水通道蛋白对动物适旱性作用的研究进展
2013-12-10于伟江张建萍
于伟江 张建萍
(塔里木大学生命科学学院/新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,新疆阿拉尔 843300)
水是生命之源。水分参与了生物从出生到衰老死亡过程中几乎所有的生命活动。对于干旱区的生物来说水更为珍贵,如何能高效地利用有限的水资源决定着其生命能否继续下去。水通道蛋白(Aquaporin,AQP)是一族细胞膜上高效转运水分子的特异孔道蛋白,其种类和表达与水的分泌、吸收及细胞内外水的平衡有非常密切的关系[1]。水通道蛋白构成水分转运的特异性通道,能增强细胞膜对水分的通透性。
1 水通道蛋白的发现及命名
1988年,Agre等在鉴定人红细胞Rh血型抗原物质时,偶然发现了一种分子质量(Mr)为28 kDa的整合膜蛋白。氨基酸测序发现该蛋白和眼晶状体主体内在蛋白(major intrinsic protein,MIP)具有高度同源性,因此有了暂时的名称通道膜整合蛋白CHIP(Channel-like integral membrane protein)。并根据其Mr命名该蛋白为:形成通道的28 kDa膜整合蛋白,简称 CHIP28[2]。1991 年,他们完成了CHIP28的cDNA分子克隆,阐明了其cDNA的分子组成[3]。在功能鉴定时,他们将体外反转录合成的CHIP28 cDNA注入非洲爪蟾卵母细胞中,在低渗溶液中观察卵母细胞的膨胀度,发现表达了该基因的卵母细胞体积迅速增加,并于5 min内破裂[3]。为进一步确定CHIP28的功能,研究人员又将其重构于人工合成的脂质体,通过获得的活化能及渗透系数的数据表明,该脂质体对水表现出极高的通透性,但不通透其它小分子[4]。基于CHIP28这些明确的水转运功能,有人推荐使用“水孔蛋白(aquaporin)”这一名称。1997年,该蛋白被基因组命名委员会正式命名为Aquaporin-1(AQP1)。Agre本人由于这项奠基性的工作被授予诺贝尔化学奖。随着研究的不断深入,不断有新的AQP被发现,迄今为止,在哺乳动物体内已经发现13种水通道蛋白(AQP0~AQP12),统称为 aquaporin(AQP)[5]。这 13 种 AQP在通透水的功能上有着相似之处,但由于表达部位不同,因而又各自发挥着特异的生理功能。
2 水通道蛋白的分子结构
哺乳动物水通道蛋白的分子大小在26 000 000~34 000 000范围之内,其氨基酸序列同源性为19% ~52%[6]。因AQP是一类保守的疏水小分子整合膜蛋白,各种亚型三维结构相似,常以AQP1的结构作为代表。人AQP1基因位于第10号染色体,褐家鼠AQP1基因位于第4号染色体。AQP1的单体是一条由269个氨基酸残基构成的肽链,该单肽链在细胞膜上往返形成6个跨膜的α螺旋,并在胞内外形成5个环(A~E loop)。A、C及E环位于胞外,B和D环及肽链的N端和C端都位于胞内。目前,被人们广为接受的AQP1单体分子的三维结构为“沙漏”模式[6]:肽链中的B环和E环具有高度保守的NPA(Asn-Pro-Ala)特征性序列,分别位于细胞内和细胞外的疏水B环和E环向脂质双分子层中打折,两个NPA序列在折叠中形成一个供水分子通过的孔道。插入突变NPA区域后AQP1的水通道功能就丧失了,说明NPA区域具有重要的功能意义[7]。几乎所有AQP分子的B环和E环上都存在高度保守的NPA特征性序列。然而也存在例外情况:在AQP11和AQP12中仅发现E环上具有NPA序列,另一个在 B环上的 NPA序列分别由 NPC(Asn-Pro-Cys)和 NPT(Asn-Pro-Thr)替代[8]。