游动放线菌原生质体诱变选育阿卡波糖高产菌株*
2013-10-30王远山牛鑫淼郑裕国
王远山,牛鑫淼,郑裕国
1(浙江工业大学生物与环境工程学院,浙江 杭州,310014)
2(浙江工业大学生物工程研究所,浙江 杭州,310014)
阿卡波糖(acarbose)是一种低聚糖,由环己烯醇氨(valienamine),4-氨基-4,6-双脱氧葡萄糖和麦芽糖组成[1]。它还是一种α-葡萄糖苷酶抑制剂,能够有效抑制肠道内的某些消化酶,降低餐后的血糖水平,主要用于治疗Ⅱ型糖尿病,具有毒副作用小、作用温和持久等优点[2]。阿卡波糖最早由游动放线菌Actinoplanes sp. SE50、SE82 和SE18 的代谢产物中分离出来,并于1990 年由德国Bayer 公司生产上市,目前已经得到了广泛的应用。
目前,阿卡波糖的工业规模生产均通过游动放线菌发酵获得[3],菌株的生产能力很关键,因此很多研究工作都集中于菌株的选育和发酵优化。程新等[4-5]从土壤中筛选到1 株游动放线菌Actinoplanes sp. A56,经过培养基优化后阿卡波糖产量达到了1 043 mg/L。朱皖宜等[6]利用紫外和培养基中添加麦芽糖的诱变模式,获得了较出发菌株提高74%的突变株。张琴等[7]采用紫外、亚硝基胍、硫酸二乙酯和亚硝酸多次诱变,筛选到的突变株发酵水平提高了35%。Wang 等[8-9]通过对发酵培养条件的优化使得阿卡波糖的产量达到4 210 mg/L,而且在发酵不同时间补加葡萄糖、麦芽糖和黄豆饼粉混合物,阿卡波糖的产量提高到了4 878 mg/L。随着原生质体技术的发展,原生质体诱变技术被广泛应用于菌株的育种。由于原生质体没有细胞壁,对各种物理化学诱变较敏感,而又具有细胞的全能性及再生能力,所以是理想的诱变材料[10]。除了原生质体诱变外,以原生质体融合为基础的基因组重排技术也得到了广泛的应用,但是利用原生质体技术选育阿卡波糖高产菌株的报道相对较少。马妮[11]通过原生质体紫外、NTG诱变结合甲胺、麦芽糖耐受模型筛选得到1 株产量达3 700 mg/L 的突变株。郑斐[12]利用基因组重排技术得到的阿卡波糖高产融合子产量达到3 714 mg/L。
本实验对Actinoplanes utahensisZJB-08196 原生质体制备再生条件进行了研究,再对其原生质体进行了紫外诱变,取得了良好的效果。此外,原生质体制备再生也是基因组重排技术中非常关键的环节,本实验的研究内容也为进一步通过基因组重排技术选育阿卡波糖高产菌株奠定了基础。
1 材料与方法
1.1 材料
1.1.1 菌种
犹他游动放线菌(Actinoplanes utahensis)ZJB-08196,浙江工业大学生物工程研究所保藏。
1.1.2 培养基
