长吻养殖群体与野生群体遗传多样性分析
2013-07-23肖明松马玉涵
肖明松 崔 峰 康 健 马玉涵
(安徽科技学院生命科学学院, 凤阳 233100)
1 材料与方法
1.1 材料
主要试剂 蛋白酶K、琼脂糖、饱和酚购自上海生工生物工程技术有限公司, 引物DL1 (5′-AC CCCTGGCTCCCAAAGC-3′) 和 DH2 (5′-ATCTTAG CATCTTCAGTG-3′)[18]由上海生工生物工程技术有限公司合成,Taq酶、dNTPs 购自大连TaKaRa公司;其他试剂均为国产分析纯。
主要仪器 PCR 仪、核酸分析仪和离心机为Eppendorf (德国), Alphalmager Is-2200凝胶图像分析系统(美国), 电泳仪等常规仪器均为国产。
1.2 方法
模板 DNA的制备 每个样品随机取肌肉100—200 mg, 剪碎, 匀浆后置于1.5 mL 的微量离心管中, 加入含蛋白酶K 的裂解液(10 mmol/ L Tris-HCl, pH 8.0, 1 mmol/ L EDTA, pH 8.0, 1%SDS) ,55℃消化过夜。次日分别用Tris-饱和酚、氯仿/异戊醇(24∶1) 提取、纯化DNA, 之后用−20℃冷冻的无水乙醇沉淀DNA, 自然干燥后, 溶于灭菌双蒸水中,所得用基因组 DNA 样品用紫外分光光度计测量DNA 样品的浓度和纯度, 同时辅以琼脂糖凝胶电泳检测基因组 DNA 的完整性并估测分子量, 置于−20℃冰箱保存备用。
PCR 反应 PCR 反应总体积为 30 μL, 其中含 10×buffer反应缓冲液 3.0 μL, 3 μL dNTP(2.5 mmol/L), 0.4 μmol/L 引物, 0.25 μLTaq酶, 25 ng DNA, 用灭菌双蒸馏水补足体积。PCR反应条件为: 94℃下预变性5min, 94℃下变性30s, 58℃下退火1min, 72℃下延伸1min, 共35个循环, 最后72℃下延伸10min, 4℃保存。
琼脂糖凝胶电泳 PCR 扩增产物在1.2 %的琼脂糖凝胶(含0.5 g/mL EB) 上电泳, 120 V 稳压电泳1h。电泳结果在Alphalmager Is-2200'凝胶图像分析系统中拍照、保存。
DNA序列测定和分析 经2 %琼脂糖凝胶电泳检测后, 将 PCR 产物送往上海生工生物工程技术有限公司进行序列测定, 测得的序列用Clustal X 1.81 程序[19]进行比对, 并辅以人工校对, 进行同源性比较。所有序列均递交 GenBank (序列号为JN131500-JN131517和 AB054127-AY297099)。利用MEGA 3.0 软件包[20]分析序列特征、统计碱基组成和转换与颠换值、计算遗传差异和遗传距离。采用邻接法(Neighbor-joining tree, NJ)构建分子系统发育树, 系统树各分支的置信度由 1000次自举法(Bootstrap)检验。利用DnaSP 4.0软件[21]计算各群体的单倍型多样度(H)、平均核苷酸差异数(K)、核苷酸多态性(Pi); Arlequin 3.1软件[22]进行群体间的欧氏距离平方(Squared euclidean distance) 矩阵分子变异分析(AMOVA), 统计长吻养殖群体与野生群体间的遗传分化指数(Fst), 并根据Nm≈(1−Fst)/(2Fst)得到群体间的基因流值(Nm)。为进行谱系生物地理学(Phylogeography)分析, 利用 TCS 1.21[23]软件, 依据统计简约原理[24], 构建单倍型的网状图。相对于传统的系统发育分析方法, 网状图更能够有效地揭示种群水平上的谱系关系(Genealogical relationship),更清楚地反映与种内基因进化有关的特性, 如是否保持祖先的单倍型、是否存在多重后代的单倍型以及种群间的序列差异较低的特性[25]。
2 结果
2.1 长吻线粒体控制区序列变异、单倍型和单倍型分布
8个, 缺失位点1个。含有4个转换位点(Ts), 1个颠换位点(Tv), 平均转颠换比(Ts/Tv)为8.27。在长吻野生群体中, 69个个体共检测到 35个变异位点(占全部碱基数的4.43%)。其中简约信息位点24个, 单突变位点10个, 缺失位点1个。含有5个转换位点(Ts), 1个颠换位点(Tv), 平均转颠换比(Ts/Tv)为9.80。在长吻养殖群体和野生群体中, 135个个体共检测到42个变异位点, 其中简约信息位点28个,单突变位点12个, 缺失位点2个(表1)。长吻养殖群体和野生群体的碱基变化不大(表2), 其中碱基G的含量显著低于其他碱基的含量, A+T含量(60.6%)明显高于C+G含量(39.4%)。
