美洲棘蓟马对不同蔬菜寄主的偏好性
2013-12-16石宝才宫亚军王泽华康总江马吉德魏书军
朱 亮,石宝才,宫亚军,王泽华,康总江,马吉德,魏书军,*
(1.北京市农林科学院植物保护环境保护研究所,北京 100097;2.华南农业大学昆虫学系,广州 510642)
美洲棘蓟马 Echinothrips americanus Morgan属缨翅目 Thysanoptera蓟马科 Thripidae针蓟马亚科Panchaetothripinae,原产于北美东部[1],目前已经扩散至欧洲和亚洲等地[2-4]。国内于2010年报道了该虫入侵我国北京和陕西省杨凌地区[5-7]。
Brodbeck等在研究缨翅目害虫营养生态学时发现这类害虫大部分种表现出取食的多样化[8],因此有必要深入研究这类昆虫对其寄主的选择习性,从而为其入侵风险评估提供重要资料。国内仅对缨翅目中西花蓟马的寄主研究较多,发现该虫既对花卉寄主具有选择性又对蔬菜寄主具有选择性[9-11]。Milne和Walter研究了烟蓟马Thrips tabaci Lindeman的寄主种类及其危害特征[12]。Varga和Fedor对美洲棘蓟马的嗜好寄主和非嗜好寄主进行了分类研究[13]。
目前发现美洲棘蓟马的寄主有48科106种[13],主要涉及的是观赏植物,适合的寄主有天南星科Araceae、大戟科Euphorbiaceae、夹竹桃科Apocynaceae等。该虫也可对温室作物造成危害[2],例如,美洲棘蓟马在荷兰危害温室甜椒,成为温室中的重要害虫[14]。然而,国内外尚未有关于美洲棘蓟马对蔬菜寄主的偏好性与适应性方面的系统报道。目前研究的美洲棘蓟马的蔬菜寄主仅仅为豆科Leguminosae中的四季豆Phaseolus vulgar、黄豆Glycine max和茄科Solanaceae中的青椒Capsicum anuum[13]。本文报道美洲棘蓟马对我国主要蔬菜作物的偏好性及产卵选择性,为该虫入侵我国后的风险评估与防控提供科学依据。
1 材料与方法
1.1 供试虫源与寄主植物
供试昆虫美洲棘蓟马来自北京市农林科学院植物保护环境保护研究所,于室内饲养在芹菜Apium graveolens上,连续饲养5代,温度25℃,相对湿度70%,光照16 L∶8 D[6]。供试寄主植物为豆科Leguminosae的豆角Vigna unguiculata和黄豆G.max,伞形科Apiaceae的胡萝卜Daucus carota和茴香Foeniculum vulgare,茄科Solanaceae的茄子 Solanum melongena、辣椒 Capsicum frutescens和番茄 Solanum lycopersicum,葫芦科Cucurbitaceae的黄瓜 Cucumis sativus和南瓜 Cucurbita moschata,十字花科 Brassicaceae的油菜 Brassica chinensis、小白菜Brassica campestris和甘蓝Brassica oleracea,紫苜蓿Medicago sativa作为非蔬菜类的对照寄主植物。试验所用13种寄主苗均为出土后1—2月的苗,苗高不超过0.3 m。其中除紫苜蓿和茴香叶片较小较多外,11种寄主均只有4—6片叶。
