不对称还原大位阻二芳香基甲酮的羰基还原酶的基因克隆及性质分析
2013-11-12李哲刘卫东陈曦贾士儒吴洽庆朱敦明马延和
李哲,刘卫东,陈曦,贾士儒,吴洽庆,朱敦明,马延和
1 天津科技大学生物工程学院,天津 300457
2 中国科学院天津工业生物技术研究所 工业酶国家工程实验室,天津 300308
光学纯二芳香基甲醇是合成许多手性药物的重要中间体,例如:(R)-nebenodine,(R)-orpheadrine和 (S)-carbinoxamine[1-2]。使用芳香基亲核试剂对芳基醛进行加成是合成二芳香基甲醇常用的方法[1],但是芳香基亲核试剂有许多缺点,如价格昂贵、不适合大规模应用或产物的 ee值和收率较低等[3]。对前手性二芳香基甲酮进行不对称还原是获得二芳香基甲醇最直接的方法。用化学合成的方法可以达到这种目的,但只对一些具有特殊结构的底物,如苯环的邻位有取代基或者羰基的一边是芳香杂环时,产物的ee值才较高[4-5],而对于羰基两侧的取代基差异性较小,如对位甲基取代的二苯基甲酮,则产物的光学纯度非常低(ee<26%)。
羰基还原酶具有高度的化学、区域和立体选择性,常用于手性醇的合成,一些酶催化的反应已经被用于工业生产[6-8]。虽然利用分离纯化的羰基还原酶催化二芳香基酮不对称还原已有报道[3,9],但因为该类底物的位阻大、羰基两侧的取代基差异性较小,常常使得反应的活性和立体选择性不能满足医药工业的需求。因此,对二芳香基甲酮的不对称还原仍是生物催化中很有挑战性的难题之一,寻找具有较高反应活性和立体选择性的还原较大位阻二芳香基甲酮的羰基还原酶具有很好的理论和应用价值。
丰富多样的微生物为发现新的羰基还原酶提供了宝贵的资源,然而传统的菌种筛选方法耗时、低效。近年来,基因组测序技术的发展使得可以通过基因数据分析快速发现新的目标酶[10-11]。毕赤酵母 GS115基因组序列已被测定[12],本文通过对毕赤酵母GS115的基因组数据进行分析,发现一个新的羰基还原酶,能够不对称还原二芳香基甲酮类化合物,并对其酶学性质进行了研究,为该酶的进一步应用奠定了基础。
1 材料与方法
1.1 菌株与质粒
毕赤酵母Pichia pastoris GS115为实验室保存,大肠杆菌Escherichia coli Rosseta2 (DE3)、DH5α、表达质粒 pET32a (+)均购自 Novagen公司。
1.2 培养基、主要试剂及仪器
P. pastoris GS115采用YPD培养基在30 ℃进行培养。大肠杆菌DH5α和Rosseta2 (DE3)在37 ℃用Luria-Bertani培养基进行培养。
限制性内切酶、T4 DNA连接酶、DNA和蛋白Marker购于Fermentas公司;高保真PCR扩增试剂盒购自大连宝生物工程有限公司;DNA凝胶回收试剂盒购于北京赛百盛生物技术公司。二芳香基甲酮类化合物购于Sigma公司;二芳香基醇消旋体通过NaBH4还原制得;蛋白纯化仪:ÄKTA purifier 10,所用层析柱均为GE公司,高效液相色谱仪为 Agilent 1206型;手性分离柱OD-H及AD-H柱均购于DAICEL公司。其他所用试剂均为分析纯。
1.3 目的基因的克隆表达
P. pastoris GS115中目的基因pascr序列从EMBL数据库获得 (GenBank Accession No.XM_002492630.1)。根据所选择的蛋白对应的核酸序列设计引物,正向:5¢-CGCCATATG GTTTC TAAGGTTTTATTGACA-3¢,反向:5¢-CCGCTC GAGTTTATTAGCACGCAATATCT-3¢,下划线部分分别为NdeⅠ和XhoⅠ的酶切位点。PCR扩增条件:94 ℃预变性 5 min;94 ℃变性 30 s,55 ℃退火30 s,72 ℃延伸50 s,30个循环;72 ℃延伸15 min。PCR产物通过琼脂糖凝胶电泳分离纯化,核酸回收试剂盒回收目的片段,回收产物用NdeⅠ和 XhoⅠ进行双酶切,酶切产物再经琼脂糖凝胶电泳回收后用T4 DNA连接酶连接到用相同内切酶切过的质粒pET32a (+),连接产物转化到大肠杆菌DH5α感受态中,挑取阳性克隆进行测序验证。将测序正确的重组质粒转化Rosseta2(DE3)感受态细胞。
