1954—2005年中国北方针叶林分布区的气候变化特征.应用生态学报,2008,19(5):942-948.
[19]贾丙瑞,周广胜.北方针叶林对气候变化响应的研究进展.地球科学进展,2009,24(6):668-674.
[23]严俊霞,秦作栋,张义辉,李洪建.土壤温度和水分对油松林土壤呼吸的影响.生态学报,2009,29(12):6366-6376.
[24]牟长城,程伟,孙晓新,吴云霞.小兴安岭落叶松沼泽林土壤CO2,N2O和CH4的排放规律.林业科学,2010,46(7):7-15.
[29]韩广轩,周广胜.土壤呼吸作用时空动态变化及其影响机制研究与展望.植物生态学报,2009,33(1):197-205.
[30]姚玉刚,张一平,于贵瑞,沙丽清,邓云,谭正洪.热带雨林土壤呼吸测定代表性时段研究.南京林业大学学报:自然科学版,2011,35(4):74-78.
[31]王秀伟.兴安落叶松人工林碳循环关键过程的研究[D].哈尔滨:东北林业大学,2006:12-43.
[33]冯保平,高润宏,张秋良,赵全民.不同经营方式下兴安落叶松林土壤微生物年季动态研究.内蒙古农业大学学报:自然科学版,2009,30(4):74-79.
[38]贾淑霞,赵妍丽,丁国泉,孙玥,许旸,王政权.落叶松和水曲柳不同根序细根形态结构、组织氮浓度与根呼吸的关系.植物学报,2010,45(2):174-181.