1.2 试验方法
1.2.1 美洲棘蓟马成虫对不同蔬菜寄主的寄主偏好性及产卵选择性
选用13种供试寄主植物的健壮无虫苗各1株,在网罩密封的养虫架(1.1 m ×1.1 m ×0.8 m)内围成一圈。4组重复中寄主植物的排列顺序不同,以随机方式排布,但同一科寄主植物不相邻,最小间隔距离为35 cm,且将紫苜蓿和茴香分布于其他植物之间,苗间距为20 cm。将美洲棘蓟马雌成虫100头接入同一片青椒C.annuum叶片上,再将该叶片放在13种供试寄主植物围成的圆圈的圆心位置,72 h后记录每株寄主植物上的成虫的总数量,作为单株寄主植物成虫分布量。美洲棘蓟马雌成虫死亡及逃逸的数量明显降低,4组重复试验中成虫死亡及逃逸的总和分别占到37%、17%、17%和16%,成虫剩余量分别为63、83、83和84头。由于青椒叶片表面有蜡质层,较为光滑,易于蓟马的扩散,且青椒叶片很快会干枯,对研究结果的影响较小。因此本研究以青椒叶片作为初始虫源的释放载体。
剔除各苗上的成虫,将各供试植物分开培养,每日记录不同寄主蔬菜上孵化的若虫数,直到无若虫出现为止,统计出每株寄主植物上若虫总数,用总若虫数作为总产卵量[11]。试验共重复4次,在温度(25±1)℃,相对湿度(70±5)%,密闭无光养虫室进行。养虫室在试验前经过灭菌、消毒和虫源处理。
1.2.2 美洲棘蓟马在不同蔬菜寄主上的发育历期及存活率
分别取5种寄主植物黄瓜、南瓜、油菜、黄豆和辣椒的叶10片,用棉花包住其叶柄放于塑料培养皿中,在棉花上加水保湿。分别挑美洲棘蓟马雌成虫30头于每个培养皿的叶片上,8 h后移去成虫。每日8:30和20:30在镜下各观察1次,将刚孵化的若虫分别移入相应寄主蔬菜叶片的新塑料培养皿中饲养,并记录每头若虫孵化时间。每个培养皿中仅放1头若虫。每天8:30和20:30各观察1次,定期更换叶片,记录美洲棘蓟马从1龄若虫到成虫羽化中各个阶段的时间,以及各个阶段刚开始时该虫的数量。每种蔬菜设置30个重复。最后计算未成熟期各个阶段的发育历期和存活率。各发育阶段存活率=发育阶段结束时虫量/发育阶段开始时虫量。整个未成熟期存活率=羽化的成虫数量/初始孵化1龄若虫的数量(卵期的存活率默认为100%)。试验在温度为(25±1)℃、相对湿度为(70±5)%、光周期为16 L:8 D的培养箱中进行。
1.3 数据统计分析
本研究中美洲棘蓟马成虫对不同蔬菜寄主的寄主偏好性和产卵选择性部分利用对数线性模型在DPS v13.5软件中对数据进行分析[15-16]。首先将13种寄主植物设定为变量因子A,将4个重复设定为变量因子B,从而组成结构为[A][B]的模型。然后在对各个因子最大似然估计进行卡方检验,最后通过参数估计得到各个寄主植物优势比的95%置信区间。当任意两个处理95%置信区间范围重合则差异不显著,反之差异显著。
利用DPS v13.5[15]对美洲棘蓟马在不同蔬菜寄主上的发育历期进行方差分析和多重比较 (Duncan氏新复极差测验法)。
2 结果与分析
2.1 美洲棘蓟马成虫对不同蔬菜寄主的寄主偏好性
美洲棘蓟马成虫在南瓜上分布量最大,平均每株寄主植物达到了23.5头(表1)。除南瓜以外,成虫分布量较大的还有油菜、黄瓜、辣椒和黄豆。以上5种寄主植物上的成虫分布量显著大于其他供试寄主植物。而茴香、紫苜蓿和甘蓝上的成虫分布量均为0,可见美洲棘蓟马不喜欢取食这3种植物。
