生命信息安全控制原理的探讨
2013-08-27谌南辉
谌南辉
(江西农业大学 动物科学技术学院,江西 南昌 330045)
1 免疫认知的历史沿革
早期的认知,因天花康复后不再患天花,得名“免疫”。随后发现相关事件不限于病原微生物,如过敏反应、移植物排斥、妊娠反应、输血反应等。由于这些非传染性因子,即非经典免疫元素导致认知的积淀,终于从微生物学脱颖而出,形成一门新学,即免疫学。
事物的潜态是事物发生的动因,而显态是由潜态所决定的现象反映。人们认知常以显态的表现去作潜态的诠释,即用现象去诠释和定义潜态的本质。比如认为排异是免疫的本质,对于肠道正常菌群不排斥,则用“耐受”来诠释。排异与耐受都是现象,而现象发生的真正动因(潜态)才是需要探究的。
得天花则为危,一旦康复则为安,不再患,古人曰:免疫。免疫是现象,事件之因是机体的化险为夷、转危为安的生命机制。
研究显示,免疫应答不仅针对病原相关分子模式(pathogen associated molecule patterns,PAMP)外源性成分,也可由自身物质所诱导,即自身正常细胞所发出的危险信号也会感召机体的应答。人们一直在讨论模式。从克隆选择学说、SNSD(self-nonsef determination)模式、双信号模式、双信号模式的延伸-协同刺激模式、INS模式,到新近的危险模式(danger model)[1]。从克隆选择学说,到SNSD(self-nonsef determination)模式、双信号模式、双信号模式的延伸-协同刺激模式、INS模式基本上是基于排异的思维,而新近的危险模式开始从自己与非己的观念冲破出来,出现了有意义的内容和新的思维。
2 生命信息识别是保障生命安全的第一要素
细菌(bacteria)和古细菌(archaea)有一套防御机制抵御外来侵入性因子,在成簇的、有规律间隔的多次重复短片段(clustered regularly interspaced short palindromic repeat,CRISPR)基础上建立适应性免疫系统,其效应酶蛋白Cas9清除外源DNA片段。该系统将外源病毒或质粒DNA短片段“集中”到细胞基因组特定区域上,将外源DNA作为一种记忆储存起来。这段DNA转录生成CRISPR前体RNA,前体RNA生成后被切割成一段一段的重复RNA片段,这些小RNA分子即成熟CRISPR RNA(crRNA)。crRNA招募CRISPR相关蛋白与被细胞记住的外源入侵DNA或mRNA片段结合,将它们彻底摧毁。病毒再次入侵时,细菌编码的Cas9因子发挥作用,消灭来犯病毒[2]。Cas9蛋白与crRNA和tracrRNA协同识别外来DNA分子,与之结合将其降解,该机制具有记忆性。生命为了自身的生存,如细菌那样,必须识别感知外来遗传信息物质,并以一种有效机制将其消除。高等生物存在更为丰富多彩的识别装置,TCR识别细胞内传递的危险信号;BCR、Ig感知体液环境中的危险信号。模式识别受体(pattern recognition receptor,PRR)识别外源病原微生物,然后将其清除。生命信息识别针对危险因素而存在,它是生物历史演化的结果,对每一个生命个体来说,是保障生命安全存在的第一要素。