固体培养基CPC(g/L):蔗糖30,蛋白胨2,酪氨酸1,K2HPO41,KCl 0.5,MgSO4·7H2O 0.5,FeSO4·7H2O 0.1,琼脂20,pH 自然。
菌丝培养基(g/L):葡萄糖10,酵母粉3,蛋白胨30,黄豆饼粉5,淀粉0.5,MgSO42,K2HPO40.5,pH 7.0。
再生培养基:R2YE 培养基[13]。
种子培养基(g/L):玉米淀粉16,黄豆饼粉40,甘油20,CaCO32,K2HPO40.5,pH 自然。
发酵培养基(g/L):麦芽糖浆271.4 mL,葡萄糖40,黄豆饼粉17,谷氨酸钠5,FeCI30.5,CaCO32.5,CaCl21.88,K2HPO41,甘油5,pH 6.5 ~7.0。
1.2.3 试剂
P 缓冲液:见参考文献[13];溶菌酶,购自Sigma 公司;牛血清白蛋白(BSA),购自上海聚源生物科技有限公司;L-脯氨酸,购自上海青析化工科技有限公司;聚乙烯吡咯烷酮K-30(PVP),购自汕头市陇西化工厂;麦芽糖浆,由杭州中美华东制药有限公司提供;乙腈,为色谱纯;其他试剂均为国产分析纯试剂。
1.1.4 仪器
SBA 40 E 生物传感器(山东微生物研究所);Osmometer Modal 3205 全自动冰点渗透压仪(Advanced);LC-10AT 型高效液相色谱仪及SPD-10A 检测器(Shimadzu)。
1.2 方法
1.2.1 菌丝培养及收集
从茄形瓶固体培养基上挑取1 cm2大小的菌落接种到菌丝培养基中,28 ℃培养56 h 后,取6 mL 转接到含甘氨酸的菌丝培养基(30 mL/250 mL 三角瓶)中进行二级培养。将二级培养液3 500 r/min 离心10 min 收集菌体,菌体首先用无菌水洗涤1 次,再用P缓冲液洗涤2 次,待用。
1.2.2 原生质体制备及再生
先将收集到的菌丝称重,根据菌丝体湿重加入适量的溶菌酶溶液,混合均匀。35 ℃恒温水浴,摇床转速120 r/min,每隔30 min 吸取酶解液并用相差显微镜观察原生质体释放情况。酶解结束后,用脱脂棉纯化原生质体,3 500 r/min 离心10 min 收集原生质体,最后用P 缓冲液洗涤2 次。
将制备好的原生质体悬液,用P 缓冲液稀释适当倍数,轻轻涂布于固体再生培养基上,28 ℃恒温培养5d,菌落计数,计算原生质体形成率和再生率。
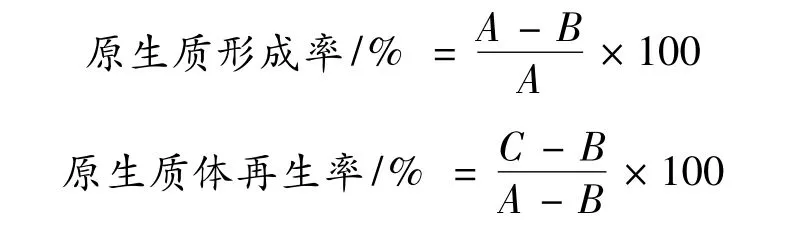
式中,A:总菌落数,酶处理之前的菌悬液涂布于CPC 平板生长的菌落。
B:未原生质体化菌落,酶解结束后,酶解混合液用蒸馏水稀释,涂布CPC 平板生长的菌落。
C:再生菌落,纯化后的原生质体经P 缓冲液稀释,涂布于再生培养基平板生长的菌落。
1.2.3 原生质体显微镜计数方法
用规格为25 cm×16 cm 的血细胞计数板在相差显微镜下直接计数。