2.2 美洲棘蓟马成虫对不同蔬菜寄主的产卵选择性
美洲棘蓟马产卵量较大的寄主植物有黄瓜、黄豆、油菜、辣椒和南瓜(表2),其中油菜上产卵量最大,每株达到了50.32粒卵。从统计检验的结果来看,美洲棘蓟马在以上5种寄主植物上产卵量显著高于除豆角以外的其他7种寄主植物。而美洲棘蓟马在胡萝卜、甘蓝和紫苜蓿上都没有产卵,说明美洲棘蓟马不喜好在这3种寄主植物上进行种群的繁衍。

表1 美洲棘蓟马成虫对不同蔬菜寄主的寄主偏好性Table 1 Host preference of Echinothrips americanus for different host vegetables
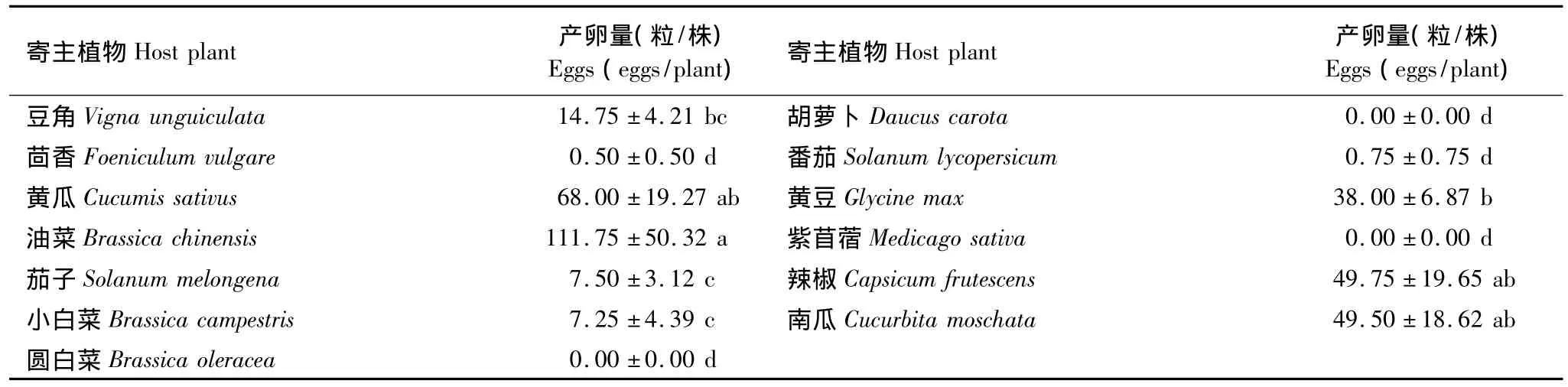
表2 美洲棘蓟马对不同蔬菜寄主的产卵选择性Table 2 Oviposition preference of Echinothrips americanus to different host vegetables
2.3 美洲棘蓟马寄主偏好性与产卵选择性的关系
在不同寄主植物上的美洲棘蓟马成虫分布量和产卵量基本一致(图1)。美洲棘蓟马成虫分布量大的寄主植物上其产卵量同样大。油菜上成虫分布量仅次于南瓜排名第二,而产卵量则是最大。黄瓜、黄豆、油菜、辣椒和南瓜上成虫分布量和产卵量均较大,说明美洲棘蓟马成虫较喜好这5种寄主植物。
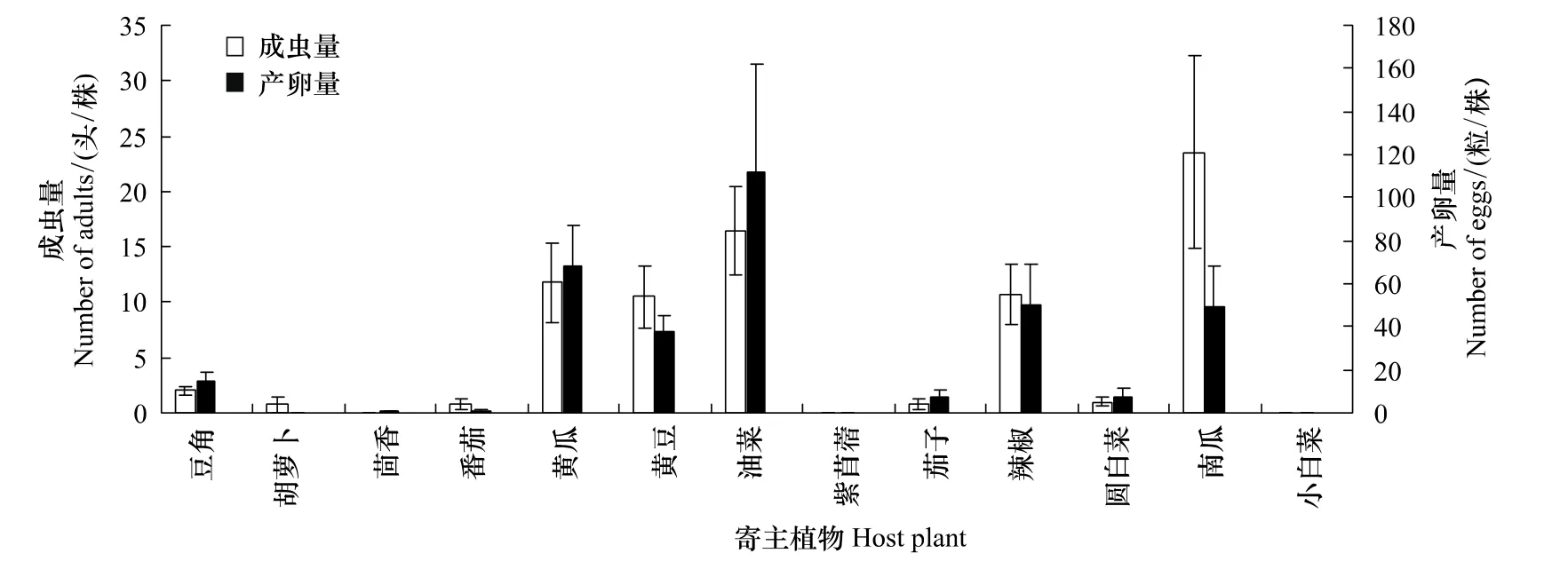
图1 美洲棘蓟马对于不同蔬菜寄主寄主偏好性与产卵选择性的关系Fig.1 Relationship between feeding preference and oviposition preference of Echinothrips americanus for different host vegetables
2.4 美洲棘蓟马在不同蔬菜寄主上的发育历期
在不同寄主植物上,美洲棘蓟马未成熟期的各个阶段发育历期各不相同(表3)。在不同的发育阶段中卵的发育历期最长,在6—9 d之间。油菜上卵期最短,黄豆上最长。该虫在南瓜上的1龄若虫期最短,平均在2.35 d,显著短于油菜和黄豆上1龄若虫期;在油菜上1龄若虫期最长,平均为3.04 d。该虫在黄豆、南瓜和辣椒上的2龄若虫期显著长于油菜,其中在黄豆上历期最长,其次是南瓜和辣椒,在油菜上最短。美洲棘蓟马预蛹期很短,各个寄主植物上的预蛹期都不到2 d。不同寄主植物上的预蛹期差异不显著,在黄瓜上预蛹期最长,平均达到了1.53 d,在辣椒上预蛹期最短,平均为1.21 d。蛹期比预蛹期发育历期长,但在不同寄主植物上的平均历期均在2 d以下。在辣椒上蛹期最长,平均为1.92 d,在黄豆上蛹期最短,平均为1.15 d;在黄豆上的蛹期显著短于黄瓜、南瓜、油菜和辣椒。
5种不同寄主植物上的美洲棘蓟马未成熟期的发育历期有所不同,但均在15—17 d之间。在黄瓜上发育最快,平均为15.09 d,在黄豆上发育最慢,平均为16.77 d。在黄瓜和油菜上的未成熟期显著短于辣椒和黄豆上,在南瓜上的未成熟期显著短于在黄豆上的历期。