3 生命信息中危险因素的属性与种类
所谓危险因素一定是与生命相关联的物质,它具有生命信息属性和生命安全性属性。它可以是遗传信息本身,如DNA、RNA,或者是遗传信息产物,如蛋白质、肽,或者是遗传信息产物指导下的合成产物及修饰产物,如细菌的肽聚糖、脂多糖以及脂蛋白、糖蛋白等。一个物质之所以能触发生命机体信息安全控制系统,这个物质必定需要具有上述两个属性,缺一不可。例如半抗原没有免疫原性,是因为半抗原或者缺乏生命信息属性、或者缺乏生命安全性属性,即不具备对机体的危险性。正常菌群虽然具有生命信息属性,但是缺乏生命安全性属性,常规情况下不具有对宿主的危险性属性,故不会引起应答反应。
危险因素分为外源性和内源性。前者来源于病毒、细菌、寄生虫。移植物、血型及变应原等也属于外源性。后者有HSPs、线粒体、透明质酸寡糖、硫酸乙酰肝素多糖片段、纤维蛋白原、纤连蛋白III型重复外结构A、膜表面活性蛋白A、β防御素、高迁移率蛋白组1(HMGB1)等[3]。
4 生命信息识别感应器的形成是保障生命安全控制的硬件基础
4.1 生命信息识别感应器的形成
生命信息识别感应器的形成有“固有”和“适应”两种。前者有Toll样受体家族、病毒RNA识别受体家族、DNA识别受体家族、NOD样受体、C型凝集素受体家族、清道夫受体、甲酸基多肽受体、补体受体等。这些识别感应器的形成,可以理解为生物在对生命信息博弈进化中演变产生,是生命历史性的积淀产物,对于一个个体来说,是由“父母”所赐,所以认为是固有的。从历史的长河来看,是物种在亿万年历史的长河中,由物种本身适应环境信息的变化,世代适应积淀,世代通过遗传信息“博弈”的记忆而“固定”下来。后者有TCR、BCR/Ig。TCR的产生,由胸腺T细胞通过双阴性、双阳性和单阳性阶段,即通过一系列的基因重排和选择,其过程发生在胸腺。研究证实TCR存在受体编辑现象[4]。BCR有着与TCR相类似的过程,这些过程发生在骨髓或法氏囊及囊样组织。BCR更重要的是体细胞高频突变,其过程发生在如脾脏、淋巴结的器官组织中。论“适应”,比如受体编辑,体细胞高频突变,是个体对环境生命信息的一种适应性应答,但是发生在胸腺或者骨髓、法氏囊的过程却与外界环境信息毫无直接关联,可以说,也是由“父母”所赐,本应认作“固有”。
4.2 生命信息识别感应器的种类与特性
4.2.1 PRR PRR识别PAMP,包括细菌脂多糖、脂蛋白、肽聚糖、鞭毛蛋白、非甲基化CpG DNA、病毒双链RNA、真菌细胞壁等。PRR识别PAMP后引发受体配体反应,然后向细胞内传递微生物感染信号,激发机体的应答,从而将病原微生物清除。根据PRR存在的形式可分为膜型、分泌型和胞质型3种形式。膜型,包括TLR家族中多数成员,甘露糖受体(MR)和清道夫受体(SR);分泌型,包括甘露糖结合凝集素(MBL)、C反应蛋白(CRP)和脂多糖结合蛋白(LBP)等;胞质型,包括TLR家族中TLR3、TLR7/TLR8和TLR9,以及NLR家族中所有的成员。
