Typha × glauca Godr., 香蒲属(香蒲科)中国新记录杂种及其形态特征
2013-04-19朱秀玉
朱秀玉 王 东
(华中师范大学生命科学学院, 武汉 430079)
Typha × glauca Godr., 香蒲属(香蒲科)中国新记录杂种及其形态特征
朱秀玉 王 东
(华中师范大学生命科学学院, 武汉 430079)
香蒲属(Typha L.)为多年生水生或沼生草本植物, 种间存在十分普遍的杂交现象, 其中一些杂种在湿地生态系统中有重要的作用。在查阅大量腊叶标本基础上, 结合野外居群生物学工作, 作者发现中国一新记录杂种, 即T. × glauca Godr. (T. angustifolia L. × T. latifolia L.), 并新拟“粉绿香蒲”作该杂种的中文名。对粉绿香蒲的形态特征进行了研究, 讨论了其与亲本水烛和宽叶香蒲的区别, 并给出检索表。
香蒲属; 水烛; 宽叶香蒲; 粉绿香蒲; 新记录
香蒲属(Typha L.)隶属香蒲科(Typhaceae), 世界约24种[1—4], 分布于南北两半球的温带和热带地区。我国约有12种, 南北广泛分布[5,6]。该属种间存在十分普遍的杂交现象, 欧洲、美洲等地已经报道有7个杂种[7—13], 其中T. angustifolia × T. latifolia (= T. × glauca Godr.)、T. angustifolia × T. domingensis和T. domingensis × T. latifolia (= T. × provincialis Camus)[8]等3个杂种经过细胞遗传学实验确认。在欧洲和北美等地, 香蒲属杂种是湿地尤其是在严重人为干扰或易受较大水位波动影响生境中的优势植物, 它们在湿地生态系统中的作用一直受到植物学家和生态学家的关注[9—11,14—19]。
我们在研究香蒲属标本过程中, 发现采自新疆西部的10份标本, 其形态特征属于杂种T. × glauca Godr.的范畴。该杂种以前在国内未见报道, 属于中国的一分布新记录, 本文予以报道。
1 材料与方法
1.1 腊叶标本研究
研究存放中国科学院植物研究所植物标本馆(PE)、中国科学院华南植物园植物标本馆(IBSC)、中国科学院成都生物研究所植物标本馆(CDBI)、中国科学院武汉植物园植物标本馆(HIB)、武汉大学植物标本馆(WH)、四川大学植物标本馆(SZ)等所有馆藏香蒲标本以及我们野外采集的香蒲标本[存放华中师范大学植物标本馆(CCNU)], 尤其是馆藏标本中被鉴定为水烛(T. angustifolia L.)和宽叶香蒲(T. latifolia L.)的标本。形态特征对比研究包括外部形态、花、果实、种子及花粉等。
1.2 形态观察和测量
用直接观察法研究植物的营养器官和繁殖器官的形态特征。在NIKON SMZ-1500体视显微镜下观察花、果实和种子, 在OLYMPUS BX51型光学显微镜下观察花粉粒, 并分别测量大小和拍照。取腊叶标本的花药, 并经冰醋酸浸泡、醋酸酐处理、离心沉淀淘洗后制作装片, 用于花粉粒的观察和大小测量。选取10个以上样本用于各项数据在显微镜下性状的测量, 求平均值。
2 分类与形态学描述
粉绿香蒲(杂种) 新拟 图版Ⅰ: 6—11, 20
Typha × glauca Godr. (T. angustifolia L. ×T.latifolia L.). Fl. Lorr. 3: 20, 1844.
Synonymy: T. latifolia var. elongata Dudley in Bul. Cornell Univ. (Sci.) 2: 102, 1886; T. angustifolia var. elongata (Dudley) Wiegand in Rhodora 26: 1, 1924.