表3 美洲棘蓟马在不同蔬菜上的发育历期Table 3 The developmental duration of Echinothrips americanus on different vegetables
2.5 美洲棘蓟马在不同蔬菜寄主上的存活率
在不同寄主植物上,美洲棘蓟马未成熟期的各个阶段存活率存在差异(表4)。1龄和2龄若虫期存活率低于预蛹期和蛹期。1龄若虫在油菜和辣椒上存活率最高,达94.4%,而在南瓜上存活率最低,只有74.6%。除了辣椒以外,2龄若虫在其他寄主植物上的存活率均达到了90%—95%。预蛹期和蛹期存活率都在90%以上,最高达100%。可见,不同寄主植物对美洲棘蓟马预蛹和蛹的存活率影响不大。整体来看,未成熟期存活率在辣椒上最高,达80.1%,在黄豆上存活率最低,仅为64.3%。

表4 美洲棘蓟马在不同蔬菜上的存活率Table 4 The survival rate of Echinothrips americanus on different vegetables
3 讨论
3.1 美洲棘蓟马成虫对不同蔬菜寄主的偏好性
通过美洲棘蓟马成虫在不同的寄主植物的分布和产卵具有明显选择性。美洲棘蓟马成虫寄主偏好选择分为3组。第1组按虫量从大到小排列为南瓜、油菜、黄瓜、辣椒和黄豆,其成虫分布量均在10头/株以上。第2组为豆角、小白菜、胡萝卜、番茄和茄子,其成虫分布均在2头/株以下。第3组为茴香、紫苜蓿和圆白菜,其成虫分布量均为0。美洲棘蓟马产卵选择也分为3组。第1组按虫量从大到小排列为油菜、黄瓜、辣椒、南瓜和黄豆,其产卵量均在38粒卵/株以上。第2组为豆角、茄子、小白菜、番茄和胡萝卜,其产卵量均在15粒卵/株以下。第3组为茴香、紫苜蓿和圆白菜,产卵量均为0。可见,根据成虫寄主偏好性和产卵分布划分的寄主完全一致。这与裴昌莹等和袁成明等报导的西花蓟马成虫分布量和产卵量相一致的结论相符合[10-11]。然而美洲棘蓟马成虫寄主偏好和产卵选择的寄主植物的排序并不完全一致,油菜产卵量最大而南瓜上的成虫分布量最大。原因可能是南瓜的叶面积较大造成。王荣州等报导叶面积可以造成美洲斑潜蝇寄主选择上的差异[17]。余道坚等报导较小的叶面积会造成更多的竞争,不利于美洲斑潜蝇的取食和产卵[18]。本试验中茴香和紫苜蓿的成虫分布量和产卵量都为0,原因之一可能是其叶表面积太小。
国内已有研究报导造成美洲斑潜蝇和南美斑潜蝇成虫寄主选择差异的原因是寄主植物叶片的物理性状和化学成分[17-19]。物理性状方面除了叶表面积外,还有叶的表皮毛及叶表皮的硬度和厚度。在研究美洲斑潜蝇对黄瓜品种的寄主选择试验中发现,不同品种黄瓜的叶表皮毛的密度会对美洲斑潜蝇取食和产卵造成影响[20]。美洲棘蓟马成虫的寄主选择行为也可能受这些因素的影响。本研究中黄瓜上美洲棘蓟马的成虫分布量和产卵量都很高,可能是因为黄瓜较其他寄主叶表皮毛较密,使得昆虫一旦取食就难以脱离。而圆白菜上成虫分布量和产卵量都为0,可能是圆白菜叶表有厚厚的蜡质层,不易被锉吸式口器的昆虫取食造成[21]。叶片的营养物质和次生物质可能会影响美洲棘蓟马成虫寄主选择[19,22-23]。植物叶片中的次生物质影响植食性昆虫对寄主取食和产卵的选择[24]。不能适应柏树叶片次生物的柏小爪螨就不能取食相应的寄主[25]。有研究发现由于番茄中含有特异的抑制西花蓟马产卵物质,当其他西花蓟马嗜好寄主(例如西葫芦等)同时存在时会造成西花蓟马在番茄叶片上基本不产卵的现象。当撤去其他寄主后,西花蓟马又重新在番茄叶片上产卵[10]。美洲棘蓟马成虫在番茄上成虫分布量和产卵量也很低。这可能是由于其他嗜好的供试寄主存在,并且美洲棘蓟马成虫对番茄上次生物质较敏感所造成。