4.2.2 识别感应器总体归纳 由存在的方式,如体液型的有C3b、Ig、凝集素等;细胞膜表面存在的有:BCR、TCR、TLR1/2/4/5/6以及C型凝集素受体家族、清道夫受体、整合素;细胞器存在的有:识别 DNA 受体家族(DAI、AIM2)、识别肽聚糖受体家族(NLRs);在内体及溶酶体中的有:TLR3、7、8、9 及识别RNA受体家族RLRs、PKR等。
感应器识别信息后被激活的定为阳性识别,受到抑制的则为阴性识别。阳性识别又分为信息经分解分析后再进行识别以及信息无需经过分解就能直接进行识别的2种情况。总体情况归纳总结如图1。
TCR识别的命题,一直受到关注。MHC分子给T细胞提供的是一个短肽,为线性多肽。人们会联想BCR识别模式,会从抗原与抗体的关系推测TCR与MHC-多肽的关系。抗原抗体的结合是以空间构象互补关系所决定。从线性多肽出发,其应视为一段密码符号,在本质上是遗传信息的线性符号,由此可见,TCR本质上是识别一个遗传信息(密码),而不是与BCR一样纯粹识别一个空间构象。MHC分子代表自身遗传信息产物,由自身遗传信息产物MHC分子给予T细胞,即给予TCR提呈一个外源多肽遗传信息,其传递的模式如图2所示。

图1 生命信息安全控制识别感受器的方式、种类与定位Fig.1 The mode,category and range of identification sensors of life information security control
由图2可以说明MHC限制现象,即TCR只能接受来自自身遗传信息提供给予的外源遗传信息,换句话说,非自身遗传信息递给的情况下,是不能接受任何一种信息的。K Christopher Garcia等[5]研究,认为TCR与MHC之间的结合存在遗传特异性,并提出了密码子假说(codon hypothesis)。从TCR结合过程可见[6],常规 αβTCR 采用斜线对接模式,结合MHC分子的部位覆盖整个凹槽,其CDR环是可塑性的,采用诱导契合型结合方式,其CDR环在结合前后会有大的构象变化。研究表明,TCR对pMHC识别主要集中于多肽的中部。据此能够说明,BCR与TCR识别方式有着本质不同,前者是刚性构象式识别,比喻为锁钥式识别,属性为空间构象信息;后者是柔性线性式识别,比喻为印章式识别,即印章边框比作MHC,印章中的文形比作短肽信息,其属性为遗传密码信息。前者需要通过体细胞高频突变,从而选择亲和性高的克隆用以严密调整空间构象,以利更精细地适合抗原的构象;后者不需要体细胞高频突变这个环节,因为TCR识别的是一段遗传信息的“文字密码”,因此不需要特地调整构象。诚然,遗传信息的感知同样离不开构象信息传递的方式,比喻为印章,同样存在构象的情景,但它不是刚性,它具有一定弹性和柔性。

图2 MHC-多肽与TCR之间的作用机制Fig.2 The action mechanism between MHC-polypeptide and TCR
5 生命信息感知与处理过程是保障生命安全控制不可或缺的事件
生命信息感知与处理过程发端于由识别到感知,由分析到传递,进而引发细胞的活化、分化、增殖,并产生效应。