多年生水生或沼生草本, 雌雄同株。地上茎直立, 粗壮, 高约2—3 m。叶条形, 粉绿色, 叶片长73—140 cm, 宽10—16 mm; 叶鞘耳状抱茎。雌雄花序分离或紧密连接, 相距(0—)1—12 mm; 雄花序长25—40 mm, 雄花由1枚或2枚雄蕊组成, 花药长约2—3 mm, 长矩圆形, 花粉粒单体、二合体、三合体或四合体; 雌花序长26—43 cm, 宽19—22 mm,深棕色; 雌花具小苞片, 小苞片长约7.6 mm, 浅褐色至先端白色, 比柱头窄; 孕性雌花柱头长1.2—1.6 mm, 宽0.1—0.2 mm, 条状披针形, 子房狭披针形, 长约1.2 mm; 不孕雌花子房长0.9—1.5 mm, 近倒圆锥形, 先端圆形。花果期6—9月。
China. Xinjiang(新疆): Huocheng(霍城), 生于积水塘、人工沟渠中。2010–8–13, D. Wang(王东) 6248, 6249(CCNU); Tacheng(塔城), 生于水塘中。2010–8–9, D. Wang(王东)6223–1(CCNU)。中国分布新记录。欧洲(英国、法国)、北美(美国、加拿大)等地也有分布。
3 讨论
粉绿香蒲最早是由法国学者D. A. Godron于1844年在法国东北部Lorraine地区植物志Flore de Lorraine中命名的一种植物。作者在原描述中指出该植物在外部形态上与宽叶香蒲相近, 与水烛有明显区别。1886年美国学者W. R. Dudley在美国纽约州Cayuga湖泊发现一种与宽叶香蒲形态相似的植物, 但因其雌花序长达30 cm, 他把这种植物定名为T. latifolia var. elongata, 1924年美国学者K. M. Wiegand在复查Cayuga湖泊水生植物时, 根据T. latifolia var. elongata在外部形态上更接近于水烛而将其命名为T. angustifolia var. elongata, 这两个变种名称后来被证明都属于粉绿香蒲[7]。除少数北美学者认为该分类群应作为种来处理外[7,20], 多数欧美学者根据来自形态学、杂交、血清学、电泳、酶和分子等方面的研究证据, 都支持它是杂种的观点[8,12,13,21—24]。据文献记载, 粉绿香蒲广泛分布于欧洲、北美等地[7,9,11—13,15,16], 如它在北美东北部大西洋沿岸到中部广阔的草原地带不但分布较广, 而且数量多[9,11]。种加词“glauca”意指叶片粉绿色, 因此本文新拟“粉绿香蒲”作该杂种的中文名。
产我国新疆的粉绿香蒲与国外分布的植物在形态上有一定的差异, 表现在叶片(宽度)、雌花序(大小)、雌花序与雄花序的间距、柱头(形态、大小)等性状方面(表 1)。在叶(叶片宽度、叶鞘形态)、雌花序(颜色、大小)、雌花(大小、孕性雌花的柱头、丝状毛、小苞片以及不孕雌花的子房形态)等许多重要的分种特征方面, 粉绿香蒲表现出介于宽叶香蒲和水烛两种之间的中间类型的性状。一方面, 它的雌花序深棕色, 雌雄花序有时相互靠近或紧密连接,叶片具渐狭的鞘, 孕性雌花柱头条状披针形, 不孕雌花子房倒圆锥形, 与宽叶香蒲的特征接近(图版Ⅰ: 2, 5, 7, 11); 另一方面, 它的叶片有时具耳状叶鞘,孕性雌花具小苞片, 雌雄花序有时远离等特征, 又与水烛的许多特征相符(图版Ⅰ: 9, 14)。同时它又有明显的可鉴别特征, 如花粉粒除单体、四合体外, 还有二体、三体出现, 花粉粒常败育; 柱头条状披针形(比宽叶香蒲的卵状披针形柱头窄, 但比水烛的线形柱头宽); 雌花小苞片浅褐色至先端白色(宽叶香蒲雌花无小苞片, 水烛小苞片浅褐色至深褐色), 比柱头窄等(图版Ⅰ: 6—11, 20)。上述区别特征国外很多学者都有记述[7—9,12,13]。另外, 粉绿香蒲与T. × provincialis Camus不同, 后者通常无耳状叶鞘, 在叶片基部和叶鞘的近轴面有黏液状腺体等; 与T. angustifolia × T. domingensis也不同, 后者花粉单体、小苞片急尖至渐尖等[8]。目前四合体花粉粒在香蒲属中见于宽叶香蒲、小香蒲(T. minima Funck)、T. alelseevii Mavrodiev、T. tzvelevii Mavrodiev、粉绿香蒲和T. × provincialis, 而二体和三体花粉粒则仅见于粉绿香蒲[1,2,4,7,13]。

表1 粉绿香蒲的形态变异Tab. 1 Comparison of morphological characters of Typha × glauca between our study and other studies
根据文献记载, 粉绿香蒲在北美是由于20世纪水烛从大西洋沿岸向其他地区迅速扩散并与乡土种宽叶香蒲杂交产生的, 在其亲本水烛和宽叶香蒲分布的重叠区普遍存在[9,12]。它依赖营养繁殖, 具有较强的竞争优势与入侵扩张能力和较高的净初级生产力,这不仅增加了底泥中有机物含量, 也影响了湿地植物群落的结构和功能以及沼泽演替方向[8—10,14—16,18,19]。水烛和宽叶香蒲在我国西北、东北、华北、华中和华南等地均有广泛的分布, 两物种也有较大的分布重叠区, 理论上推测粉绿香蒲在我国也应该有较广的分布, 但我们检查了我国主要植物标本馆馆藏中所有鉴定为水烛和宽叶香蒲的标本, 却仅在我们采自新疆的标本中发现有粉绿香蒲, 该杂种在我国的地理分布范围尚有待于进一步研究。下面给出粉绿香蒲与亲本近缘种的分种检索表, 以更好的了解其鉴别特征。
分种检索表
1. 雌花无小苞片; 柱头披针形或卵状披针形; 花粉四合体; 叶宽5—25 mm; 叶通常与花序等长或稍超出..........................................................................................................................................宽叶香蒲 T. latifolia
1. 雌花具小苞片; 柱头条形、线形或条状披针形; 花粉粒全部或部分单体; 叶明显超出花序。
2. 雌花序浅棕色; 柱头条形或线形; 小苞片先端浅棕色至深棕色, 比柱头宽; 花粉粒单体; 叶宽3—8 mm ........................................................................................................................................水烛 T. angustifolia
2. 雌花序深棕色; 柱头条状披针形; 小苞片先端灰白色或近于无色, 比柱头窄; 花粉粒常单体、二体、三体和四合体混合; 叶宽10—16 mm........................................................................粉绿香蒲 T. × glauca
致谢:
感谢中国科学院植物研究所植物标本馆(PE)马欣堂先生和田希娅女士、中国科学院华南植物园植物标本馆(IBSC)杨亲二研究员、中国科学院武汉植物园植物标本馆(HIB)李建强研究员、中国科学院成都生物研究所植物标本馆(CDBI)高信芬研究员、武汉大学植物标本馆(WH)汪小凡教授和四川大学植物标本馆(SZ)何兴金教授在检查标本中给予的热情帮助。
[1] Graebner P. Typhaceae. In: Engler A. Das Pflanzenreich [M]. Leipzig. 1900, 8(2): 8—16
[2] Mavrodiev E V. New species of cat-tail (Typha L.) from Caucasus [J]. Feddes Repertorium, 1999, 110(1—2): 127—132
[3] Mavrodiev E V. A new species of cat-tail (Typha L.) from sect. Engleria (Leonova). N. Tzvel [J]. Feddes Repertorium, 2000, 111(7—8): 571—575
