茄子种质资源遗传多样性的形态标记分析
2013-09-03王志敏于晓虎丁泽琴汤青林田时炳王永清杨
张 念 王志敏 于晓虎 丁泽琴 汤青林 田时炳 王永清杨 洋 宋 明*
(1西南大学园艺园林学院,南方山地园艺学教育部重点实验室,重庆市蔬菜学重点实验室,重庆400715;2重庆市农业科学院蔬菜花卉研究所,重庆 400055)
种质资源遗传多样性研究对于揭示物种演化过程和进化潜能、了解群体遗传结构及多态性有重要价值,从而为物种起源研究、亲本的选配、品种的分类及保护等提供依据(解新明和云锦风,2000)。形态标记是利用作物肉眼可见的、特定的、简便直观的外部特征,或者借助简单测试就可以识别的某些性状,主要包括农艺性状、植物学性状、工艺性状和抗性等进行作物标记的方法,具有简单直观的特点,是杂种鉴定和遗传研究领域常用的经典方法(钟淮钦 等,2005)。在杂种优势利用中,应用形态标记进行纯度鉴定、杂种鉴别和品种权益保护具有重要实践意义(涂金星和傅廷栋,2001)。
茄子(Solanum melongenaL.)起源于亚洲南部热带地区,古印度为其最早的驯化地,中国栽培茄子的历史悠久,是茄子的第二起源地,拥有丰富的茄子种质资源。随着分子生物技术的快速发展,各种生物化学和分子标记已经被广泛地应用于茄子种质资源的鉴定和分类研究。卢婷等(2008)用25对SSR引物对66份茄子材料的遗传多样性进行了分析,认为茄子种间遗传距离较大,栽培种内遗传基础相对狭窄。毛伟海等(2006)用ISSR标记从分子水平对南方长茄资源的遗传多样性进行了分析,也得出类似的结论。牛艳秀等(2010)应用WRKY转录因子分子标记构建了34个茄子品种的指纹图谱,用遗传相似性分析将供试品种大体分为5个类群。但是目前茄子的分子标记存在标记数目有限、分子遗传图谱密度不够饱和、不能覆盖整个基因组等问题。形态性状的鉴定和描述更直接、简单,有利于在田间进行直接选择,目前仍是种质资源研究的最基本的方法和途径。Polignano等(2010)对茄子属及其近缘种属的98份材料进行种间及种内的多样性研究,对农艺性状进行定量测定和定性描述,使用主成分分析将它们聚类为3个差异显著的类群,并将有关果实的描述单独作为一个分类变量。