3.2 美洲棘蓟马在不同蔬菜寄主上的发育历期和存活率
不同的寄主植物上美洲棘蓟马未成熟各个时期的发育情况有所不同。不同的寄主主要影响其预蛹期以前的虫态,在这个发育期间存活率从74.6%到95.2%不等。预蛹期和蛹期的发育历期差异较小,平均存活率均接近100%。有研究表明西花蓟马在预蛹期和蛹期阶段的高存活率是其在此期间不食不动所造成的[11]。而美洲棘蓟马在预蛹期和蛹期期间同样不食不动,可能是其高存活率的主要原因。
美洲棘蓟马在不同寄主植物上的发育历期和存活率并不与其成虫寄主偏好和产卵选择完全一致。美洲棘蓟马在黄瓜上发育最快,但在油菜上产卵量最大。具体原因有待进一步研究。然而美洲棘蓟马在油菜上发育速率仅次于黄瓜排第二,但在油菜上的存活率最高。试验结果基本符合最适觅食理论[26]。有研究表明造成棉大卷叶螟在不同寄主植物上的发育历期和存活率差异的原因主要是寄主植物叶片的营养物质[27];造成针叶小爪螨在不同寄主植物上的生长发育状况差异的原因主要是寄主植物叶片的次生物质[28]。造成美洲棘蓟马在不同寄主植物上的发育历期和存活率差异的原因可能是寄主植物叶片的营养物质和次生物质。Mattson、McNeill和Southwood发现寄主植物可溶性蛋白质与可溶性糖的比例高,昆虫的生长发育速度和存活率就高[29-30]。
3.3 小结
该研究首次系统比较分析了美洲棘蓟马对蔬菜类寄主寄主的偏好性,结果表明该虫对黄瓜、油菜、辣椒等多种蔬菜类寄主植物具有较高的偏好性,且在这些寄主植物上具有很高的存活率,并能迅速完成其生长发育。因此,有必要加强该蓟马在蔬菜上的监测预警。美洲棘蓟马室内寄主选择试验由于缺少天敌及竞争的胁迫,必然造成其与自然界的情况有所差异,今后需要进一步对田间的发生情况进行调查研究,以更加全面的了解该虫的寄主范围与寄主偏好性。
致谢:浙江大学唐启义教授对本研究数据分析给予指导,卢先敏和金桂华在供试植物的种植等试验过程中给予帮助,特此致谢。
[1] Oetting R D.Echinothrips americanus,a thrips of increased significance on ornamentals//Proceedings of the 32nd Southern Nurserymen Association Research.Conference,Atlanta,Georgia,USA.1987:194-195.
[2] Mound R,Pollini A.Echinothrips americanus,a new pest of the Italian greenhouse.Informatore Fitopatologico,1999,49(6):61-64.