APCs通过其所携带的识别感应器感知内源性或外源性危险信号,由此APCs被激活。激活后经由溶酶体酶分析方式或蛋白酶体分析方式对信息进行分析加工,并将加工好的遗传信息通过MHC分子-短肽的形式传递给T细胞,其结果在APCs与T细胞接触之间形成“免疫突触”,即信息簇集的方式进行信息传递,其分子表现形式为MHC-短肽-TCR,同时由共刺激作用以及细胞因子给T细胞发出信号,从而激活T细胞内信号传递,这样T细胞被激活成为细胞毒性T细胞(CTL)或辅助性T细胞(Th)。如上过程概括为,由信息的识别导致信号的产生,信息分子传递、转化生成信号分子,T细胞一方面识别并接受由APCs传递的遗传信息,同时接受共刺激和细胞因子的信号,在这里TCR是信息受体,更确切地说,是遗传信息的受体,除此之外的属于信号受体,诚然信号是信息的载体[7]。
“免疫记忆”是生命信息安全控制中的重要事件。所谓“免疫记忆”,在本质上是对遗传信息的记忆,离开了遗传信息的识别与分析就不存在“免疫记忆”。对于B细胞来说,B细胞所表现出来的记忆,只有在T细胞的辅助下才能发生,也就是说,只有当T细胞产生了对遗传信息记忆的条件下,才有B细胞对构象信息的记忆。通常所说的,胸腺依赖性抗原具有记忆性,而非胸腺依赖性抗原没有记忆性,就是这个道理。
NK细胞对信息的分析与识别感受机制取决于抑制性信号与活化性信号之间的平衡状态,NK细胞表面抑制性受体感受靶细胞表面MHC I类分子后,传导抑制性信号,从而抑制NK细胞的活化,靶细胞免受攻击。一旦活化性信号强势抑制性信号,导致诱导NK细胞的活化。这样NK细胞是对信息分析识别的权衡确定其抑制或活化状态,由此保障其以最大限能力来区分正常细胞和异常细胞,从而确定是否对靶细胞行使“killing”效应。
6 生命信息安全策略行为取决于对生命信息识别分析处理的结果
概括地说,信息、信号间的相互作用与相互制约决定细胞的命运和效应功能。信息与信号的有无、多寡影响应答行为策略的结局。
初始T细胞(naive T cells)富集于脾脏、淋巴结,经由血液循环和淋巴循环行使监视功能。一旦机体遭遇危险因子,非成熟DC通过 PRR识别感受PAMP,在感染部位发挥吞噬作用,并迁移至淋巴结,通过MHC分子提呈短肽,为T细胞提供遗传信息,表达共刺激分子,如CD40、CD80和CD86,分别与T细胞之CD154、CD28和CD152相互作用,为T细胞活化提供信号,并产生细胞因子和趋化因子,为T细胞提供信号,通过上述信息、信号作用于T细胞,即与CD4+T细胞或CD8+T细胞相互作用,共同作用决定T细胞的分化和功能。
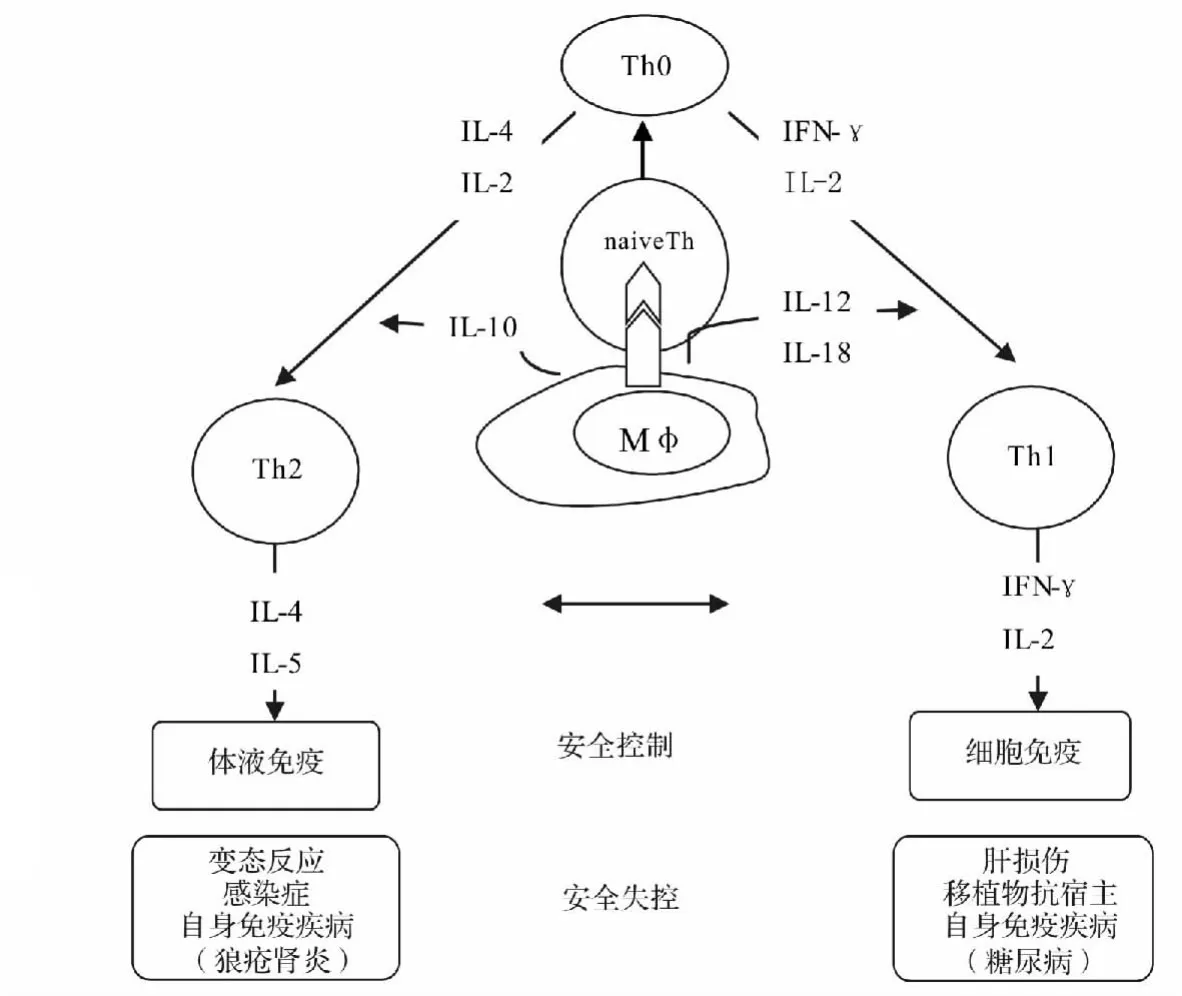
图3 生命信息安全控制中Th1/Th2的平衡与失控Fig.3 The balance and imbalance of Th1/Th2 in life information security control
当缺乏TCR遗传信息,T细胞将不会反应;只有TCR遗传信息而缺乏共刺激信号,T细胞将进入无能状态(anergy)、耐受甚至凋亡[8]。由于DC呈递短肽(遗传信息)的浓度以及表达共刺激分子和分泌细胞因子的不同,决定着T细胞尔后的命运结果,这样初始CD4+T细胞就可能分化成不同的辅助性T细胞,即Th1、Th2、Th17和Threg细胞。不同的Th有着相异的生命功能,从而发挥不同的效应,形成不同的应答结局。Th1负责辅助“细胞免疫”方面的效应,Th2负责辅助“体液免疫”方面的效应。由图3可见[9],不同的细胞因子作为信号所产生的作用效应决定Th细胞的不同取向和功能。