李树贤等(2009)发现同源四倍体茄子株形指数与平均结果数呈很强的负相关性,相关遗传力及间接选择效率为:叶形指数>植株开展度>始花节位>茎粗>株高。形态学性状数据是种以上或种内分类的不可缺少的重要依据之一,尤其是茄子种质资源在形态上的变异非常显著,运用形态学鉴定仍然不失为一种合适的方法(Prohens et al.,2005)。本试验采用传统形态标记,对从国内外搜集的76份茄子材料的遗传多样性构成特性及亲缘关系进行了研究,旨在为茄子种质资源的充分利用和科学育种提供可行的参考。
1 材料与方法
1.1 材料
试验所用76份茄子种质资源由重庆市农业科学院蔬菜花卉研究所提供(表1)。
1.2 方法
1.2.1 形态学性状调查项目 试验于2012年4~8月在重庆市农业科学院蔬菜花卉研究所茄科蔬菜育种试验基地进行。采用随机完全区组设计,调查方法参照李锡香和朱德蔚(2005)关于茄子的农艺性状描述及赋值标准。每小区选取5株生长状况较一致的茄子样本,使用游标卡尺、直尺、卷尺、比色卡、天平等测量工具,对27个形态学性状(花部性状3个,叶部性状9个,茎部性状2个,果实性状12个,植株性状1个)(表2)进行调查,记录原始数据后取平均值。
1.2.2 表型性状赋值 性状赋值取决于其类型,叶片长度、单果质量等数量性状直接取其测量值,而果皮色、叶形等非数据型质量性状(有序多态性状)的赋值则根据不同表现型,按状态的等级依次编码为0、1、2……具体赋值标准参考李锡香和朱德蔚(2005)的文献。
1.2.3 数据处理 采用SPSS18.0数据分析软件,应用斯皮尔曼(Spearman)等级相关系数对部分表型性状进行双变量相关分析;系统聚类分析选择组间联接方法,以皮尔森(Pearson)相关性系数进行聚类。
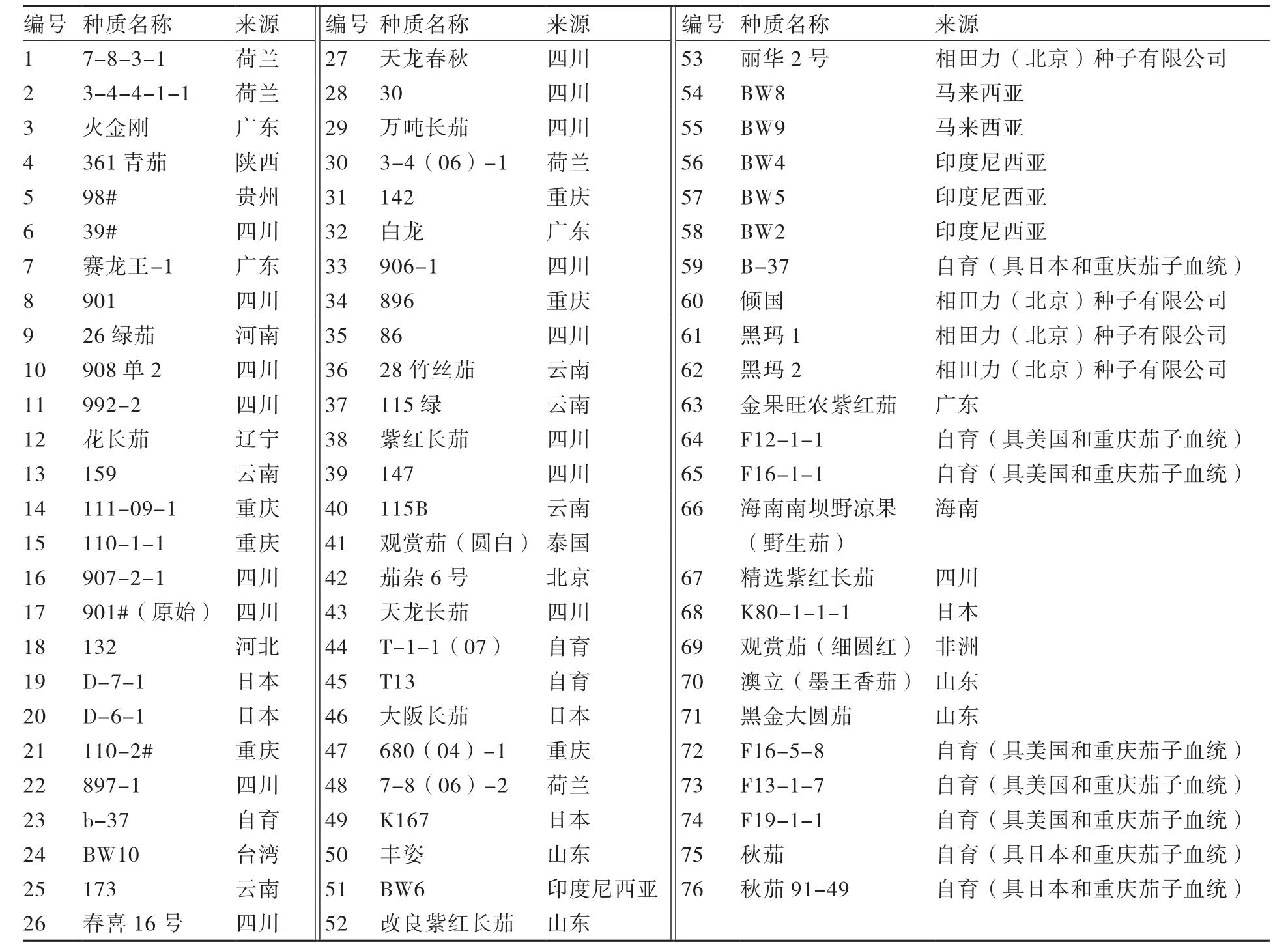
表1 供试茄子种质资源

表2 不同时期田间调查性状的分布
2 结果与分析
2.1 形态学性状基本数据分析