1.2.4 干重和湿重测量方法
准确取10 mL 培养液于离心管,3 500 r/min 离心10 min,弃掉上清液,于90 ℃烘箱中烘干至恒重,称量干重,每样做3 个平行,取均值。
准确取10 mL 培养液于离心管,3 500 r/min 离心10 min,弃掉上清液,称量湿重,每样做3 个平行,取均值。
1.2.5 原生质体紫外诱变
将原生质体悬液的浓度调整为106个/mL,取5 mL 悬液于直径6 cm 的平板中,放置于2 根8 W 的紫外灯管下30 cm 处进行诱变。分别照射40、60、80、100、120 s,黑暗放置2 h 后,涂布再生培养基于28 ℃恒温再生。
致死率计算。根据各诱变组再生菌落数(M)和对照组再生菌落数(N),计算不同诱变时间处理后的致死率:
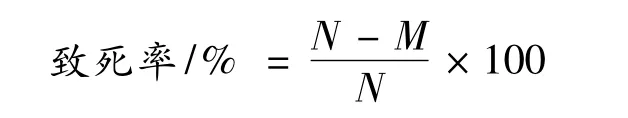
1.2.6 高产突变株筛选
再生菌落先转接于茄形瓶,再从茄形瓶中挑取1 cm2大小的菌落接种于100 mL 种子培养基中28 ℃,200 r/min 振荡培养72 h,以10%转接量转接于50 mL 发酵培养基中,于28 ℃,200 r/min 振荡培养7 d,取样测定阿卡波糖含量。
1.2.7 检测方法
发酵液渗透压测定:发酵液12 000 r/min 离心10 min,取上清液250 μL 测定渗透压。
葡萄糖测定:生物传感器检测[8]。
阿卡波糖检测:HPLC 检测[14]。色谱柱Luna NH2(4.6 mm×250 mm,5 μm);流动相:乙腈-磷酸盐缓冲液(75 ∶25);检测波长210 nm,流速1.0 mL/min,柱温40 ℃。
2 结果与分析
2.1 甘氨酸对菌丝生长和原生质体形成的影响
在放线菌的菌丝培养过程中添加甘氨酸,可以增加溶菌酶对菌丝细胞壁的敏感性。因为甘氨酸在放线菌细胞壁合成过程中,甘氨酸可以代替结构相似的丙氨酸从而干扰细胞壁的正常合成,有助于溶菌酶瓦解细胞壁。而甘氨酸的加入时间各异,有的放线菌需要刚开始培养时就加入[15],有的放线菌则在对数期再加入,然后继续培养一定时间,才能达到理想的酶解效果。图1 是菌丝培养初始就加入不同浓度的甘氨酸对菌株生长曲线的影响,并以不加甘氨酸为对照。从图中可以看出,甘氨酸对ZJB-08196 的抑制非常明显,当甘氨酸的浓度为3 g/L 时,菌株生长已经受到强烈的抑制,而浓度增加到5 g/L 时,几乎已经无法生长。从对照组的生长趋势可以看出,菌体生长过程中适应期较长,如果初始就加入甘氨酸,则菌体在适应期就会受到抑制,从而显著地影响后续生长,不能获得原生质体制备所需足量的菌体。所以,本实验选择二级培养的方式进行菌体培养,即菌体一级培养56 h 左右到达对数生长后期,再以20%(v/v)的接种量转接到含甘氨酸的新鲜菌丝培养基中进行二级培养。