将成功转化的单克隆接种于 10 mL含有终浓度100 μg/mL氨苄青霉素以及34 μg/mL氯霉素的LB液体培养基中,37 ℃、200 r/min培养过夜,然后转接到含有同样抗生素的800 mL的LB培养基中,培养到OD600为1.0左右,向培养基中添加终浓度为0.1 mmol/L异丙基-β-D硫代半乳糖苷 (Isopropyl-β-D-thiogalactopyranoside),37 ℃诱导4 h,于4 ℃、7 000×g离心15 min收集菌体。
1.4 目的蛋白的分离纯化
收集菌体用生理盐水洗涤 2次后重悬于100 mL预冷的缓冲液A中 (20 mmol/L Tris-HCl,pH 8.0,5%甘油,500 mmol/L NaCl,20 mmol/L咪唑),高压匀浆破碎,20 000×g离心30 min,弃去细胞碎片。上清用0.22 μm滤膜过滤后上样,上样流速为 3 mL/min,用 B液 (20 mmol/L Tris-HCl,pH 8.0,5%甘油,500 mmol/L NaCl,500 mmol/L咪唑)进行0~250 mmol/L咪唑线性洗脱,收集不同组分。用SDS-PAGE以及活力测定检验目的蛋白所在组分,合并纯度较高的样,使用超滤管 (Millipore截留分子量为10 kDa)进行超滤浓缩和脱盐。蛋白质浓度用BCA试剂盒进行测定,使用牛血清白蛋白BSA作为标准蛋白。
1.5 酶的辅酶偏好性及分子量测定
羰基还原酶 PasCR的辅酶偏好性是以苯乙酮为底物使用96孔板在酶标仪上进行活力测定,反应总体积为200 μL,体系中含有酶液、底物以及辅酶NADPH或NADH,通过连续监测340 nm吸光度下辅酶的消耗 (ε=6 220 L/(mol·cm))进行测定。每分钟消耗1 μmol辅酶所需要的酶量定义为一个酶活单位。动力学参数在最适pH下按上述方法分别以苯乙酮和 4-氯-3-羰基丁酸乙酯为底物测定。
羰基还原酶 PasCR在溶液中的聚集状态通过结合凝胶层析以及SDS-PAGE来进行确定。所用层析柱为Superdex 200 10/300 GL。流动相为20 mmol/L磷酸缓冲液,150 mmol/L NaCl,pH 7.2,流速为0.4 mL/min。蛋白的分子量大小通过和标准蛋白的保留体积进行计算得到。标准蛋白为:Ovalbumin (43.0 kDa),Conalbumin (75.0 kDa),Conalbumin (158.0 kDa),Ferritin (440.0 kDa),Thyroglobulin (669.0 kDa)。
1.6 pH及温度对酶活的影响
PasCR的最适反应pH通过在不同pH的缓冲液中测定酶的活力确定。缓冲液分别为100 mmol/L的醋酸/醋酸钠缓冲液 (pH 4.0~6.0),100 mmol/L的磷酸钠缓冲液 (pH 6.0~8.0),100 mmol/L Tris-HCl缓冲液 (pH 8.0~9.0)。PasCR的最适反应温度通过以苯乙酮为底物在不同温度下测定酶的活力确定。
pH稳定性是将酶在不同pH的缓冲液中4 ℃孵育30 h,然后测定剩余活性,不同pH缓冲液如上所述。酶热稳定性是将纯化后的酶放在含有100 mmol/L NaCl的Tris-HCl (20 mmol/L,pH 8.0)缓冲液中不同温度下分别孵育不同时间,然后检测残余酶活力。
1.7 不同化学物质对酶稳定性的影响
在等量的酶液中分别加入金属螯合剂EDTA、变性剂等不同化学物质,室温下孵育0.5 h后,测定其残留活性,以不处理的酶液的活性作为对照。
1.8 酶对二芳香基甲酮类化合物的不对称还原