对AQP1晶体学数据分析表明,水孔蛋白由4个亚基组成四聚体(图1A),每个亚基都有6个跨膜α螺旋组成(图1B和C)。每个水孔蛋白亚基单独形成一个供水分子运动的中央孔,孔的直径稍大于水分子的直径,约0.28 nm,水孔长约2 nm。研究发现,水分子的跨膜转运是通过水孔蛋白亚基的中心通道实现的,不是通过四聚体的中央孔洞(4个亚基衔接处的中心缝隙)[9]。尽管现在还没有完全揭示为何AQP1在对水分子快速通过的同时能有效阻止质子的通过,表现出对水分子的特异通透性,但已有的数据表明,这种特异性与两个半跨膜区NPA序列有关。AQP1中央孔的孔径无法通过比水分子大的物质,而两个NPA中的Asn残基所带的正电荷也排斥了质子的通过,因此,AQP1是一个高度特异的亲水通道。
3 水通道蛋白的分布及生理功能
应用免疫电镜、免疫组织化学、反转录PCR、实时荧光定量PCR、原位杂交、免疫印迹等方法,发现水通道蛋白广泛分布于机体的组织细胞中,尤其在与液体分泌和吸收有关的上皮细胞及内皮细胞中表达较多。在哺乳动物体内AQP的13种亚型,即AQP0~AQP12(表1)。依据它们的基因结构和通透性,将它们划分为3组:传统水通道蛋白(包括AQP0~2、AQP4~6及 AQP10)、甘油水通道蛋白(包括 AQP3、AQP7及 AQP9)和未明确分类的AQP8、AQP11 及 AQP12[11]。
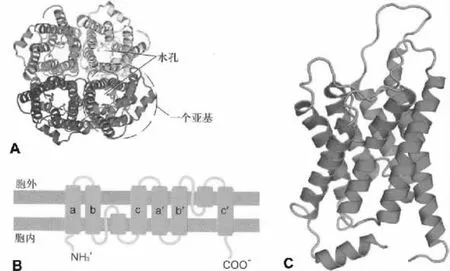
图1 水通道蛋白的分子结构[10]

表1 水通道蛋白成员的组织分布及功能
4 水通道蛋白与动物的适旱性
每种AQP都有其独特的组织分布和细胞表达位置,以满足机体对水分的需要。在肺的水代谢方面,Ma等[12]和 Bai等[13]发现与野生型小鼠相比,AQP1或AQP5基因敲除小鼠肺内水转运被抑制高达90%,AQP1和AQP5基因共同敲除小鼠肺内水转运进一步被抑制50%,这些结果说明AQP在肺泡水分转运中起重要作用。King等[14]研究证明AQP1是肺血管通透性的决定因素。Song等[15]证明AQP5基因敲除小鼠其气道粘膜下腺分泌液减少。Yasui等[16]发现在生产期间,大鼠肺 AQP1、AQP4及AQP5 mRNA表达量增加,提示这些AQP可能与清除肺水有关。
在肾水代谢方面,Ma等[17]发现与野生型小鼠相比,AQP1基因敲除小鼠表现出多饮和多尿的症状,若限制饮水则会引起机体严重脱水,同时尿浓缩能力也严重受损。Chou等[18]离体实验证明,AQP1缺失将导致肾脏近曲小管和亨勒袢降支细段通透性降低,重吸收水分的能力降低。AQP2是肾脏重吸收水分浓缩尿液从而调节机体水平衡的主要分子基础,AQP2只存在于集合管(CD)中,在其他部位未见表达,而CD是尿液分泌最后的决定性部位,这决定了AQP2对肾脏水调节独特的作用。Sasaki等[19]研究发现缺水口渴可使AQP2 mRNA在肾脏的表达量增加。Tsumura等[20]发现多饮水影响AQP2基因的转录而导致其表达下降。AQP3分布于肾皮质到肾乳头的整个CD系统中,AQP4则主要表达于主细胞的基底膜。AQP3和AQP4共同对由AQP2转运进入细胞内的水分起分散作用。Kim等[21]研究发现与野生型小鼠相比,AQP3基因敲除小鼠的饮水量和尿量增加12倍以上,出现严重尿崩症和尿浓缩障碍的症状。
在消化道的水代谢方面,每天在人类的消化道的液体转运量有8~10 L之多,而且至少有7种水通道在消化道上皮表达。Ma等[22]发现就盲肠粪便中水分含量,AQP4基因敲除小鼠和野生型小鼠之间没有差别,但排泻的粪便中的水分含量差异显著,AQP4基因敲除小鼠排泻的粪便中的水分含量明显升高。Wang等[23]在体结肠灌流实验证明,AQP4基因敲除小鼠,其结肠上皮水的通透性明显降低,证明AQP4参与结肠液体中水分的重吸收。这些研究表明,AQP的表达与肺泡水分转运、尿的浓缩、排泄粪便中水分含量的多少等都有紧密的关系。然而,AQP在野生动物适应干旱环境中的作用尚未见报道。