[4] Mavrodiev E V. Two new species of Typha L. (Typhaceae Juss.) from the Far East of Russia and from Mongolia [J]. Feddes Repertorium, 2002, 113(3—4): 281—288
[5] Sun K, Simpson D A. Flora of China [M]. Beijing: Science Press. 2010, 161—163
[6] Zhao H, Xie H, Li G, et al. An application of Q-cluster analysis on the study of the classification of hydrophyte genus Typha in northeast China [J]. Acta Hydrobiologica Sinica, 1998, 22(1): 86—89 [赵宏, 谢航, 李刚, 等. Q型聚类分析在东北水生植物香蒲属分类研究中的应用. 水生生物学报, 1998, 22(1): 86—89]
[7] Hotchkiss N, Dozier H L. Taxonomy and distribution of North American cattails [J]. American Midland Naturalist, 1949, 4l(1): 237—254
[8] Smith S G. Experimental and natural hybrids in North America Typha (Typhaceae) [J]. American Midland Naturalist, 1967, 78(2): 257—287
[9] Grace J B, Harrison J S. The Biology of Canadian Weeds. 73. Typha latifolia L., Typha angustifolia L. and Typha × glauca Godr. [J]. Canadian Journal of Plant Science, 1986, 66(2): 361—379
[10] Smith S G. Typha: its taxonomy and the ecological significance of hybrids [J]. Archiv für Hydrobiologie, 1987, 27(1): 129—138
[11] Thieret J W, Luken J O. The Typhaceae in the southeastern United States [J]. Harvard Papers in Botany, 1996, 8, 27—56
[12] Kuehn M M, White B N. Morphological analysis of genetically identified cattails Typha latifolia, Typha angustifolia, and Typha × glauca [J]. Canadian Journal of Botany, 1999, 77(6): 906—912
[13] Finkelstein S A. Identifying pollen grains of Typha latifolia, Typha angustifolia, and Typha × glauca [J]. Canadian Journal of Botany, 2003, 81(9): 985—990
[14] Bonnewell V, Pratt D C. Effects of nutrients on Typha angustifolia × latifolia: productivity and morphology [J]. Journal of the Minnesota Academy of Science, 1978, 44(1): l8—20
[15] Shay J M, Shay C T. Prairie marshes in western Canada, with specific reference to the ecology of five emergent macrophytes [J]. Canadian Journal of Botany, 1986, 64(2): 443—454
[16] Galatowitsch S M, Andeson N O, Ascher P D. Invasiveness in wetland plants in temperate North America [J]. Wetlands,1999, 19(4): 733—755
[17] Sarena M S, Allison A S. The potential for hybridization between Typha angustifolia and Typha latifolia in a constructed wetland [J]. Aquatic Botany, 2004, 78(4): 361—369
[18] Nancy C T, Daniel J L, Pamela G, et al. Patterns of environmental change associated with Typha × glauca invasion in great lakes coastal wetland [J]. Wetlands, 2009, 29(3): 964—975
[19] Larkin D J, Freyman M J, Lishawa S C, et al. Mechanisms of dominance by the invasive hybrid cattail Typha × glauca [J]. Biological Invasions, 2012, 14(1): 65—77
[20] Fernald M L. Gray’s Manual of Botany [M]. 8thed. New York: American Book Company. 1950, 60—61
[21] Fassett N C, Calhoun B. Introgression between Typha latifolia and T. angustifolia [J]. Evolution, 1952, 6(4): 367—379
[22] Lee D W, Fairbrothers D E. A serological and disc eletrophoretic study of North American Typha [J]. Brittonia, 1969, 21(3): 227—243
[23] Bayly I L, O’Neill T A. A study of introgression in Typha at Point Pelee Marsh, Ontario [J]. Canadian Field Naturalist, 1971, 85(4): 309—314
[24] Sharitz R R, Wineriter S A, Smith M H, Liu E H. Comparison of isozymes among Typha species in the eastern United States [J]. Canadian Journal of Botany, 1980, 67(9): 1297—1303
TYPHA × GLAUCA GODR., A HYBRID OF TYPHA L. (TYPHACEAE) NEWLY RECORDED IN CHINA AND ITS MORPHOLOGICAL CHARACTERISTICS
ZHU Xiu-Yu and WANG Dong
(School of Life Sciences, Central China Normal University, Wuhan 430079, China)