葡萄糖脱氢酶偶联的辅酶再生体系用于催化反应。1 mL磷酸钠缓冲液 (100 mmol/L,pH 6.5)的反应体系中包含:底物100 μL (母液100 mmol/L溶于甲醇),葡萄糖18 mg,NADP+0.5 mg,葡萄糖脱氢酶2 mg,PasCR 2 mg,30 ℃、200 r/min摇床反应。过夜反应后,用相同体积的甲基叔丁基醚萃取,取出有机相层,除去溶剂后用HPLC流动相溶解,HPLC检测ee值及转化率,产物的构型通过和文献比较确定[9]。
2 结果与分析
2.1 羰基还原酶PasCR序列比对及分析
基因pascr位于毕赤酵母GS115的3号染色体上,编号为 chr3_0449,其所编码蛋白的标注为“putative dihydrokaempferol 4-reductase”。Blast结果显示 PasCR与短链脱氢酶家族的蛋白有较高的同源性。PasCR与小家鼠 Mus musculus delta-5-3-β-hydroxysteroid 脱氢酶 (3β-HSD Gene ID M58567)[13]、木兰假丝酵母 Candida macedoniensis的羰基还原酶 (MER Gene ID AB183149)[14]、赭色掷孢酵母 Sporobolomyces salmonicolor AKU4429的羰基还原酶 (SSCR Gene ID AF160799)[15]、鲁氏接合酵母Zygosaccharomyces rouxii的羰基还原酶(Zygosaccharomyces gene ID AF178079)[16]、酿酒酵母 Saccharomyces cerevisiae的羰基还原酶(YOL151w Gene ID Z74893)[17]分别有 25%、38%、31%、31%、43%的同源性,序列比对的结果如图1所示。短链脱氢酶家族的蛋白主要有3个功能区:辅酶结合区、反应催化区及底物结合区。从比对结果可以看出PasCR的N端非常保守,主要是结合辅酶,三联催化基团 (Tyr174、Ser135、Lys178)这在短链脱氢酶家族中也是非常保守的;而C端同源性较低,主要决定了酶的底物谱及立体选择性[18]。因此,与这些已知的短链脱氢酶家族中的酶相比,PasCR可能具有比较独特的底物谱及立体选择性。
2.2 羰基还原酶PasCR的构建表达及纯化
以毕赤酵母GS115基因组为模板,通过PCR扩增的基因片段 (1 065 bp),经测序验证和目的基因一致。成功构建出工程菌pET32a(+)-pascr,并在大肠杆菌 Rosseta2 (DE3)中进行了重组表达,蛋白的C端带有6×His tag标签。经Ni-NTA纯化,得到了高纯度的目的蛋白。蛋白表达及纯化SDS-PAGE图谱见图2。PasCR单体的分子量为 39.2 kDa,凝胶层析结果显示酶的分子量大约为80 kDa,说明其在溶液中以二聚体形式存在。
2.3 酶动力学参数测定

图1 羰基还原酶PasCR序列比对结果Fig. 1 Amino acid sequence alignment of PasCR from P. pastoris GS115 with Mus musculus delta-5-3-β-hydroxysteroid dehydrogenase (3β-HSD), Candida macedoniensis carbonyl reductase (MER), Sporobolomyces salmonicolor AKU4429 carbonyl reductase (SSCR), Zygosaccharomyces rouxii carbonyl reductase(Zygosaccharomyces )and Saccharomyces cerevisiae carbonyl reductase (YOL151w).

图2 PasCR蛋白纯化SDS-PAGE图谱Fig. 2 SDS-PAGE analysis of the purified PasCR.1: molecular mass standard; 2: uninduced; 3: crude cell-free extract; 4: purified PasCR.