表1 长吻线粒体控制区单倍型分布及单倍型的变异位点Tab.1Alignment of mtDNAcontrol region haplotypes of variable positions and distribution of mtDNAcontrol region haplotypes of Leiocassis longirostris
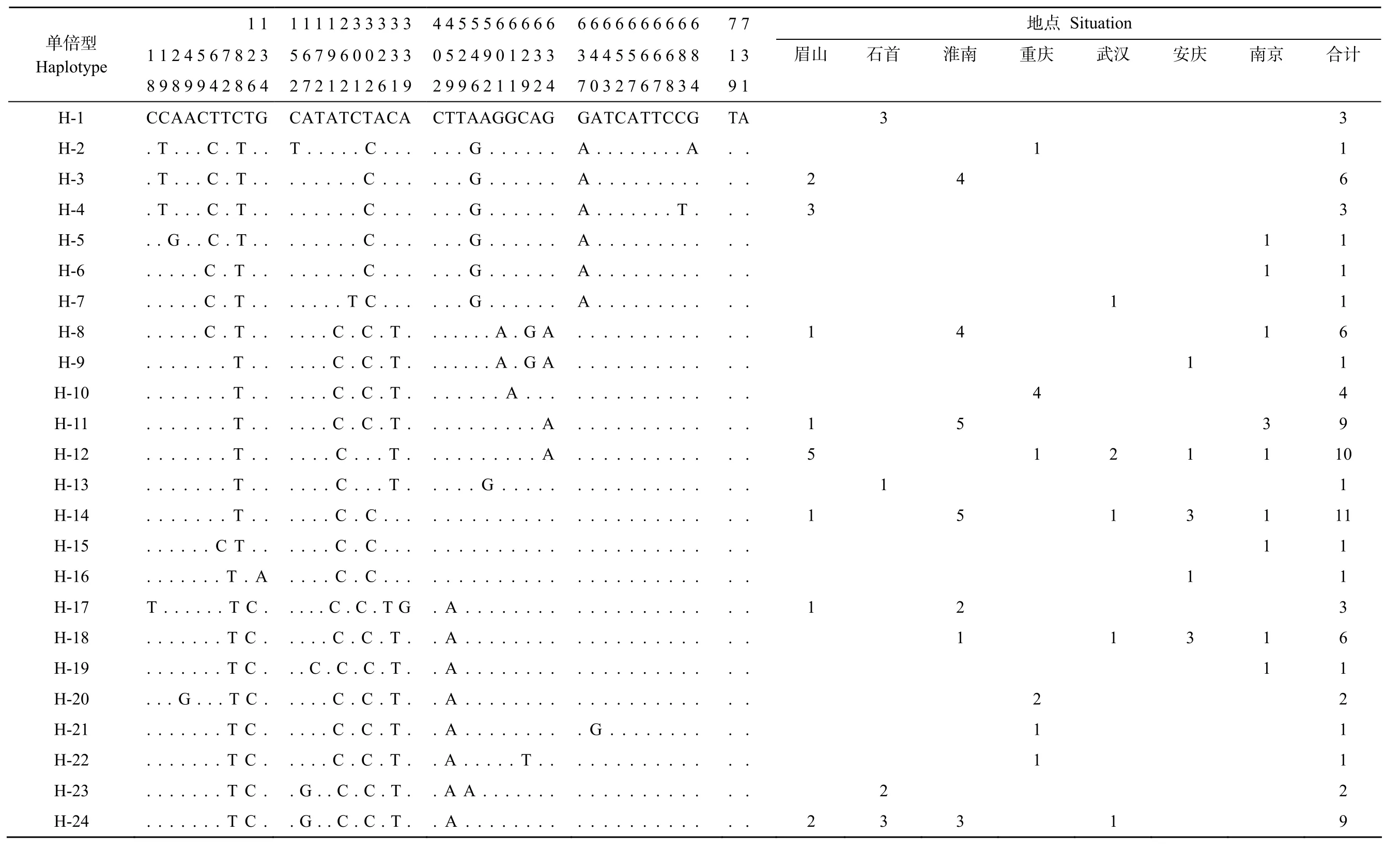
表1 长吻线粒体控制区单倍型分布及单倍型的变异位点Tab.1Alignment of mtDNAcontrol region haplotypes of variable positions and distribution of mtDNAcontrol region haplotypes of Leiocassis longirostris
计合316311161491 011 11136121129京111311111南庆安11313 ation汉武1 2111 Situ点地庆1 41 211重南淮4 455213首3 1 23石山2311511 2眉7 7 1 3 9 1 TA................................................................A........CG.....T................................TC..............................................AT..............................................TC......................................G.......6 6 6 6 6 6 6 6 6 6 3 4 4 5 5 6 6 6 8 8 7 0 3 2 7 6 7 8 3 4 GA.......... A.. A.. A.. A.. A.. A.. ... ... ... ... ... ... ... ... ... .AG G A G A.A.A.. ... ... .GC............A.A.A........................T......AG..4 4 5 5 5 6 6 6 6 6 0 5 2 4 9 0 1 2 3 3 2 9 9 6 2 1 1 9 2 4..............................TA.G.G.G.G.G.G..................G...................A...CT...................................A.A.A.A.A.A.A.A CA.1 1 1 2 3 3 3 3 3 6 7 9 6 0 0 2 3 3 7 2 1 2 1 2 6 1 9................G.......TA...........T.T.T.T.T.T....C..T.T.T.T.T.T.T.T TC.C.C.C.C.C T C..C.C.C.C.....C.C C..C.C.C.C.C.C.C.C.............C.C.C.C.C.C.C.C.C.C.C.C.C.C.C.C TA...................................C......G.G.CA....1 1 1 TG.. T 1 1 2 4 5 6 7 8 2 3 5 8 9 8 9 9 4 2 8 6 4 2.. ... ... ... ... ... ... ... ... ... ... ... ..A C. .C. .C. .C. .C. .C. .C. .C. .CTTC.T.T.T.T.T.C.T.C.T.C.C.T...T...T...C.T...T...T...C C T..T.T...T.T...T...T.T.T.T..AA...C..........CC.........T.T.T G...............................................T............G................e型倍单Haplotyp H-1 H-2 H-3 H-4 H-5 H-6 H-7 H-8 H-9 H-10 H-11 H-12 H-13 H-14 H-15 H-16 H-17 H-18 H-19 H-20 H-21 H-22 H-23 H-24