Typha L. (Typhaceae), one of the most common aquatic plants found in marshes and shallow water, is almost cosmopolitan. However, the distribution of approximately 24 species has two main centers: Eurasia and North America. To date 12 species have been reported in China. Typha was described by Linnaeus in 1753, and its taxonomy remains confusing because of the variability in its reproductive and vegetative characteristics, and its frequent hybridization. Seven hybrids of Typha species were reported from North America and Europe but none of them has been recorded in China. We have discovered a hybrid cattail, Typha × glauca Godr. (T. angustifolia L. × T. latifolia L.), through extensive field collections and herbarium studies. That was a newly recorded taxon for China. We studied the hybrid T. × glauca in the field and examined Typha collections from the following herbaria including PE, IBSC, CDBI, HIB, WU, SZ, and CCNU. Both vegetative and reproductive characteristics were measured for available specimens of the hybrid and parental species from China. Pollen grains of Typha species were collected from our field collections, and were examined under a light microscope at × 1000 magnification. Morphological features of the hybrid and parental species was illustrated and discussed, and an identification key for them was provided. The hybrid was morphologically variable and generally intermediate to the parental species. The most readily diagnostic features of the hybrid were: 1. leaves were glaucous; 2. pollen was typically a mixture of monads, dyads, triads, and tetrads (vs. tetrads in T. latifolia and monads in T. angustifolia), often abortive; 3. pistillate bracteoles were narrower than the stigma (vs. broader in T. angustifolia and absent in T. latifolia), and 4. the stigmas were liner-lanceolate (vs. linear in T. angustifolia, and lanceolate to ovate-lanceolate in T. latifolia). Importantly, the presence /or absence of dyads and triads of pollen grains could be used to distinguish the hybrid and parental species, implying that pollen morphology is of importance in taxonomic treatment of genus Typha. In North America, Typha × glauca is a well-known cattail hybrid and can occur wherever T. angustifolia and T. latifolia grow together and often out-competes the parental species in habitats that are heavily disturbed or subject to high-magnitude water level fluctuations. The hybrid plants could interfere with wetland communities by forming large monospecific stands, out-competing native species, and altering substrate characteristics, suggesting that the appearance of hybrid plants may be used as indicators of disturbance and, perhaps, of wetland health. After checking the available specimens assigned to either T. angustifolia or T. latifolia that were deposited in the Herbaria of China, we found that T. × glauca was confined to the Xinjiang Province of China. Given the paucity of field collections on Typha plants over the past several decades and thus resultant relatively limited information on the hybrid in China, the distributional range and its ecological role of the hybrid cattail need to be studied further. In summary, we concluded that the field identification characters of the leaves and inflorescences may be used with some degree of reliability to tentatively separate T. × glauca from the parental species, but microscopic floral characters should be used to confirm the identification. To positively identify T. × glauca and parental species microscopic floral characters must be used.