PasCR专一性地利用NADPH作为辅酶,在以NADH作为辅酶时对苯乙酮没有表现出活力。PasCR在最适pH条件下对苯乙酮、4-氯-3-羰基丁酸乙酯和辅酶NADPH的动力学参数列于表1。从表中可以看出羰基还原酶 PasCR对脂肪族的4-氯-3-羰基丁酸乙酯的亲和力和催化效率都远高于芳香族的苯乙酮,因此 PasCR对 4-氯-3-羰基丁酸乙酯具有更高的催化活性。
2.4 酶的最适反应pH、温度及pH和热稳定性
以苯乙酮为底物,羰基还原酶 PasCR在pH 6.5的磷酸缓冲液中表现出了最高活性,结果如图 3所示。在 pH 6.5的条件下羰基还原酶PasCR在35 ℃条件下表现出了最高活性,而到60 ℃时没有任何活性 (图 4),可能在该温度下PasCR已失活。

表1 PasCR对苯乙酮、4-氯-3-羰基丁酸乙酯和辅酶NADPH的动力学参数Table 1 Kinetic parameters of PasCR for acetophenone, ethyl 4-chloro-3-oxobutanoate and NADPH

图3 酶的最适反应pHFig. 3 Effect of pH on enzyme activity in different buffers, acetic acid/sodium acetate buffer (●), sodium phosphate buffer (■)and Tris-HCl buffer (▲). The maximum activity in pH 6.5 was taken as 100%.

图4 酶的最适反应温度Fig. 4 Effect of temperature on enzyme activity. The optimal temperature of the carbonyl reductase PasCR was estimated at various temperatures (25 °C − 60 °C)in phosphate buffer (100 mmol/L, pH 6.5).
pH稳定性研究表明,在pH 4.5~9.0范围内PasCR有较好的稳定性 (图5)。热稳定研究表明,在30 ℃的条件下PasCR有很好的稳定性,而在45 ℃和 50 ℃条件下 PasCR的稳定性较差,在45 ℃时PasCR的半衰期约80 min,而在50 ℃的条件下PasCR的半衰期只有10 min (图6)。

图5 酶的pH稳定性Fig. 5 Effect of pH on enzyme stability in different pH buffers, acetic acid/sodium acetate buffer (pH 4.5−6.0,●), sodium phosphate buffer (pH 6.0−8.0, ■)and Tris-HCl buffer (pH 8.0−9.0, ▲).
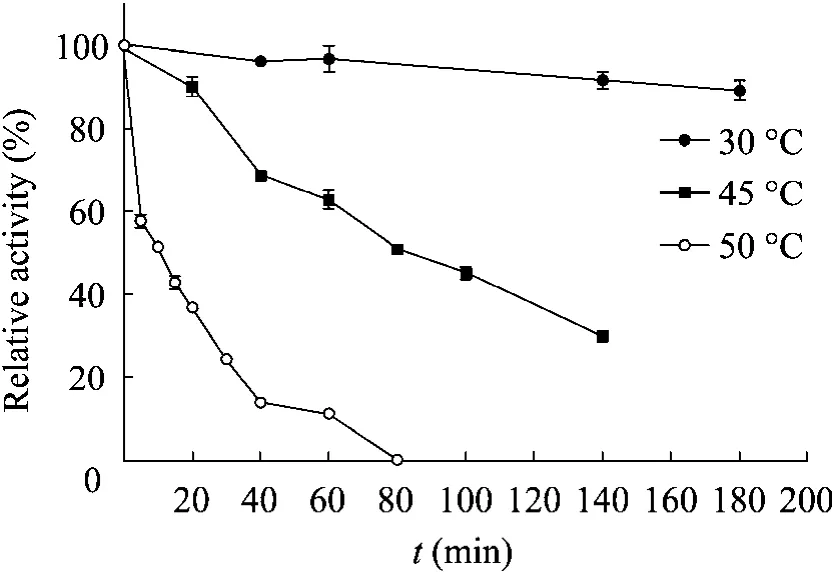
图6 酶的温度稳定性Fig. 6 Effect of temperature on enzyme stability, the samples was withdrawn at different time intervals, the activity of enzyme without pre-incubation was taken as 100%.