2.2 单倍型分布及单倍型聚类分析
利用DnaSP 4.0 软件计算各群体的单倍型(表1)。由表1可知, 在长吻不同群体中, 135个体共检测出46 个单倍型, 即H-1 - H-46。其中28个独享单倍型, 占 20.74%,18个共享单倍型, 单倍型 H-8、H-11、H-12、H-14、H-18、H-24、H-37和 H-43为野生群体和养殖群体共享。另外, 在长吻养殖群体中, 66个体共检测出 18 个单倍型, 在野生群体中 69个体共检测出36 个单倍型(表略)。TCS 1.21软件生成的统计简约网状图显示出星状的分布态势,没有将 46个单倍型区分为对应不同地理区域或者地理种群的单系群。单倍型H- 14位于星状图的中心,其他单倍型则与其依短支相连(图1)。依据溯祖理论 (Coalescence theory), 线粒体控制区单倍型的星状拓扑结构表明所研究的种群经历了明显的种群扩张[26], 单倍型 H-14由于其分布最广和享有该单倍型的个体在所有的种群中所占的比重也明显的高于其他单倍型, 而且在网状图中处于基部位置, 说明单倍型 H-14可能是原始的单倍型。
表2 长吻序列碱基组成Tab.2 Nucleotide compositions of mtDNA control region in Leiocassis longirostris

表2 长吻序列碱基组成Tab.2 Nucleotide compositions of mtDNA control region in Leiocassis longirostris
群体 Populations T C A G眉山 Meishan 31.6 25.2 29.1 14.1石首 Shishou 31.5 25.2 29.1 14.2淮南 Huainan 31.5 25.3 29.0 14.2平均Average 31.6 25.2 29.0 14.2重庆 Chongqing 31.5 25.3 29.1 14.1武汉 Wuhan 31.5 25.3 29.1 14.1安庆 Anqing 31.5 25.3 29.0 14.2南京 Nanjing 31.5 25.3 29.1 14.1平均 Average 31.5 25.3 29.1 14.1

图1 基于线粒体控制区单倍型的统计简约网状图Fig.1 The statistical parsimony network inferred from mtDNA control region haplotypes
2.3 群体遗传多态性参数统计
利用 DnaSP 4.0 软件计算各群体的单倍型多样性(Hd)、平均核苷酸差异数(K)、核苷酸多态性(Pi)。由表3可知, 长吻4个野生群体的单倍型多样性Hd值均较高, 而从衡量群体遗传水平的K值和Pi值来看, 重庆野生群体明显高于其他群体。长吻3个养殖群体的平均单倍型多样性和平均核苷酸多样性相对较低 (Hd=0.8867±0.0013,Pi=0.0056±0.0013)。
2.4 群体遗传分化
群体间的序列差异反映的是生物间亲缘关系的远近, 使用MAGA 3.0 软件中Tamura-Nei 模型计算群体内及群体间的遗传距离(表4)。结果表明, 四川眉山和湖北石首人工养殖群体间的亲缘关系较远,而安徽淮南养殖群体与湖北石首和四川眉山人工养殖群体间的亲缘关系较近, 长吻养殖群体与野生群体存在一定的遗传距离。分子变异分析(AMOVA)的结果表明, 长吻群体内存在较高的遗传变异(96.42%), 而群体间的遗传变异较小(3.21%) (表 5),分子遗传变异主要来自地理区内群体内个体间。7个群体两两比较的遗传分化指数(Fst)显示, 安庆群体和石首群体之间的遗传分化最大(Fst=0.1125), 而武汉群体和安庆群体之间的遗传分化最小(Fst=0.0014), 其中长吻养殖群体间的遗传分化高于野生群体间的遗传分化。 此外, 群体间的遗传距离分析与遗传分化指数的分析结果是一致(表 4)。Arlequin 3.1 软件统计养殖群体与野生群体间的遗传分化指数和基因流分别为Fst=0.0358、Nm=13.47。
表3 长吻7个群体遗传多样性统计参数Tab.3 Demographic parameters estimated from seven Leiocassis longirostris populations