[3] Itoh K,Oguri A,Suzuki A.Occurrence and control of insect pests in Perilla on Aichi Prefecture.Proceedings of the Kansai Plant Protection Society,2003,45:71-72.
[4] Karadjova O and Krumov V.Echinothrips americanus Morgan(Thysanoptera:Thripidae),a new pest of the Bulgarian greenhouses//Proceedings of the International Scientific Conference at the 50th Anniversary of the University of Forestry.Plant Protection Section,Sofia,Bulgaria,2003:122-125.
[5] Mirab-balou M A,Lu H,Chen X X.First record of Echinothrips americanus Morgan(Thysanoptera:Thripidae)in mainland China,with notes on distribution and host plants.Acta Zootaxonomia Sinica,2010,35(3):674-679.
[6] Wei S J,Mirab-balou M,Shi B C,Liu J,Kang Z J,Chen X X,Lu H.External morphology and molecular identification of the newly found invasive pest Echinothrips americanus Morgan(Thysanoptera:Thripidae)in China.Acta Entomologica Sinica,2010,53(6):715-720.
[7] Zhang X C.New Record of Echinothrips americanus Morgan(Thysanoptera:Thripidae)in China Mainland and Its Biology[D].Yangling:North A &F University,2010:10-11.
[8] Brodbeck B V,Funderburk J,Stavisky J,Andersen P C,Hulshof J.Recent advances in the nutritional ecology of Thysanoptera,or the lack thereof//Marullo R,Mound L A,eds.Thrips and Tospoviruses:Proceedings of the 7th International Symposium on Thysanoptera.Canberra:Australian National Insect Collection,2002:145-153.