由表3可知,76份茄子种质所调查的27个形态学性状的变异系数中,叶刺最大,为493.3%;果面斑纹和果萼刺次之,分别为100.9%和82.4%;果实弯曲度变异系数最小,为10.9%。茄子近缘野生种及半栽培型茄子一般具有叶刺,该性状可作为茄子驯化及改良的农艺性状之一。在果实相关性状中,果面斑纹及果面斑纹色具有丰富的遗传变异。
对茄子9种果形(1扁圆、2圆球、3高圆、4卵圆、5长卵、6短筒、7长筒、8长条、9细长)及7种果皮颜色(1白、2白绿、3绿、4橘红、5鲜紫、6紫红、7黑紫)的出现频率进行统计分析(表4),76份种质中,果形为长卵形的种质有30份,占所有果形的39.5%,长卵、短筒、长筒以及长条果形的累计百分比为68.4%,说明供试材料中长棒状茄子所占的比例较大。
对果皮颜色(表5)的分析可知,商品果色为黑紫色的品种有40个,约占所有商品果色的52.6%,鲜紫、紫红、黑紫色果实的累计百分比为76.3%,供试茄子的商品果色以紫色为主。
2.2 形态学性状相关性分析
以Spearman等级相关系数对76份茄子材料选取有关花、叶、果实的7个农艺性状进行相关性分析(表6),其中花柱长度、果皮颜色与果形间呈极显著正相关,相关系数分别为0.542与0.471;叶长与果形间呈极显著负相关,相关系数为-0.421;花柱长度与叶长呈极显著负相关,相关系数为-0.402;花柱长度与首位花节、叶形指数、果皮颜色,叶形指数与果形、果皮颜色等性状间均呈显著正相关。除此之外,其他性状间的相关性总体水平都较低,相关系数在0.020~0.202之间。
2.3 形态学性状聚类分析
对76份茄子种质材料进行表型性状聚类,首先对各性状调查数据进行标准化转换(0-Z转换),采用Q型聚类(以个案分群),以皮尔森(Pearson)相关性系数为度量标准,使用组间联接法生成所有供试材料的树状图(图1)。
在Pearson相关性系数为23处将所有材料分为三大类群:第一大类群共包含25个品种:BW4、BW5、BW8、BW6、BW2、3-4(06)-1、30、观赏茄(细圆红)、海南南坝野凉果、观赏茄(圆白)、115绿、147、F16-1-1、秋茄、秋茄91-49、142、115B、澳立(墨王香茄)、39#、茄杂6号、361青茄、黑金大圆茄、908单2、28竹丝茄、906-1。