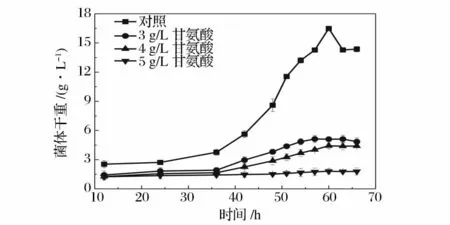
图1 不同浓度甘氨酸浓度下ZJB-08196的一级生长曲线Fig.1 Growth curves of ZJB-08196 in first stage of different glycine concentration
放线菌通常以甘氨酸和溶菌酶配合使用来制备原生质体,但是不同的品系所需的甘氨酸浓度差别较大,浓度从1 g/L 到15 g/L 都有报道[16]。表1 显示的是二级培养时不同浓度的甘氨酸对菌体生长和原生质体释放的影响。为了便于直接比较,酶解时菌体(湿菌体)质量浓度都调整为0.1 g/mL。从表中可知,当甘氨酸浓度为4 g/L 时,酶解得到的原生质体数最大达到2.88 ×107个/mL。
2.2 菌龄对原生质体形成和再生的影响
不同生长阶段的菌体对溶菌酶的敏感性是不同的,而且由于生理活性的差异也会导致再生效果的不同。一般原生质体的制备选择处于对数生长中后期的菌体,这是由于此阶段的菌体细胞壁中的肽聚糖含量较低,对溶菌酶的作用最敏感,而且原生质体修复能力也较强,再生效果好。而对数生长早期的菌体相对脆弱,酶的过度作用会影响原生质体的再生[17]。图2 是菌体在含4 g/L 甘氨酸菌丝培养基中的二级生长曲线,从图中可以看出,菌体在24 h 时进入稳定期。图3 是菌丝二级培养时间对原生质体形成和再生的影响,可以看出,原生质体的形成率随菌龄的增加缓慢增加,在20 h 处达到最大,随后下降。再生率趋势与形成率基本一致,但是在22 h 达到最大值。综合考虑形成率与再生率,菌体的最佳菌龄为22 h。而22 h 是菌体生长的对数后期,这表明处于对数生长后期的菌体最适合制备原生质体。已有过文献报道,处于对数后期或稳定期的放线菌菌体制备的原生质体有较高的再生能力[18]。
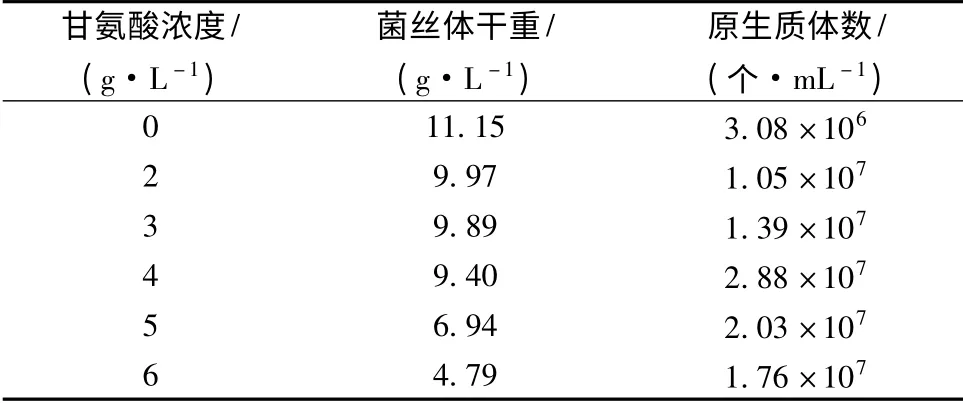
表1 不同浓度甘氨酸对菌丝生长和原生质体形成的影响Table 1 The effect of glycine concentration on mycelial growth and protoplast formation
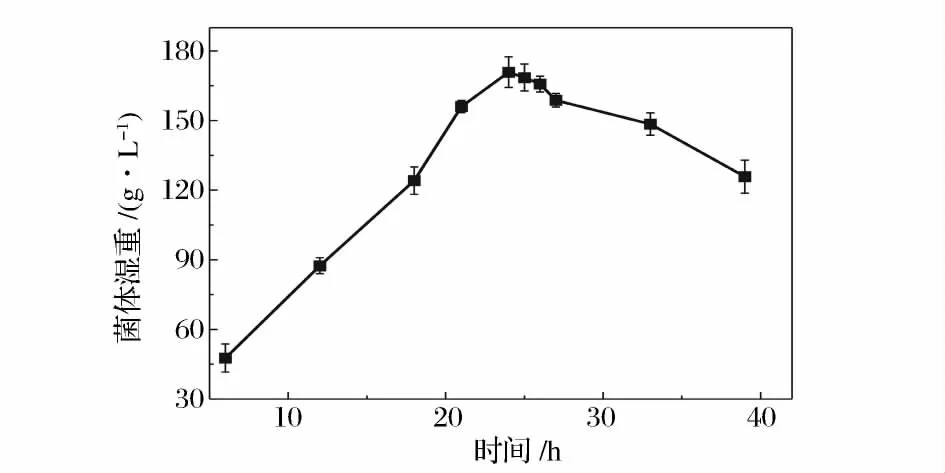
图2 ZJB-08196 的二级生长曲线Fig.2 Growth curves of ZJB-08196 in second stage
2.3 酶浓度、酶解时间、酶解温度和菌体(湿菌体)质量浓度对原生质体的形成和再生的影响