干旱区野生动物的生存环境极度干旱。自然选择是生物进化的驱动力。面对压力环境,生物会进化为不同环境下所特有的分类群或生存特征[24]。这种针对压力环境所产生的适应性反应可以导致加快生物谱系的进化或者特定压力敏感蛋白的进化。因此环境压力可以加速特定压力敏感蛋白的进化速率,产生针对特定环境的新功能或对原有功能的加强,可提高动物对这种压力环境的适合度[25]。正如Yang等[26]发现青藏高原的高原鼠兔(Ochotona curzoniae)在寒冷压力的作用下,其瘦素蛋白(leptin)发生了适应性的功能进化,产生了新的功能或加强了原有功能。笔者的前期研究工作发现,塔里木盆地的塔里木兔(Lepus yarcandensis)在干旱压力的作用下,其肾脏AQP1和AQP2的表达比家兔强,并且表达的部位也更加广泛,这有利于增强塔里木兔肾脏对水的重吸收能力,以保存体水。该研究结果表明,塔里木兔肾脏水通道蛋白对其适应干旱缺水的环境,具有重要作用。综上所述,鉴于干旱区野生动物干旱缺水的生存环境及AQP在动物水代谢调节中的关键作用,推测在干旱区野生动物中,AQP可能作为一种干旱敏感蛋白,在动物对干旱环境的生态适应过程中起到了重要的作用。
致谢:中国科学院动物研究所王德华研究员对本文的撰写提供了有益帮助,在此表示衷心的感谢。
[1]才丽平,赵金茹,林庶茹,等.水通道蛋白研究进展[J].解剖科学进展,2003,9(2):167 -170.
[2]Denker B M,Smith B L,Kuhajda F P,et al.Identification,purification,and partial characterization of a novel Mr28,000 integral membrane protein from erythrocytes and renal tubules[J].J.Biol.Chem,1988,(263):15634-16542.
[3]Preston G M,Agre P.Isolation of the cDNA for erythrocyte integral membrane protein of 28 kilodaltons:member of an ancient channel family[J].Proc.Natl.Acad.Sci.USA,1991,(88):11110 -11114.
[4]李敏,陈少军,陈学群,等.脑水肿的AQP4调节机制研究进展[J].浙江大学学报(医学版),2013,(1):60-64.
[5]Van Os CH,Deen P M T,Dempster J A.Aquaporins:Water selsction channels in biological membranes.Molecular structure and tissue distribution[J].Bipchim Biophy Acta,1994,(1197):291 -309.
[6]Jung J S,Preston G M,Smith B L,et al.Molecular structure of the water channel through aquaporin CHIP,the hourglass model.J Biol Chem,1994,269:14648—14654.