其特点是果形多为圆球形以及短棒状或扁圆形。第二大类群包含25个品种:花长茄、159、173、BW10、BW9、金果旺农紫红茄、白龙、896、901、992-2、680(04)-1、b-37、万吨长茄、897-1、111-09-1、B-37、110-1-1、98#、86、901#(原始)、火金刚、赛龙王-1、132、改良紫红长茄、907-2-1。其特点是多为重庆、四川等国内本地品种,果实以长棒状和紫红色为主。第三大类群包含26个品种:F12-1-1、F16-5-8、丰姿、T-1-1(07)、3-4-4-1-1、7-8(06)-2、K167、F13-1-7、7-8-3-1、精选紫红长茄、春喜16号、紫红长茄、黑玛2、丽华2号、天龙春秋、110-2#、倾国、天龙长茄、D-7-1、D-6-1、黑玛1、T13、大阪长茄、F19-1-1、K80-1-1-1、26绿茄。其特点是多为引种或具国外血统的杂交种,果实以长棒状和紫黑色为主。
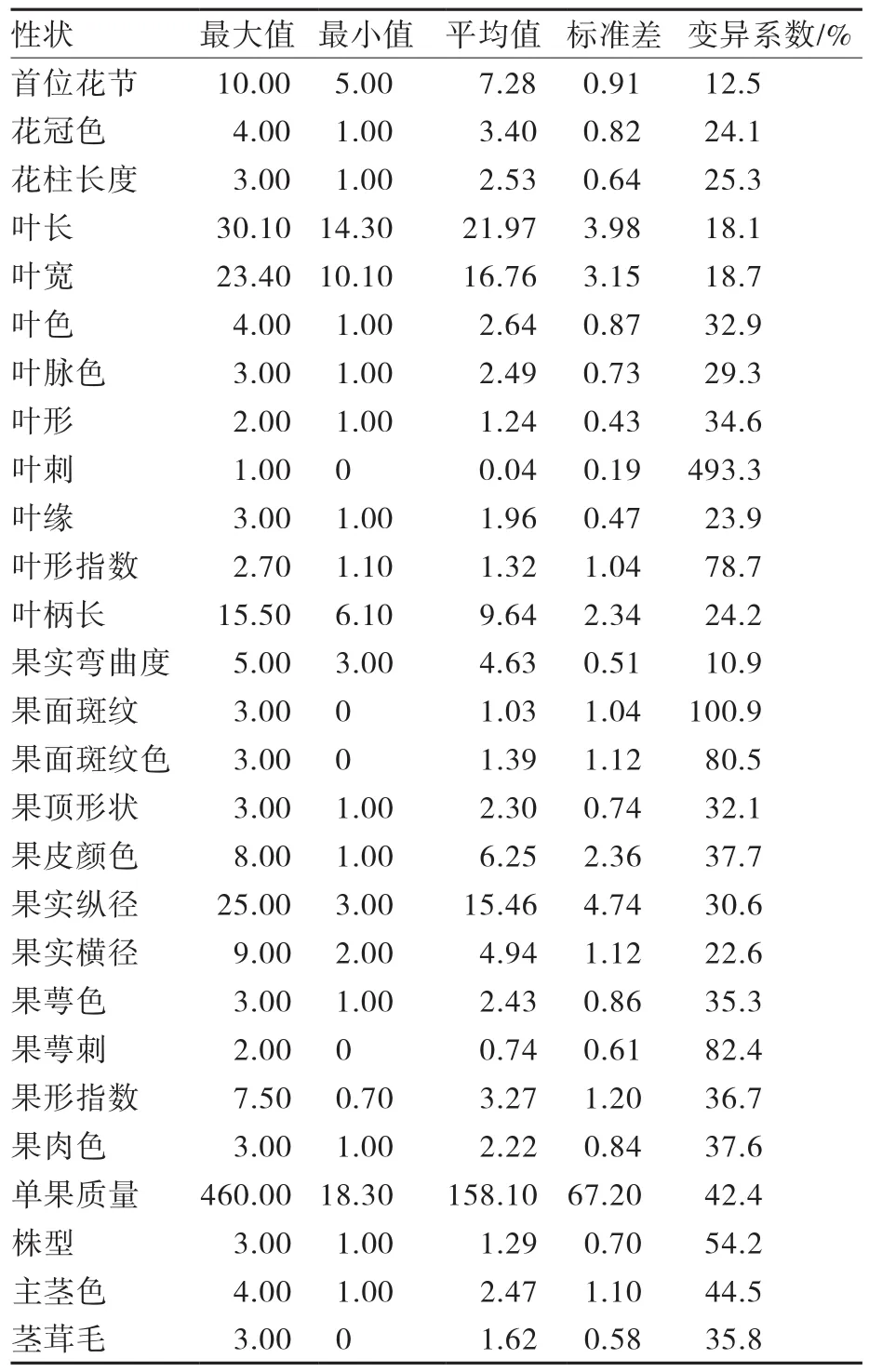
表3 供试茄子材料形态多样性的统计数据
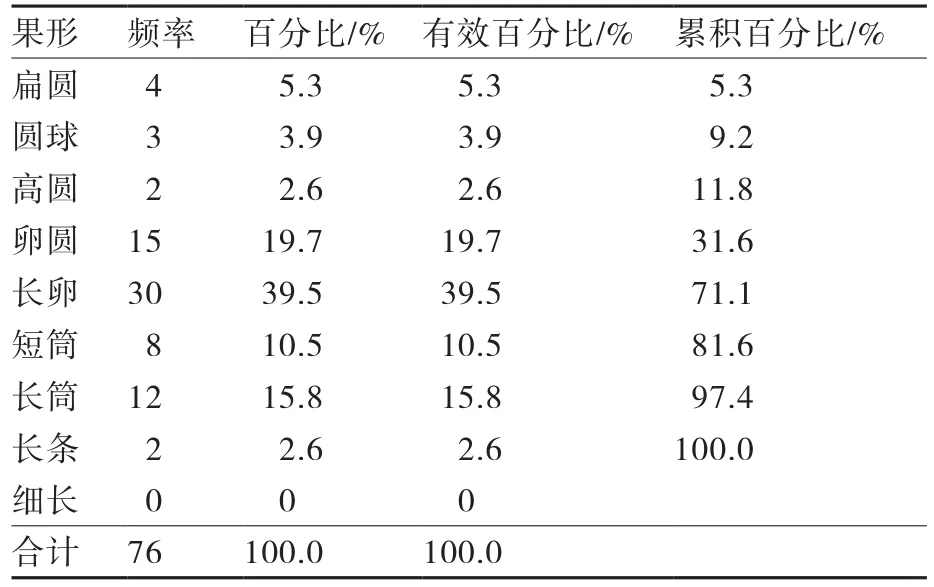
表4 供试茄子材料9种果形性状的频率分布

表5 供试茄子材料7种果皮颜色性状的频率分布

表6 供试茄子材料7个表型性状的相关性分析
在Pearson相关性系数19.5处可将供试材料分为七个类群:
第一类群共包含10份种质:BW4、BW5、BW8、BW6、BW2、观赏茄(细圆红)、3-4(06)-1、30、海南南坝野凉果、观赏茄(圆白)。这些材料的表型性状与其他种质间差异均较大,被聚为一类。其中BW4、BW5、BW8、BW6、BW2果实为绿色,果形短棒状或扁圆形,聚为一个亚类。观赏茄(细圆红)、海南南坝野凉果、观赏茄(圆白)等为近缘野生种,果实为圆形小果,白或橘红色。第二类群共包含15份种质:115绿、147、F16-1-1、秋茄、秋茄91-49、142、115B、澳立(墨王香茄)、39#、茄杂6号、361青茄、黑金大圆茄、908单2、28竹丝茄、906-1。