细菌和放线菌细胞壁的主要成分是肽聚糖,可以用溶菌酶来水解细胞壁。由于不同菌种品系的细胞壁各组分比例以及生理状态的差异,还有酶自身的纯度及性质的不同,所以要求的酶浓度以及酶解时间不尽相同。用于水解细胞壁的酶浓度过低,水解缓慢,不利于原生质体的形成,而浓度过高,处理时间过长,菌体表面的细胞壁水解太彻底,又会影响原生质体的活性,致使再生率下降。从图4 -A 酶浓度对原生质体形成和再生影响可以看出,随着酶浓度的增加,原生质体的形成率逐渐增加,再生率则在10 mg/mL 达到最大值,综合考虑形成率和再生率,最佳的酶浓度为10 mg/mL。酶解温度要兼顾酶的活力和对原生质体活性的影响。文献报道放线菌的最适酶解温度一般在30 ℃到37 ℃之间[19]。从图4 -B 可以看出,原生质体的形成率和制备率同时在35 ℃时达到最大,所以酶解的最适温度为35 ℃。从图4 -C 的酶解时长对原生质体形成和再生影响可以看出,虽然酶解时间越长形成率越高,但是超过4 h 后,原生质体的再生率显著下降,所以最终选择4 h 为酶解时长。
在确定的酶浓度、酶解时间及酶解温度条件下,酶解液中菌体的质量浓度也会影响原生质体的形成和再生,所以酶解时混合液中的菌丝体量要加以控制。如图4 -D 所示,菌体浓度小于0.2 g/mL 时,原生质体的形成率下降幅度并不显著,而再生率则逐渐增大,在0.2 g/mL 左右时达到最大,所以最佳的菌体质量浓度为0.2 g/mL 左右。
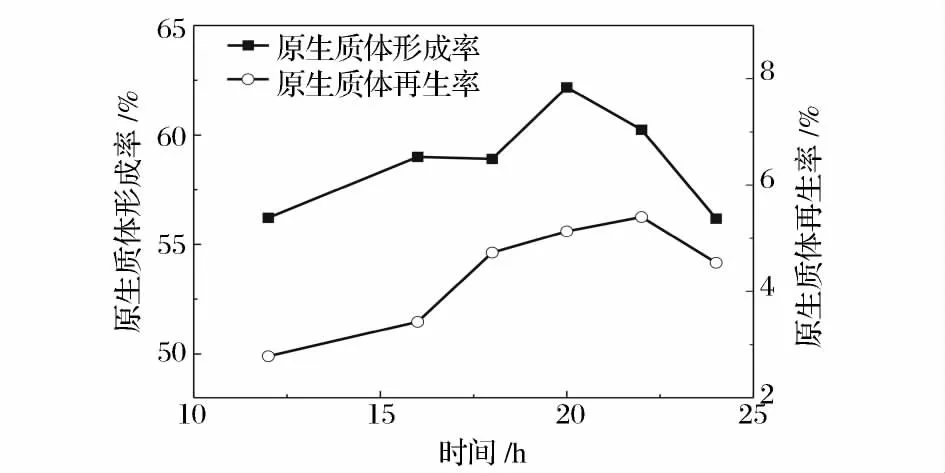
图3 菌龄对原生质体形成和再生的影响Fig.3 The effect of mycelium age on protoplast formation and regeneration
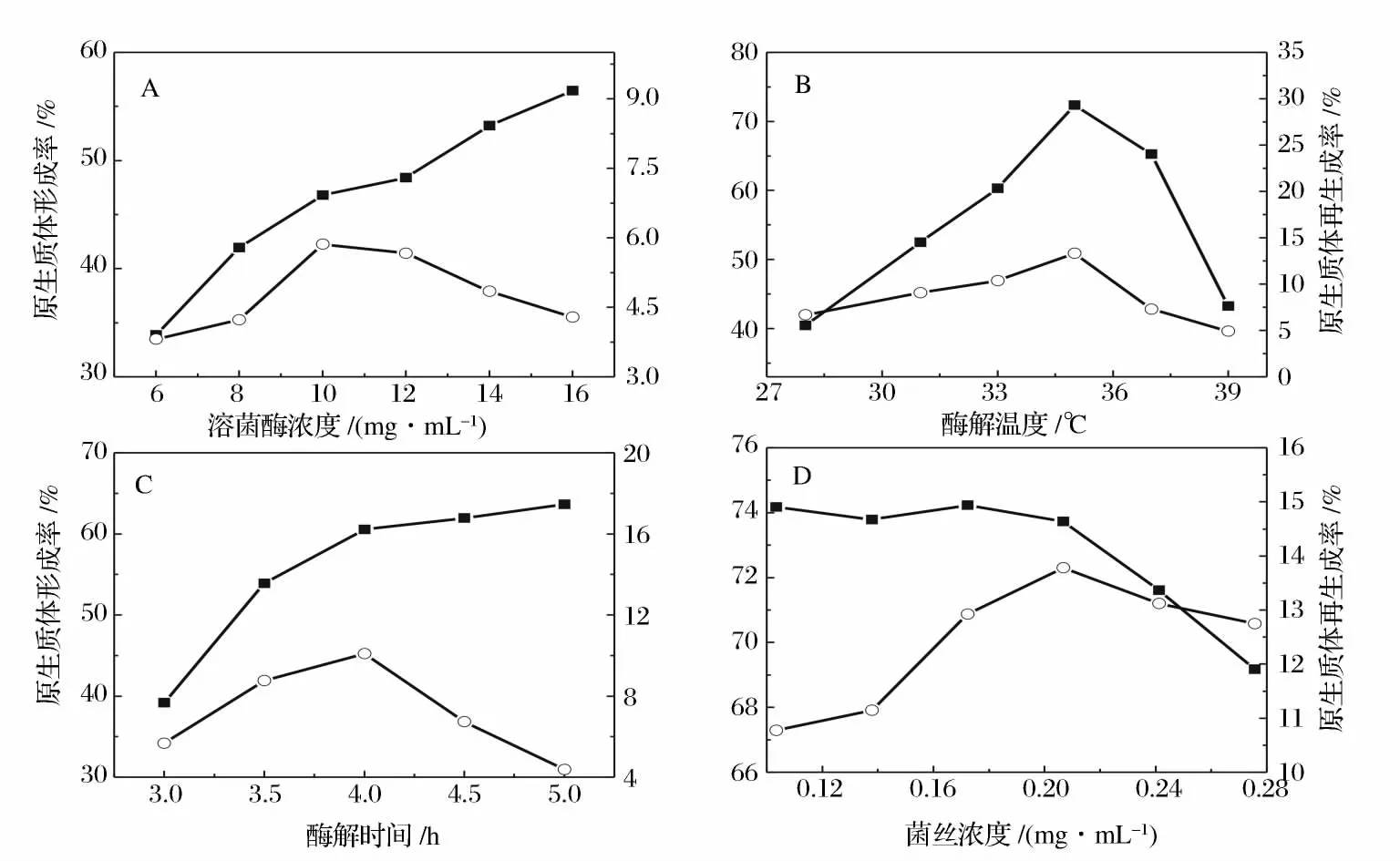
图4 溶菌酶浓度、酶解温度、酶解时间和菌体浓度对原生质体形成和再生的影响。Fig. 4 The effect of lysozyme concentration,enzymolysis time,enzymolysis temperature and mycelial concentration on protoplast formation and regeneration
2.4 原生质体再生培养基的优化
2.4.1L-脯氨酸对再生的影响
L-脯氨酸是再生培养基中非常重要的组成成分,可以用来合成细胞壁中肽聚糖,促进原生质体的再生。从表2 中可知,当L-脯氨酸的浓度达到4 g/L时,再生率最高达到10.6%,继续增加L-脯氨酸的浓度,再生率反而下降,因此最佳的L-脯氨酸浓度为4 g/L。

表2 再生培养基中L 脯氨酸浓度对原生质体再生的影响Table 2 The effect of L-proline concentration of regeneration medium on protoplast regeneration
2.4.2 BSA 和PVP 对再生的影响