表1 TCR-肽-MHC相互作用改变促使CD4+T细胞变型转变Tab.1 The inter-action in TCR-peptide-MHC induce transformation of the T cell CD4+
由表1可见[10],TCR配体上肽成分序列的改变,这种序列的改变将决定Th1和Th2细胞性质的改变。其机制是对MHC II类分子具有高亲和力的短肽,能诱导细胞向Th1细胞亚类转变,而改变的肽配体(altered peptide ligand,APL)将改变应答发生的方向。研究表明,TCR-肽-MHC三者间的相互作用不仅控制功能性应答的幅度,而且决定应答发生的方向。
7 生命信息安全控制行为过程
NK细胞在杀伤靶细胞过程,通过穿孔素和粒酶系统介导靶细胞坏死及膜TNF家族分子介导靶细胞凋亡。前者需要NK细胞识别受体与靶细胞直接接触方可实现,后者则不需要。NK细胞抗肿瘤,通过“内识别”,如NCRs、NKG2D等直接识别恶性转化的癌细胞并被活化,也可能由辅助细胞,如巨噬细胞、DC的作用下被活化。后者通过PRR,如TLR2、RIG-1、TLR7等进行识别,通过分泌多种细胞因子或直接接触将信号传递到NK细胞,使NK细胞发挥杀伤作用及分泌炎性细胞因子功能。
CTL也是通过穿孔素和粒酶系统介导靶细胞程序性坏死,并通过其FasL与受感染细胞上的Fas相互作用,以这样两种机制导致靶细胞凋亡。受病毒感染的细胞将病毒蛋白的肽经蛋白酶体分析方式,由MHC I类分子呈递信息到细胞表面,这样该细胞便成为CTL杀伤的靶,CTL表达与MHC I类的非多态区结合的CD8,以限制CTL只识别在MHC I类分子中呈递肽的细胞,以稳定TCR与MHC-肽的相互作用。另外表面共刺激分子与黏附分子,如LFA-1对CTL与靶细胞的相互作用和对激活其细胞毒机理发挥重要作用。活化步骤还诱导CTL的FasL表达。CTL含有大量溶解细胞的颗粒,其内含有蛋白酶、粒酶A、粒酶B和穿孔素。颗粒向靶细胞接触点附近的膜移动。在与膜融合时,颗粒释放穿孔素,穿孔素聚合在靶细胞的膜中,造成允许蛋白酶进入的孔。由于酶裂解作用,其产物诱导细胞凋亡。
8 生命信息安全控制错位、缺失、偏离及其危害
由图3可见,当Th1及Th2过度偏离时有可能引发“免疫性疾病”,如Th1型的有肝损伤、糖尿病、移植物抗宿主反应。Th2型的有呼吸道过敏反应、特应性皮肤炎。也就是说,当Th1/Th2平衡的失控将导致系统的紊乱,发生对机体安全的危害。
MHC II类分子缺陷又称裸淋巴细胞综合症(bare lymphocyte syndrome,BLS),以MHC II类分子表达缺失为特征。由于MHC II类分子缺陷导致CD4+T细胞不能呈递肽,患者细胞免疫和体液免疫出现严重缺陷,临床表现为许多机会性致病菌感染[4]。