表3 长吻7个群体遗传多样性统计参数Tab.3 Demographic parameters estimated from seven Leiocassis longirostris populations
群体 Populations 单倍型多样性 Hd 平均核苷酸差异数 K 核苷酸多样性 Pi眉山 Meishan 0.8972±0.0361 4.8854 0.0064±0.0013石首 Shishou 0.8580±0.0570 4.3500 0.0049±0.0021淮南 Huainan 0.9048±0.0260 4.9444 0.0056±0.0011平均 Average 0.8867±0.0013 4.7266 0.0056±0.0013重庆 Chongqing 0.9470±0.0320 6.5580 0.0083±0.0008武汉 Wuhan 0.9810±0.0310 6.1520 0.0078±0.0007安庆 Anqing 0.9170±0.0420 3.9580 0.0050±0.0006南京 Nanjing 0.9430±0.0540 4.2860 0.0054±0.0007平均 Average 0.9736±0.0070 5.3030 0.0087±0.0015
表4 长吻群体间的遗传分化指数(左下角)和遗传距离(右上角)Tab.4 Fixation Index (F) (below) and genetic distance (above) in three populations of Leiocassis longirostris
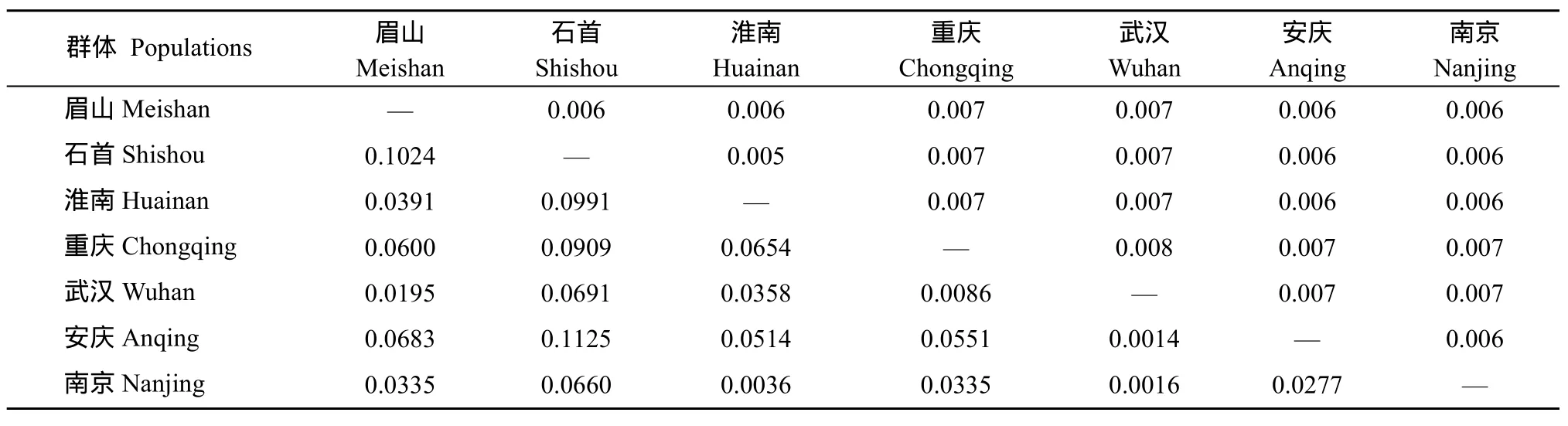
表4 长吻群体间的遗传分化指数(左下角)和遗传距离(右上角)Tab.4 Fixation Index (F) (below) and genetic distance (above) in three populations of Leiocassis longirostris
群体 Populations 眉山Meishan石首Shishou淮南Huainan重庆Chongqing武汉Wuhan安庆Anqing南京Nanjing眉山Meishan — 0.006 0.006 0.007 0.007 0.006 0.006石首Shishou 0.1024 — 0.005 0.007 0.007 0.006 0.006淮南Huainan 0.0391 0.0991 — 0.007 0.007 0.006 0.006重庆Chongqing 0.0600 0.0909 0.0654 — 0.008 0.007 0.007武汉Wuhan 0.0195 0.0691 0.0358 0.0086 — 0.007 0.007安庆Anqing 0.0683 0.1125 0.0514 0.0551 0.0014 — 0.006南京Nanjing 0.0335 0.0660 0.0036 0.0335 0.0016 0.0277 —

表5 长吻?群体分子变异分析结果Tab.5 Analysis of molecular variance (AMOVA) for the Leiocassis longirostris populations
2.5 群体间的亲缘关系及聚类分析

图2 长吻各单倍型基于线粒体控制区序列构建的NJ树Fig.2 NJ tree based on the control region of 7 populations of Leiocassis longirostris
3 讨论
3.1 长吻群体的遗传多样性
遗传多样性的研究是生物多样性研究的重要内容, 只有通过遗传多样性的研究才能从本质上揭示物种多样性的起源、变异和进化。核苷酸多态性(Pi)作为一个衡量群体间遗传多态性的重要指标, 表示各种 mtDNA 单倍型在群体中所占的比例。由实验结果可知, 在长吻养殖群体中66个个体线粒体控制区790 bp的序列中, 总核苷酸突变位点为27个,共检测出18个单倍型。长吻野生群体中69个个体共检测到35个变异位点, 36个单倍型。与长吻野生群体相比, 养殖群体的单倍型多样性和核苷酸多样性水平均较低(Hd=0.8867±0.0013,Pi=0.0057±0.0013)。另外, 从 3个养殖群体内部的核苷酸位点突变情况来看, 安徽淮南群体内有22个核苷酸位点的变异, 湖北石首群体共产生12个变异位点, 四川眉山群体有17个变异位点, 分别只占总群体变异的81.48%、44.44%和62.96%。这表明遗传变异主要存在于群体内, 群体间的基因变异较小。