BSA 一方面对原生质体起保护作用,可以保持原生质体结构和功能的完整性[20]。另一方面,BSA 也是一种营养物质,可以通过代谢转化为细胞壁合成的前体物质,加速细胞壁的再生。如表3 所示,在再生培养基中添加BSA 可有效提高再生率,当BSA 浓度达到0.3 g/L 时,再生率最高达到15.41%。
PVP 也是一种原生质体的保护剂,可以给原生质体提供稳定的代谢环境,促进原生质体的再生。如表3 所示,再生培养基中加入PVP 也可以提高原生质体的再生率,当浓度达到0.2 g/L 时,再生率最高为15.14%。从结果来看,添加BSA 后的最大再生率要稍高于PVP,但是BSA 价格较PVP 要高,结合经济性来考虑,PVP 是BSA 的良好替代品。
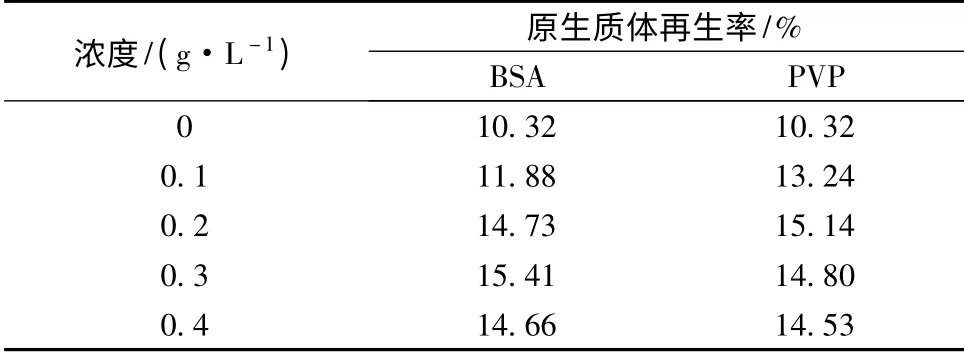
表3 再生培养基中BSA 和PVP 对原生质体再生影响Table 3 The effect of BSA and PVP of regeneration medium on protoplast regeneration
2.5 原生质体紫外诱变突变株筛选
原生质体紫外诱变致死曲线如图5 所示,原生质体的致死率随时间的延长迅速增加,当紫外照射时间为60 s 时,致死率为74.9%,当时间达到100 s 时,致死率已经达到95.5%,照射120 s 后原生质体的致死率达到100%。原生质体的致死率在40 s 到60 s 时段内变化幅度较大,这表明在此阶段紫外照射的影响最大。
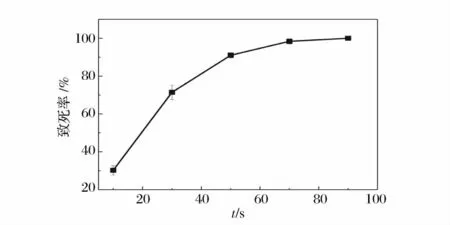
图5 不同紫外诱变时间原生质体致死率Fig.5 The lethality rate of protoplasts treated for different mutation time
从再生平板中挑取55 株诱变株,进行发酵培养。挑选出了7 株阿卡波糖产量提高10%的突变株,选择其中2 株产量超过5 000 mg/L 的菌株进行传代培养验证其遗传稳定性。经过5 代连续培养后,得到了1 株阿卡波糖产量稳定的菌株UN-52,其阿卡波糖产量达到5 165.2 mg/L。出发菌株ZJB-08196 产量为4 518.7 mg/L,UN-52 与出发菌株相比产量提高了约12%。UN-52 传代稳定性结果如表4 所示。
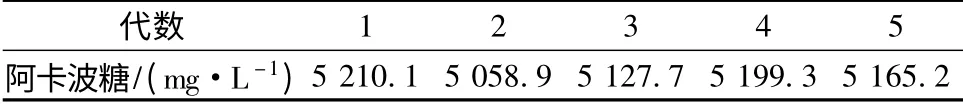
表4 传代稳定性验证结果Table 4 Verification of the mutation strain UN-52
2.6 诱变菌株和出发菌株的发酵特性比较