该类群的材料中包含多数圆球形、扁圆形茄子,以及部分短棒状茄子。可进一步划分为三个亚类:第一亚类包括115绿、147、F16-1-1 3个品种,其一致特征主要是果实卵圆形,果萼绿紫色;第二亚类包括秋茄、秋茄91-49、142、115B、澳立(墨王香茄)等品种,多为黑紫色棒状茄,花冠色及叶形基本一致;第三个亚类包括39#、茄杂6号、361青茄、黑金大圆茄、908单2、28竹丝茄、906-1等品种,果皮色多为紫色或黑紫色,株型及茎茸毛等表型一致。第三类群共包含8份种质:花长茄、159、173、BW10、BW9、金果旺农紫红茄、白龙、896。该类群内主要包括一些西南地区以外的紫红色长棒状茄子,可分为两个亚类:前7个品种聚为第一亚类,其果实为棒状,花冠均为浅紫色。第二亚类896果实为长棒状,花柱类型为花柱长于雄蕊。第四类群共包含16份种质:901、992-2、680(04)-1、b-37、万吨长茄、897-1、111-09-1、B-37、110-1-1、98#、86、901#(原始)、火金刚、赛龙王-1、132、改良紫红长茄。该组中各材料的果实以黑紫色、长棒状为主,材料主要来源于四川及重庆。第五类907-2-1单独聚为一类。第六类群共包含8份种质:F12-1-1、F16-5-8、丰姿、T-1-1(07)、3-4-4-1-1、7-8(06)-2、K167、F13-1-7。该组包含了美国与重庆地方茄子的杂交种、具美国和日本血统的中国茄子以及荷兰引种的部分茄子,这些材料的果实以棒状、黑紫色为主,果萼绿色,果面无斑纹。第七类群共包含18份种质:7-8-3-1、精选紫红长茄、春喜16号、紫红长茄、黑玛2、丽华2号、天龙春秋、110-2#、倾国、天龙长茄、D-7-1、D-6-1、黑玛1、T13、大阪长茄、F19-1-1、K80-1-1-1、26绿茄。该类群内的材料包含4份日本引种或具日本血统的茄子,以及5份一代杂种、2份国外引种、7份四川地方品种。这些材料果实多为黑紫色、长棒状以及中早熟。

图1 使用平均联接(组间)的树状图
3 结论与讨论