IL-2如图3所示,对Th细胞的分化发挥重要作用,也能对B细胞、NK细胞、单核细胞及巨噬细胞产生重要作用,是因为这些细胞表面存在IL-2的受体。IL-2受体由三个亚基,即α、β、γ链组成。γ链基因位于X染色体上,由于γ链基因发生变异,导致γ链功能异常。研究表明,γ链功能异常是引起X-连锁严重联合免疫缺陷病(X-linked severe combined immune deficiency,XSCID)的原因[11]。XSCID患者体液免疫和细胞免疫都缺陷,其外周血T细胞显著减少或缺失。胸腺几乎不见发育,T细胞早期分化异常。B细胞表面只表达IgM,其它类别未发现。这一点是由于B细胞的类别转换过程中缺失T细胞提供的必需信号的缘故。γ链属于细胞因子受体,持续表达在T细胞,B细胞,NK细胞,髓样细胞和成红细胞上。γ链与IL-2受体α链和β链共同组成高亲和力IL-2受体,参与IL-2的功能。酪氨酸激酶Jak-3主要功能是传导γ链的信号,Jak-3缺陷导致的表型与XLSCID相同。γ链蛋白细胞外功能区单个氨基酸置换,阻断了T细胞和NK细胞的分化。临床表现为口腔念珠菌病,红斑,持续性腹泻,呼吸道合胞病毒、副流感病毒3、腺病毒、卡氏肺囊虫感染以及革兰阴性菌败血症。
[1]Matzinger,P.tolerance,danger and the extended family[J].An Reviews of Immunology,1994,12:991-1045.
[2]Stan J,Brouns J.A swiss army knife of immunity[J].Science,2012,337:808-809.
[3]庞广昌.食品免疫论[M].北京:科学出版社,2008:205.
[4]曹雪涛.免疫学前沿进展[M].北京:人民卫生出版社,2009:252-259,472.
[5]Feng D,Bond C J,Ely L K,et al.Structural evidence for a germline-encoded T cell receptor-major histocompatibility complex interaction’codon?[J].Nat Immunol,2007,8(9):975-983.
[6]Borg N A,Wun K S,Kjer-Nielsen L,et al.CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor[J].Nature,2007,448(7149):44-49.
[7]唐晓波.信息安全概论[M].北京:科学出版社,2010.
[8]舒红兵.抗病毒天然免疫[M].北京:科学出版社,2009:198.
[9]齊藤 隆,竹森利忠.新用語ライブラリ— 免疫[M].2版.東京都:羊土社,2000:58.
[10]余传霖,熊思东.分子免疫学[M].上海:复旦大学出版社,上海医科大学出版社,2001:135.
[11]中村正孝,菅村和夫.インターロイキン2(IL-2)[J].Clinical Immunology,1995,(Suppl.16):29-38.