Typha L.; T. angustifolia L.; T. latifolia L.; T. × glauca; New record
Q949.7
A
1000-3207(2013)01-0029-05
10.7541/2013.29
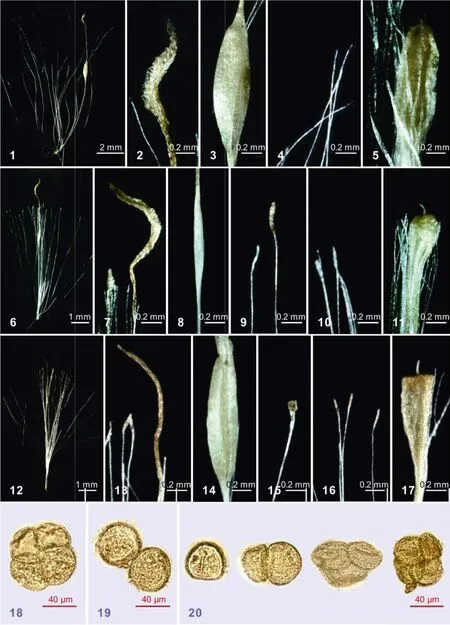
图版I PlateⅠ
1—5: 宽叶香蒲(1. 孕性雌花; 2. 柱头; 3. 孕性子房; 4. 丝状毛; 5. 不孕子房); 6—11: 粉绿香蒲(6. 孕性雌花; 7. 柱头; 8. 孕性子房; 9.小苞片; 10. 丝状毛; 11. 不孕子房); 12—17: 水烛(12. 孕性雌花; 13. 柱头; 14. 孕性子房; 15. 小苞片; 16. 丝状毛; 17. 不孕子房); 18—20: 花粉粒形态(18. 宽叶香蒲, 示四合体; 19. 水烛, 示单体; 20. 粉绿香蒲, 示单体、二体、三体、四体)
1—5: Typha latifolia (1. fertile flower; 2. stigma; 3. fertile ovary; 4. hairs; 5. sterile ovary); 6—11; T. × glauca (6. fertile flower; 7. stigma; 8. fertile ovary; 9. pistillate bracteoles; 10. hairs; 11. sterile ovary); 12—17: T. angustifolia (12. fertile flower; 13. stigma; 14. fertile ovary; 15. pistillate bracteoles; 16. hairs; 17. sterile ovary); 18—20: pollen grains (18. tetrad in T. latifolia; 19. monads in T. angustifolia; 20. the mixture of monad, dyad, triad, and tetrad in T. × glauca
2011-12-06;
2012-04-13
国家自然科学基金项目(30870151); 中国科学院植物研究所系统与进化植物学国家重点实验室开放课题(LSEB2011-06); 国家自然科学基金委重大国际合作项目(31110103911); 中国科学院战略性先导科技专项—应对气候变化的碳收支认证及相关问题(XDA05050206)项目资助
朱秀玉(1984—), 女, 山东临沂人; 硕士研究生; 主要从事植物分类学和生态学研究。E-mail: wetlandplant@163.com
王东, E-mail: dongwang.cn@gmail.com