安徽淮南群体的基因多样性水平最高, 石首和眉山河群体较低。Wang,et al.[27]对取自长江上游和下游的野生长吻进行 PCR–RFLP分析, 发现野生长吻遗传多样性低, 种群数量下降, 这与已有野生种群的研究相一致[28,29]。而Yang,et al.[30]利用线粒体控制序列和核SSR标记研究长江上游、中游和下游野生长吻遗传变异和种群结构, 表明长江野生长吻单倍型多样性较高, 核苷酸多样性水平较低(Hd=0.9770±0.0041,Pi=0.0081±0.0043)。上述研究结果表明, 长吻长期的人工养殖已使得该群体的遗传多样性水平明显降低, 反映了长吻作为我国鲶形目分布最广且产量较高的特产淡水名贵经济鱼类特殊的生命进化历程和适应复杂环境的能力。造成这一现象的主要原因可能是长吻的基础群体数量较小、养殖过程的近交及遗传漂变等。而相对较高Hd值、低Pi值表明长吻这个群体可能是由一个较小的有效群体迅速增长, 尽管变异导致单倍型的多态性的积累, 但核苷酸序列的多样化还未能积累[31]。高的Hd为通过分子选育等技术有效防止品种退化提供了前提条件, 并为保持遗传多样性水平, 维持杂交选育优势提供了重要基础。这也在一定程度上说明定期补充不同地区不同群体的个体作为亲鱼进行繁育有利于保持长吻人工繁殖群体的遗传多样性, 避免近亲繁殖和瓶颈效应。
3.2 长吻群体的遗传结构分析
[1] He X F, Su L D, Zhou G R,et al.Study on domestication and reproduction test ofLeiocassis longirostris[J].Freshwater Fisheries, 1985, (1): 14—17 [何学福, 苏良栋, 周贵荣, 等.长吻的蓄养繁殖试验研究.淡水渔业, 1985, (1): 14—17]
[2] Fang J Q.Collection and domestication ofLeiocassis longirostrisfrom the Yangtze River [J].Scientific Fish Farming,1999, (7): 19 [方建清.长江长吻的收集与蓄养.科学养鱼, 1999, (7): 19]
[3] Liu J K, He B W.The Chinese Freshwater Fisheries Science(third edition) [M].Beijing: Science and Technology Press.1992, 304 [刘建康, 何碧梧.中国淡水鱼类养殖学(第三版).北京: 科学技术出版社.1992, 304]
[4] Gong J H.Test on cage culture wildLeiocassis longirostrisin reservoir [J].Reservoir Fisheries, 2006, 26(1): 54 [龚建辉.水库网箱养殖野生长吻试验.水利渔业, 2006, 26(1):54]
[5] Wu Q J.Population ecology ofLeiocassis longirostris(Gunther) (Pisces, Bagridae) with reference to the problem of maximum sustained yield [J].Acta Hydrobiologica Sinica,1975, 5(3): 387—408 [吴清江.长吻种群生态学及其最大持续渔获量的研究.水生生物学集刊, 1975, 5(3): 387—408]
[6] Xiao M S, Wan Q.The biology and culture prospect ofLeiocassis longirostrisin the Yangtze River [J].Journal of Anhui Technical Teachers College, 2001, 15(3): 49—51 [肖明松, 万全.长江长吻的生物学习性及其发展前景.安徽技术师范学院学报, 2001, 15(3): 49—51]
[7] Zhang Y Y.Researches and development ofLeiocassis longirostrisGunther [J].Southwest China Journal of Agricultural Sciences, 1998, S1(11): 134—139 [张义云.长吻(Leiocassis longirostrisGunther) 的研究与开发.西南农业学报, 1998, S1(11): 134—139]