[9] Li J T,Deng J H,Duan,D X,Li Z Y,Ding Y M,Liu Z S,Xiao C.Distribution of Frankliniella occidentalis on different varieties of Dianthus caryophyllus in the field.Chinese Bulletin of Entomology,2009,46(2):276-279.
[10] Pei C Y,Zheng C Y.Selectivity of Frankliniella occidentalis(Pergande)on different host vegetables.Chinese Journal of Eco-Agriculture,2011,19(2):383-387.
[11] Yuan C M,Zhi J R,Cao Y,Ma H.Selectivity of Frankliniella occidentalis to vegetable hosts.Acta Ecologica Sinica,2011,31(6):1720-1726.
[12] Milne M,Walter G H.Host species and plant part specificity of the polyphagous onion thrips,Thrips tabaci Lindeman(Thysanoptera:Thripidae),in an Australian cotton-growing area.Australian Journal of Entomology,1998,37(2):115-119.
[13] Varga L,Fedor P J,Suvák M,Kisel'ák J,Atakan E.Larval and adult food preferences of the poinsettia thrips Echinothrips americanus Morgan,1913(Thysanoptera:Thripidae).Journal of Pest Science,2010,83(3):319-327.
[14] Ramakers P M J,van den Meiracker R A F,Mulder S.Predatory thrips as thrips predators.Mededelingen Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen,Universiteit Gent,2000,65(2):343-350.
[15] Tang Q Y.DPS Data Processing System-Experimental Design,Statistical Analysis and Data Mining.2nd ed.Beijing:Science Press,2010:75-209.
[16] Rasmann S,Turlings T C J.Simultaneous feeding by aboveground and belowground herbivores attenuates plant-mediated attraction of their respective natural enemies.Ecology Letters,2007,10(10):926-936.
[17] Wang R Z,Bao Z X.Preliminary report on host-selection of Liriomyza sativae Blanchard.Journal of Zhejiang Agricultural Sciences,2001,(3):149-152.
[18] Yu D J,Zhang R J,Zhou C Q,Li Y T.Host plant preference of the leafminer,Liriomyza sativae(Diptera:Agromyzidae).Acta Scientiarum Naturalium Universitatis Sunyateni,2000,39(1):86-90.
[19] Han J L,Pang B P,Pang Z,Gao S J,Cui W.Host plant selectivity and mechanism of Liriomyza huidobrensis on different varieties of eggplants.Journal of Inner Mongolia Agricultural University:Natural Science Edition,2005,26(3):29-32.
[20] Li S Q,Deng W X,Zhang Q D,Cui Q.Studies on the host selection of Liriomyza sativae Blanchard to Cucumber L.varieties.Journal of Huazhong Agricultural University:Natural Science Edition,2006,25(2):134-137.
[21] Zhou F C,Huang Z,Wang Y,Li C M,Zhu S D.Host plant selection of Bemisia tabaci(Gennadius)(Hemiptera:Aleyrodidae).Acta Ecologica Sinica,2008,28(8):3825-3831.
[22] He Y X,Yang X J,Weng Q Y,Wang M M.Host selectivity of the leaf miner,Liriomyza huidobrenisis to vegetable varieties.Fujian Journal of Agricultural Sciences,2006,21(1):28-31.
[23] Yuan H X,Zhang J P,Li Q.Host preference and mechanism of Tetranychus turkestani(Vgorag& Nikolski)to different cotton varieties.Xinjiang Agricultural Sciences,2009,46(6):1258-1262.
[24] Fagoonee I,Toory V.Preliminary investigations of host selection mechanisms by the leafminer Liriomyza trifolii.International Journal of Tropical Insect Science,1983,4(4):337-341.
[25] Yu J G,Han G H,Pei Y H,Liu X H,Meng Q Y,Sun X G.Effect of different cypresses on the host selection and development of Oligonychus perditus(Acarina:Tetranychidae).Forest Pest and Disease,2006,25(6):17-20.