茄子主要集中在亚洲种植,近些年地区间的引种频繁,不同生态类型间的基因交流广泛,因此单纯依据果实的形状和地理起源分类已经不能真实地反映材料间的遗传背景和遗传差异(廖毅 等,2009)。本试验通过对76份茄子种质资源的株型、叶色、果形指数等27个形态学性状进行调查,结果发现各表型性状的变异系数中,叶刺最大,为493.3%,果面斑纹和果萼刺次之,果实弯曲度变异系数最小,为10.9%;对76份茄子材料选取有关花、叶、果实的7个农艺性状进行相关性分析,其中花柱长度、果皮颜色与果形间呈极显著正相关,叶长与果形间呈极显著负相关,花柱长度与叶长呈极显著负相关。多元统计法分析中的许多步骤,如性状的选取、数量标准化以及采用何种聚类方法等,都具有一定的主观性,不同方法往往产生不同的结果,如果性状选取不合理,即使记录可靠、运算精确并且采用较好的计算方法,也得不到正确的分类结果。可见性状的同源性、稳定性和相关性是数量分类中性状选取问题的核心(连勇 等,2006)。本试验在选取分类性状时着重考虑了这些方面的因素,选取的性状多为花器官及果实特征,选用了27个较为稳定的性状作为标记来进行多样性的研究(易金鑫,2000)。
对调查结果进行聚类分析,以种质间Pearson相关性系数为标准将供试材料聚为了三大类群和七个类群,具有相同来源、相似生态类型及亲缘关系较近的材料大多聚为同一类。从聚类结果看,栽培茄的分类既保持了传统上以果实性状作为重要依据的基础,也发生了性状相互交替聚类的现象,可以客观反映目前栽培种茄子在人工选择前提下的遗传变异的方向。聚类分析较好地区分出了国外的部分种质、具有国外茄子血统的种质以及近缘野生种材料,而对国内地方品种和杂种一代茄子材料的划分不够理想,可能与这些材料间的遗传基础较窄有关。茄子在西南地区栽培广泛,基因交流和自然变异比较快,茄子遗传育种中应注重对原始资源的保存与利用,育种工作中除了对目标性状的变异保留外,也需要对其他变异有所针对性地进行选择和保存,从而为茄子育种积累更多自然材料和性状。
李树贤,吴志娟,杨志刚,李明珠,赵萍.2009.同源四倍体茄子育种的选择Ⅱ.株形指数的遗传多样性及其相关选择.中国农学通报,25(7):183-187.