[8] Mo Y X, Wang X Q, Mo Y L.Morphological and histological observations of digestive system inLeiocassis longirostris[J].Journal of Hunan Agricultural University(Natu-ral Sciences), 2004, 30(3): 267—271 [莫艳秀, 王晓清, 莫永亮.长吻消化系统的形态学与组织学观察.湖南农业大学学报(自然科学版), 2004, 30(3): 267—271]
[9] Luo M, Jiang L K, Liu Y,et al.Comparative study on isoenzymes inLeiocassis longirostris[J].Chinese Journal of Applied & Environmental Biology, 2000, 6(5): 447—451[罗曼, 蒋立科, 刘颖, 等.野生与养殖长吻血液及不同器官同工酶的比较.应用与环境生物学报, 2000, 6(5):447—451]
[10] Wan Q, Liu E S, Shen D L,et al.Analysis on karyotype ofLeiocassis longirostrisGunther [J].Journal of Anhui Agricultural University, 2002, 29(2): 182—184 [万全, 刘恩生,申德林, 等.长吻染色体组型分析.安徽农业大学学报,2002, 29(2): 182—184]
[11] Xiao M S, Yang G.Isolation and characterization of 17 microsatellite loci for the Chinese longsnout catfishLeiocassis longirostris[J].Molecular Ecology Resources, 2009, 9(3):1039—1041
[12] Yu Y Y, Xiao M S, Chen L,et al.Isolation and characterization of microsatellite loci in the longsnout catfish (Leiocassis longirostris) [J].Aquaculture Research, 2009, 40(2):246—248
[13] Bermingham E, Avise J C.Molecular zoogeography of freshwater fishes in the southeastern United States [J].Genetics, 1986, 113(4): 939—965
[14] Grunwald C, Stabile J, Waldman J R,et al.Population genetics of shortnose sturgeonAcipenser brevirostrumbased on mitochondrial DNA control region sequences [J].Molecular Ecology, 2002, 11(10): 1885—1898
[15] Mabuchi K, Senou H, Suzuki T,et al.Discovery of an ancient lineage ofCyprinus carpiofrom Lake Biwa, central Japan, based on mtDNA sequence data, with reference to possible multiple origins of koi [J].Journal of Fish Biology,2005, 66(6): 1516—1528
[16] Kyle C J, Wilson C C.Mitochondrial DNA identification of game and harvested freshwater fish species [J].Forensic Science International, 2007, 166(1): 68—76