2.5 化学物质对酶的稳定性影响
不同化学物质对酶稳定性影响分析结果见表2,从表中可以看出金属螯合剂EDTA对酶的活力没有明显影响,说明该酶不需要金属离子的辅助。β-巯基乙醇主要是破坏二硫键,对酶的活性也没有明显影响。一些非离子型表面活性剂如曲拉通X−100当添加1%时PasCR酶活丧失了近一半,而SDS浓度为0.5%时直接使酶失活。四氢呋喃可以使酶的活性提高,但当浓度大于10%时酶的活性明显降低 (数据未显示)。

表2 不同化学物质对酶活的影响Table 2 Effect of different chemicals on PasCR activity
2.6 对二芳香基甲酮的不对称还原
PasCR对二芳香基甲酮类化合物表现出了较好的活性,对它们进行催化反应的结果如表3所示。当底物为4-氟苯甲酮时,仅有5%的转化率和2.7%的ee值,随着对位取代基团位阻的变大和供电子效应的增强,氯取代和甲基取代的二苯甲酮的转化率分别提高至40%和80%,ee值分别提高至 67%和 85%,并且产物构型均由 R变成了 S。而邻位取代基是甲基和氯时,产物的ee值分别为78%和80%,但是转化率都较低。当底物为分别为2-苯甲酰吡啶和3-苯甲酰吡啶时,与之前的文献[3,9,19-21]报道不同的是,产物的转化率达90%,但是ee值仅有22%和14%。当底物为1,2-二苯基乙酮时,产物的转化率为88%,ee值达到了92%。说明PasCR对二苯基取代的一系列大位阻底物表现出了较好的转化率和ee值。

表3 羰基还原酶PasCR对二芳香甲酮化合物的还原Table 3 Reduction of diary ketones by carbonyl reductase PasCR
3 结论
本文成功从毕赤酵母GS115中克隆出1个新的羰基还原酶基因,将其在大肠杆菌中进行表达纯化,并对其酶学性质进行了研究。结果表明,该酶以NADPH作为辅酶,能够对一些体积较大的二芳香基甲酮类物质进行不对称还原,当苯环上对位的取代基越大时产物的ee和转化率越高。最适反应pH为6.5,最适反应温度为35 ℃,在溶液中以二聚体形式存在。该酶在催化一些位阻较大的二芳香基甲酮类化合物,表现出与已经报道的羰基还原酶不同的特性。文献报道来源于赭色掷孢酵母Sporobolomyces salmonicolor的及一些商业来源的羰基还原酶对对位取代的二苯基甲酮表现出较低的立体选择性,而还原苯甲酰基吡啶时,产物的ee值可达99%[3,9]。固定化的酿酒酵母细胞不对称还原 2-苯甲酰基吡啶生成的醇ee值能达到有96%[22]。与此相反,PasCR不对称还原4-甲基二苯甲酮和4-氯二苯甲酮时,得到产物的ee较高,而对2-苯甲酰吡啶和3-苯甲酰吡啶表现出较低的立体选择性。由于对位取代的二苯基甲酮的不对称还原具有更大的挑战性,来源于毕赤酵母的羰基还原酶 PasCR将具有很好的应用价值和潜力。
[1]Schmidt F, Stemmler RT, Rudolph J, et al.Catalytic asymmetric approaches towards enantiomerically enriched diarylmethanols and diarylmethylamines. Chem Soc Rev, 2006, 35(5):454−470.
[2]Devalia J, De Vos C, Hanotte F, et al. A randomized, double-blind, crossover comparison among cetirizine, levocetirizine, and ucb 28557 on histamine-induced cutaneous responses in healthy adult volunteers. Allergy, 2001, 56(1): 50−57.
[3]Truppo MD, Pollard D, Devine P. Enzymecatalyzed enantioselective diaryl ketone reductions.Org Lett, 2007, 9(2): 335−338.
[4]Welch CJ, Grau B, Moore J, et al. Use of chiral HPLC-MS for rapid evaluation of the yeast-mediated enantioselective bioreduction of a diaryl ketone. J Org Chem, 2001, 66(20):6836−6837.
[5]Corey EJ, Helal CJ. Reduction of carbonyl compounds with chiral oxazaborolidine catalysts: A new paradigm for enantioselective catalysis and a powerful new synthetic method. Angew Chem Int Ed, 1998, 37(15): 1986−2012.