为了比较突变株UN-52 与出发菌株发酵特性,监测了2 株菌发酵过程中阿卡波糖、渗透压、生物量和葡萄糖随时间的变化,结果如图6 所示。从图中可以看出,随着发酵时间的延长渗透压和葡萄糖在逐渐降低,而阿卡波糖和生物量相应地逐渐增加。而2 株菌的发酵曲线对比可以看出,突变株UN-52 发酵过程中渗透压和葡萄糖下降得更快,而其生物量与阿卡波糖产量的增长则快于出发菌株。这说明突变株UN-52 相较于出发菌株来说,其对发酵培养基中的葡萄糖等营养物质利用得更快,所以其长势更加旺盛,阿卡波糖产量也更高。
3 讨论
原生质体的制备和再生涉及到多种影响因素,要对各影响因素综合考虑,才能获得良好的制备与再生效果。甘氨酸对菌体的生长有明显的抑制作用,先将菌体培养至生长旺盛的对数期,再转接入含甘氨酸的培养基继续培养则可以减轻抑制作用得到足够的菌体。菌体的生理状态对原生质体制备和再生有一定影响,二级培养22 h 时菌体处于生长旺盛的对数期,菌体中大部分为新生菌丝,这样更加有利于溶菌酶的酶解。在酶解过程中控制菌体的浓度也是十分必要的,菌体浓度过大,酶解体系粘度较大会影响原生质体的释放,菌体浓度过小,则细胞壁酶解过度难于再生。
已有大量文献报道过,再生培养基中添加胶体物质,如明胶、聚乙烯吡咯烷酮、琼脂糖、藻酸钙等,可以有效提高原生质体的再生率[21]。一方面,这些胶体物质可以稳定原生质体的生长环境以促进再生。另一方面,胶体产生的胶体渗透压可以防止环境中的水分渗透进入原生质体扰乱其生长[22]。本实验结合操作方便性和经济因素,最后选择添加0.2 g/L PVP,原生质体的再生率达到15.14%。
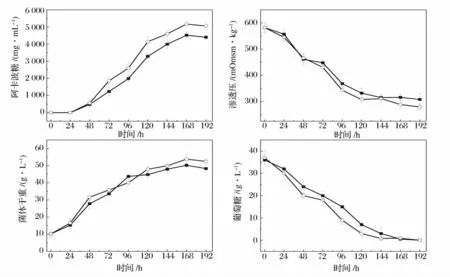
图6 ZJB-08196 和NU-52 发酵过程中阿卡波糖、渗透压、生物量和葡萄糖的变化Fig.6 The course of acarbose,osmolality,biomass and glucose by ZJB-08196 and UN-52
本实验进一步将Actinoplanes utahensisZJB-08196原生质体通过紫外诱变的方法进行诱变育种,获得了1 株比出发菌株提高了12%的突变株UN-52,产量达到了5 165.2 mg/L。突变株UN-52 产阿卡波糖的能力高于已报道的马妮[11]等人通过育种得到的菌株。通过发酵过程的比较表明,突变株UN-52 对营养物质利用得更快,如果在发酵过程中补加营养物质应该能够进一步提高产量,下一步工作主要是突变株发酵条件优化、补料策略的研究。
[1] Campbell L K,White J R,Campbell R K. Acarbose:Its role in the treatment of diabetes mellitus[J]. Annals of Pharmacotherapy,1996,30(11):1 255 -1 262.