[17] Teletchea F.Molecular identification methods of fish species:reassessment and possible applications [J].Reviews in Fish Biology and Fisheries, 2009, 19: 265—293
[18] Zhang Y, Zhang E, He S P.Studies on the structure of the control region of the bagridae in China and its phylogentic significance [J].Acta Hydrobiologica Sinica, 2003, 27(5):463—467 [张燕, 张鹗, 何舜平.鲿中国科鱼类线粒体DNA控制区结构及其系统发育分析.水生生物学报, 2003,27(5): 463—467]
[19] Thompson J D, Gibson T J, Plewniak F,et al.The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools [J].Nucleic Acids Research, 1997, 24: 4876—4882
[20] Kumar S, Tamura K, Nei M.Mega 3.0: Integrated software for molecular evolutionary genetics analysis and sequence莫永亮.长吻遗传多样性的RAPD分析.江西水产科技,2010, 122(2): 13—16]
[29] Wang H Y, Huang W Q.Preliminary analysis on the genetic diversity in four populations ofLeiocassis longirostrisby using microsatellite markers [J].Journal of Henan Agricultural Sciences, 2011, 40(2): 146—148 [王红莹, 黄文清.应用微卫星标记分析长江流域长吻4 个群体的遗传多样性.河南农业科学, 2011, 40(2): 146—148]
[30] Yang G, Xiao M S, Yu Y Y,et al.Genetic variation at mtDNA and microsatellite loci in Chinese longsnout catfish(Leiocassis longirostris) [J].Molecular Biology Reports,2012, 39(4): 4605—4617
[31] Avise J C.Phylogeography the History and Formation of Species [M].Cambridge, Massachusetts London, England:Harvard University Press.2000, 213—285
[32] Meng W, Guo Y, Hai S,et al.Genetic structure and diversity ofSchizothorax biddulphipopulations [J].Acta Hydrobiologica Sinica, 2012, 36(5): 851—857 [孟玮, 郭焱, 海萨,等.塔里木裂腹鱼群体遗传结构及遗传多样性分析.水生生物学报, 2012, 36(5): 851—857]
[33] Masatoshi N.Molecular Population Genetics and Evolution[M].Beijing: Agriculture Press.1975, 121—133 [根井正alignment [J].Briefings in Bioinformatics, 2004, 5: 150—163
[21] Rozas J, Sánchez-del Barrio J C, Messeguer X,et al.DNA polymorphism analyses by the coalescent and other methods[J].Bioinformatics, 2003, 19: 2496—2497