[6]Yang Y, Zhu DM, Piegat TJ, et al. Enzymatic ketone reduction: mapping the substrate profile of a short-chain alcohol dehydrogenase (YMR226c)from Saccharomyces cerevisiae. Tetrahedron:Asymmetry, 2007, 18(15): 1799−1803.
[7]Moore JC, Pollard DJ, Kosjek B, et al. Advances in the enzymatic reduction of ketones. Acc Chem Res,2007, 40(12): 1412−1419.
[8]Zhu DM, Wu QQ. Engineering the enantioselectivity of biocatalysts. Chin J Biotech,2009, 25(12): 1770−1778 (in Chinese).朱敦明, 吴洽庆. 生物催化剂立体选择性的基因工程改造. 生物工程学报, 2009, 25(12):1770−1778.
[9]Li H, Zhu DM, Hua L, et al. Enantioselective reduction of diaryl ketones catalyzed by a carbonyl reductase from sporobolomyces salmonicolor and its mutant enzymes. Adv Synth Catal, 2009, 351(4):583−588.
[10]Zhu DM, Mukherjee C, Biehl ER, et al. Discovery of a mandelonitrile hydrolase from Bradyrhizobium japonicum USDA110 by rational genome mining. J Biotech, 2007, 129(4): 645−650.
[11]Yu HM, Luo H, Shi Y, et al. Application of bioinformatics in researches of industrial biocatalysis. Chin J Biotech, 2004, 20(3): 325−330(in Chinese).于慧敏, 罗晖, 史悦, 等. 生物信息学在工业生物催化研究中的应用. 生物工程学报, 2004,20(3): 325−330.
[12]De Schutter K, Lin YC, Tiels P, et al. Genome sequence of the recombinant protein production host Pichia pastoris. Nat Biotech, 2009, 27(6):561−566.
[13]Bain P A, Yoo M, Clarke T, et al. Multiple forms of mouse 3 beta-hydroxysteroid dehydrogenase/delta 5-delta 4 isomerase and differential expression in gonads, adrenal glands, liver, and kidneys of both sexes. Proc Nat Acad Sci USA,1991, 88(20): 8870−8874.
[14]Kataoka M, Hoshino-Hasegawa A, Thiwthong R, et al. Gene cloning of an NADPH-dependent menadione reductase from Candida macedoniensis,and its application to chiral alcohol production.Enzyme Microb Technol, 2006, 38(7): 944−951.
[15]Kita K, Fukura T, Nakase KI, et al. Cloning,overexpression, and mutagenesis of the Sporobolomyces salmonicolor AKU4429 gene encoding a new aldehyde reductase, which catalyzes the stereoselective reduction of ethyl 4-chloro-3-oxobutanoate to ethyl(S)-4-chloro-3-hydroxybutanoate. Appl Environ Microbiol, 1999, 65(12): 5207−5211.
[16]Costello CA, Payson RA, Menke MA, et al.Purification, characterization, cDNA cloning and expression of a novel ketoreductase from Zygosaccharomyces rouxii. Eur J Biochem, 2000,267(17): 5493−5501.
[17]Choi YH, Choi HJ, Kim D, et al. Asymmetric synthesis of (S)-3-chloro-1-phenyl-1-propanol using Saccharomyces cerevisiae reductase with high enantioselectivity. Appl Microbiol Biotechnol,2010, 87(1): 185−193.
[18]Joernvall H, Persson B, Krook M, et al. Short-chain dehydrogenases/reductases (SDR). Biochemistry,1995, 34(18): 6003−6013.
[19]Takemoto M, Achiwa K. The synthesis of optically active pyridyl alcohols from the corresponding racemates by Catharanthus roseus cell cultures.Tetrahedron: Asymmet, 1995, 6(12): 2925−2928.
[20]Salvi NA, Chattopadhyay S. Studies on Rhizopus arrhizus mediated enantioselective reduction of arylalkanones. Tetrahedron, 2001, 57(14):2833−2839.
[21]Takemoto M, Tanaka K. Synthesis of optically active α-phenylpyridylmethanols by Camellia sinensis cell culture. J Mol Catal B: Enzym, 2001,15(4/6): 173−176.
[22]Takemoto M, Achiwa K. Synthesis of optically active α-phenylpyridylmethanols with Baker's yeast. Chem Pharm Bull, 1994, 42(4): 802−805.