[7]Preston G M,Jung J S,Guggino W B,Agre P.Membrane topology of aquaporin CHIP28.Analysis of functional epitope - scanning mutants by vectorial proteolysis[J].Biol Chem,1994,269:1668 -1673.
[8]Morishita Y,Sakibe Y,Sasaki S,et al.Molecular mechanisms and drug development in aquaporin water channel diseases:aquaporin superfamily(superaquaporins):expansion of aquaporins restricted to multicellular organisms.J Pharmacol Sci,2004,96:276 -279.
[9]Murata K,Mitsuoka K,Hirai T,et al.Structural determinants of water permeation through aquaporin -1.Nature,2010,407:599 -605.
[10]翟中和,王喜忠,丁明孝,等.细胞生物学[M].北京:高等教育出版社,2011.71 -73.
[11]王晶,桑建利.水通道蛋白的基本结构与特异性通透机理[J].生物学通报,2011,46(2):19-22.
[12]Ma T,Fukuda N,Song Y.Lung fluid transport in aquaporin -5 knockout mice[J].J Clin Invest,2000 ,105:93– 100.
[13]Bai C,Fukuda N,Song Y,et al.Lung fluid transport in aquaporin-1 and aquaporin-4 knockout mice[J].J Clin Invest,1999,103:555 – 561.
[14]King LS,Nielsen S ,Agre P,et al.Decreased pulmonary vascular permeability in aquaporin1 null human [J].Proc Natl Acad Sci USA,2012,99(2):1059 -1063.
[15]Song Y,Verkman AS.Aquaporin -5 dependent fluid secretion in airway submucosal glands[J].J Biol Chem,2011,276(44):41288 – 41292.
[16]Ya S M,Serlachius E,Lofgren M,et al.Perinatal changes in expression of aquaporin-4 and other water and ion transporters in rat lung[J].J Physiol,1997,505(pt 1):3-11.
[17]Ma T,Yang B,Gillespie A,et al.Severly impaired urinary concentrating ability in transgenic mice lacking aquaporin1 water channels[J].J Biol Chem,1998,273:4296– 4299.
[18]Chou C L,Knepper M A,Van Hoek A N,et al.Reduced waterpermeability and altered ultrastructure in thin dascending limb of henle in aquaprinl null mice[J].J Clin Ivest,1999,103(4):491 -496.
[19]Sasaki S,Fushimi K,Ishibashi K,et al.Water channels in the kidney collecting duct[J].Kidney Int,1995,48(4):1082-1087.
[20]Tsumura K,Li X,Murdiastuti K,et al.Downregulation of AQP2 expression in the kidney of polydipsic STR/N mice[J].Am J Physiol Renal Physiol,2006,290(2):F478-F485.
[21]Kim S W,Gresz V,Rojek A,et al.Decreased expression of AQP2 and AQP4 water channels and Na,K -ATPase in kidney collecting duct in AQP3 null mice [J].Biol Cell,2005,97(10):765 -778.
[22]Ma T,Jayaraman S,Wang K S,et al.Defective dietary fat processing in transgenic mice lacking aquaporin1 water channels[J].Am J Physiol Cell Physiol,2011,280(1):C126–1341.
[23]Wang K S,Ma T,Filiz F,et al.Colon water transport in transgenic mice lacking aquaporin4 water channels[J].Am J Physiol Gast rointest Liver Physiol,2010,279(2):G463– 4701.
[24]Huntley B,Webb T.Migration:Species’response to climatic variation caused by changes in the earth's orbit[J].J Biogeogr,1989,16:5 -19.
[25]Nei M.Selectionism and neutralism in molecular evolution [J].Mol BioI Evol,2005,22:2318 -2342.
[26]Yang J,Wang Z L,Zhao X Q,et al.Natural selection and adaptive evolution of leptin in the Ochotona family driven by the cold environmental stress[J].PLoS ONE,2008.3:e1472.