[26] Scheirs J,Zoebisch T G,Schuster D J,de Bruyn L.Optimal foraging shapes host preference of a polyphagous leafminer.Ecological Entomology,2004,29(3):375-379.
[27] Wu J H,Huang Z,Ren S X,Zhou H P.Effect of host plant on the biological characters of Sylepta derogata(Lepidoptera:Pyralidae).Acta Phytophylacica Sinica,2007,34(6):659-660.
[28] Liu X H,Meng Q Y,Li Z X,Liu X F,Xun X G.Difference of bionomics of Oligonychus ununguis in four different host plants.Forest Pest and Disease,2007,26(1):15-17.
[29] Mattson W J.Herbivory in relation to plant nitrogen content.Annual Review of Ecology and Systematics,1980,11(1):391-403.
[30] McNeill S,Southwood T R E.Role of nitrogen in the development of insect-plant relationships//Harboune J B,ed.Biochemical Aspects of Plant and Animal Coevolution.New York:Academic Press,1978:77-98.
参考文献:
[6] 魏书军,马吉德,石宝才,宫亚军,刘静,康总江,陈学新,路虹.我国新入侵外来害虫美洲棘蓟马的外部形态和分子鉴定.昆虫学报,2010,53(6):715-720.
[7] 张晓晨.中国大陆新记录种美棘蓟马Echinothrips americanus Morgan(Thysanoptera:Thripidae)及其生物学研究[D].杨凌:西北农林科技大学,2010:10-11.
[9] 李江涛,邓建华,段登晓,李正跃,丁元明,刘忠善,肖春.西花蓟马在康乃馨不同品种上的田间分布.昆虫知识,2009,46(2):276-279.
[10] 裴昌莹,郑长英.西花蓟马对不同蔬菜寄主的选择性研究.中国生态农业学报,2011,19(2):383-387.
[11] 袁成明,郅军锐,曹宇,马恒.西花蓟马对蔬菜寄主的选择性.生态学报,2011,31(6):1720-1726.
[15] 唐启义.DPS数据处理系统-实验设计、统计分析及数据挖掘 (第二版).北京:科学出版社,2010:75-209.
[17] 王荣洲,鲍祖胜.美洲斑潜蝇寄主选择性研究初报.浙江农业科学,2001,(3):149-152.
[18] 余道坚,张润杰,周昌清,李雨庭.美洲斑潜蝇寄主选择性的研究.中山大学学报:自然科学版,2000,39(1):86-90.
[19] 韩靖玲,庞保平,庞琢,高书晶,崔威.南美斑潜蝇对不同茄子品种的选择性及其机理的研究.内蒙古农业大学学报:自然科学版,2005,26(3):29-32.
[20] 李绍勤,邓望喜,张求东,崔琪.美洲斑潜蝇对黄瓜品种的寄主选择性研究.华中农业大学学报,2006,25(2):134-137.
[21] 周福才,黄振,王勇,李传明,祝树德.烟粉虱(Bemisia tabaci)的寄主选择性.生态学报,2008,28(8):3825-3831.
[22] 何玉仙,杨秀娟,翁启勇,王茂明.南美斑潜蝇对蔬菜寄主的选择性.福建农业学报,2006,21(1):28-31.
[23] 袁辉霞,张建萍,李庆.土耳其斯坦叶螨对棉花不同品种(系)的寄主选择性及机理初步研究.新疆农业科学,2009,46(6):1258-1262.
[25] 于金国,韩国华,裴元慧,刘学辉,孟庆英,孙绪艮.不同柏树对柏小爪螨寄主选择及生长发育的影响.中国森林病虫,2006,25(6):17-20.
[27] 吴建辉,黄振,任顺祥,周慧平.寄主植物对棉大卷叶螟生物学特性的影响.植物保护学报,2007,34(6):659-660.
[28] 刘学辉,孟庆英,李中新,刘学峰,孙绪艮.针叶小爪螨在四种寄主植物上的生物学特性差异.中国森林病虫,2007,26(1):15-17.