[2] 何素婷,许激扬,陈代杰. 具有α-葡糖苷酶抑制作用的抗糖尿病药物[J]. 工业微生物,2003,33(1):43 -49.
[3] Liu R L,Ding M P,Xu W,et al. Progress on glucosidase inhibitors[J]. Pharmaceutical Biotechnology,2009,16(4):388 -392.
[4] 程新,徐波,魏塞金,等. 一株产α-糖苷酶抑制剂的稀有放线菌分离与鉴定[J]. 食品与发酵工业,2008,34(9):58 -60.
[5] Wei S J,Cheng X,Huang L,et al. Medium optimization for acarbose production byActinoplanessp.A56 using the response surface methodology [J]. Africa Journal of Biotechnology,2010,9(13):1 949 -1 954.
[6] 朱皖宜,陶美珍,邢红燕,等. 阿卡波糖产生菌的诱变育种[J]. 中国医药工业杂志,2005,36(1):17 -21.
[7] 张琴,胡海峰,朱宝泉. 阿卡波糖产生菌的选育和发酵工艺优化[J]. 中国医药工业杂志,2008,39(11):820-822.
[8] Wang Y J,Liu L L,Feng Z H,et al. Optimization of media composition and culture conditions for acarbose production byActinoplanes utahensiaZJB-08196[J]. World Journal of Microbiology and Biotechnology,2011,27(12):2 759 -2 766.
[9] Wang Y J,Liu L L,Wang Y S,et al. Actinoplanes utahensis ZJB-08196 fed-batch fermentation at elevated osmolality for enhancing acarbose production[J]. Bioresource Technology,2012,103(1):337 -342.
[10] 施巧琴,吴松刚. 工业微生物育种学(第三版)[M].北京:科学出版社,2009:222 -223.
[11] 马妮. 阿卡波糖产生菌育种新方法的研究与应用[D]. 沈阳:沈阳药科大学,2006.
[12] 郑斐. 原生质体技术在阿卡波糖产生菌育种中的应用[D]. 沈阳:沈阳药科大学,2008.
[13] Hopwood D A,Bibb M J,Chater K F,et al. Genetic Manipulation of Streptomyces,a laboratory manual[M].England:John Innes Foundation press,1985:365.
[14] Wang Y J,Xue Y P,Zheng Y G,et al. Analysis and determination of anti-diabetes drug acarbose and its structural analogs[J]. Current Pharmaceutical Analysis,2011,7(1):12 -20.
[15] Carneiro-da-Cunha M G,Filho J L L,Campos-Takaki G M. Protoplast formation and regeneration fromStreptomyces clavuligenrusNRRL 3585 and clavulanic acid production[J]. Brazilian Journal of Microbiology,2002,33(4):347 -351.
[16] Allison D G,Roberts P M. Protoplast formation and reversion inActinomadura[J]. Applied Microbiology and Biotechnology,1998,28(6):580 -582.
[17] 刘志恒,姜成林. 放线菌现代生物学与生物技术[M].北京:科学出版社,2004:128 -129.
[18] Lyutzkanova D,Nikolova B,Stoilova-Discheva M,et al.Protoplast formation and regeneration inStreptomyces flavopersicus[J]. Letters in Applied Microbiology,1993,16(4):217 -219.
[19] 陈飞,关艳丽,吴红艳,等. 链霉菌11371 原生质体制备与再生[J]. 微生物学杂志,2011,31(5):54 -58.
[20] Illing G T,Normansell D,Peberdy J F. Protoplast isolation and regeneration inStreptomyces clavuligerus[J]. Journal of General Microbiology,1989,135(8):2 289.
[21] Miyamoto T,Nishioka K,Kataoka K,et al. Protoplast formation and regeneration in some lactic acid bacteria[J].Japanese Journal of Zootechnical Science,1990,61(6):481 -486.
[22] Osamu T,Sadahiro O. Efficient protoplast regeneration for some homofermentative lactobacilli and pediococci[J].Archives Microbiology,2001,177(1):36 -40.