[22] Excoffier L, Laval G, Schneider S.Arlequin ver 3.0.An integrated software package for population genetics data analysis[J].Evolutionary Bioinformatics Online, 2005, 1: 47—50
[23] Clement M, Posada D, Crandall K A.TCS: a computer program to estimate gene genealogies [J].Molecular Ecology,2009, 9: 1657—1660
[24] Templeton A K.Crandall K A, Sing C F.A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data.III.Cladogram estimation [J].Genetics, 1992, 132: 619—633
[25] Posada D, Crandall K A.Evaluation of methods for detecting recombination from DNA sequences: Computer simulations[J].Proceedings of the National Academyof Sciences of the United States of America, 2001, 98: 13757—13762
[26] Slatkin M, Hudsun R R.Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations [J].Genetics, 1991, 129: 555—562
[27] Wang Z W, Zhou J F, Ye Y,et al.Genetic structure and lowgenetic diversity suggesting the necessity for conservation of the Chinese longsnout catfishLeiocassis longirostrisGünther(Pisces: Bagriidae) [J].Environmental Biology of Fishes,2006, 75: 455—463
[28] Mo Y X, Wang X Q, Mo Y L.RAPD analysis of genetic diversity ofLeiocassis longirostris[J].Journal of Jiangxi Fisheries Science, 2010, 122(2): 13—16 [莫艳秀, 王晓清,利.分子群体遗传学与进化论.北京: 农业出版社.1975,121—133]
[34] Wright S.The gentical structure of population [J].Annals of Eugenics, 1951, 15: 323—334
[35] Wright S.Evolution in Mendelian populations [J].Genetics,1931, 16: 97—159
[36] Han X L, Xu J R, Li X R,et al.Analysis of genetic diversity ofElopichthys bambusaby AFLP Markers [J].Journal of Nan jing Normal University(Natural Science Edition), 2009,32(1): 110—114 [韩晓磊, 徐建荣, 李小蕊, 等.鳡鱼群体遗传多样性的 AFLP 分析.南京师大学报(自然科学版),2009, 32(1): 110—114]
[37] Yang X, Yang J F, Tang M L,et al.Intraspecific genetic polymorphisms ofSiniperca ScherzeriSteindacher and molecular identification withSiniperca chuatis[J].Acta Hydrobiologica Sinica, 2007, 31(6): 891—895 [杨星, 杨军峰, 汤明亮, 等.斑鳜种内遗传多态性以及与翘嘴鳜的分子鉴别.水生生物学报, 2007, 31(6): 891—895]