李锡香,朱德蔚.2005.茄子种质资源描述规范和数据标准.北京:中国农业出版社.
连勇,刘富中,陈钰辉.2006.我国茄子地方品种类型分布及种质资源研究进展.中国蔬菜,(10):9-14.
廖毅,孙保娟,黎振兴,罗少波,陈日远,孙光闻,李植良.2009.茄子及其近缘野生种遗传多样性及亲缘关系的AFLP分析.热带作物学报,30 (6):781-786.
卢婷,汪国平,林明宝,方木壬.2008.应用SSR 标记分析茄子种质资源的遗传多样性.中国蔬菜,( s):5-9.
毛伟海,杜黎明,包崇来,胡天华,朱琴妹,胡海娇.2006.我国南方长茄种质资源的ISSR标记分析.园艺学报,33(5):1109-1112.
牛艳秀,赵福宽,孙清鹏,杨爱珍.2010.茄子品种基于WRKY转录因子的指纹图谱构建及遗传相似性分析.农业科技通讯,(5):112-116.
涂金星,傅廷栋.2001.甘蓝型油菜几个苗期形态标记的遗传.华中农业大学学报,20(4):318-320.
解新明,云锦风.2000.植物遗传多样性及其检测方法.中国草地,(6):51-59.
易金鑫.2000.亚洲部分茄子品种资源数量分类.园艺学报,27(5):345-350.
钟淮钦,李富生,杨清辉.2005.甘蔗杂种真实性鉴定研究综述.中国农学通报,21(6):390-394.
Polignano G,Uggenti P,Bisignano V,Gatta D C.2010.Genetic divergence analysis in eggplant (Solanum melongenaL.) and allied species. Genet Resour Crop Evol,57:171-181.
Prohens J,Blanca J M,Nuez F.2005.Morphological and molecular variation in acollection of eggplants from a secondary center of diversity:implications for conservation and breeding.Journal of the American Society for Horticultural Science,130(1):54-63.
赵泓,柴敏,于栓仓,姜立纲,毛爱军.2004.胚珠培养获得秘鲁番茄与栽培番茄种间杂种具有非温敏抗根结线虫基因Mi-3.分子植物,2(6):833-838.
周国治,叶青静,王荣青,阮美颖,杨悦俭.2010.胚挽救和分子标记辅助筛选获得番茄抗根结线虫杂种.浙江大学学报,36(3):255-261.
Alexander L J.1963.Transfer of a dominant type of resistance to the four known Ohio pathogenic strains ofTobacco mosaic virus(TMV)fromLycopersicon peruvianumtoL. esculentum.Phytopathology,53:869.
Canady M A,Meglic V,Chetelat R T.2005.A library ofSolanum lycopersicoidesintrogression lines in cultivated tomato.Genome,48(4):685-697.
Doganlar S,Frary A S,Tanksley S D.1997.Production of interspecific F1hybrids,BC1,BC2and BC3populations betweenLycopersicon esculenturnand two accessions ofLycopersicon peruvianumcarrying new root-knot nematode resistance genes.Euphytica,95(2):203-207.
Liu Y S,Zamir D.1999.Second generationL.pennelliiintrogression lines and the concept of bin mapping.Tomato Genet Coop,49:26-30.
Monforte A J,Tanksley S D.2000.Development of a set of near isogenic and backcross recombinant inbred lines containing most of theLycopersicon hirsutumgenome in aL.esculentumgenetic background:a tool for gene mapping and gene discovery.Genome,43(5):803-813.
Rick C M.1986.Germplasm resources in the wild tomato species.Acta Hort,190:39-47.
Stommel J R.1992.Enzymic components of sucrose accumulation in the wild tomato speciesLycopersicon peruvianum.Plant Physiol,99(1):324-328.
