铵胁迫对狐尾藻(Myriophyllum spicatum)和金鱼藻(Ceratophyllum demersum)生物量分配和形态的影响*
2013-05-28袁桂香钟家有倪乐意朱天顺
袁桂香,符 辉,钟家有,倪乐意,朱天顺,李 威,宋 鑫
(1:中国科学院水生生物研究所,东湖湖泊生态系统实验站,淡水生态与生物技术国家重点实验室,武汉 430072)
(2:江西省水利科学研究院,南昌 330029)
(3:中国科学院大学,北京 100049)
随着湖泊水体富营养化的加剧,水体营养盐不断增加,透明度下降,蓝藻水华不断暴发,导致很多湖泊沉水植被生物量和分布面积不断下降,生物多样性锐减,大量沉水植物濒临灭绝[1-4].水体营养盐的适当增加能有效促进植物生长,而过高的营养盐浓度会直接或间接地抑制植物的生长,改变植物的生物量分配方式[5-8].其中,高浓度的铵态氮对沉水植物生长的影响已经引起国内外学者的广泛关注[5,9-16].对我国长江中下游31 个湖泊的调查研究发现当水体铵态氮浓度大于0.56 mg/L 时,沉水植物苦草(Vallisneria natans)几乎全部消失[5].目前尽管很多湖泊水体铵态氮含量低于这一阈值,但底泥间隙水中铵态氮含量往往是水体中的数倍[5,17-18].流水和风浪的扰动能使间隙水中铵态氮释放到水体中,引起水体中铵态氮含量短时间内显著增加[19-21],对沉水植物造成胁迫.
高浓度的铵态氮不仅降低植物叶绿素含量[14,16,22],还会抑制植物呼吸作用,影响光合作用的电子传递链[23].为了防止铵态氮对自身的毒害,通常植物会将体内的铵态氮快速地转化成游离氨基酸,在转化过程中由α-酮戊二酸提供碳骨架,使植物体内可溶性糖的含量随之减少[24-28].同时植物会将体内过多的铵态氮排出体外,在这过程中消耗大量的能量[29-30].研究表明,在胁迫条件下,植物会将更多的生物量分配给能获取生长短缺资源的部分[31].铵胁迫下,由于体内碳水化合物减少,植物会将更多的生物量分配给地上部分来增强光合作用,从而弥补铵胁迫对其影响[7].Cramer 等研究发现,与低铵浓度相比,高铵浓度下小麦(Triticum aestivum)和玉米(Zea mays)的植物干重均显著下降,而地上部分与地下部分的生物量比值增加[7].
铵胁迫对沉水植物的影响主要集中在其对体内碳氮代谢等生理方面的研究,如铵胁迫环境下沉水植物体内的游离氨基酸含量增加,可溶性碳水化合物、淀粉和可溶性蛋白质含量减少[5,6,9,11,32-33].然而铵胁迫及铵胁迫解除后对沉水植物生物量分配和形态影响的报道少见.本研究选取两种常见的沉水植物——狐尾藻(Myriophyllum spicatum L.)和金鱼藻(Ceratophyllum demersum L.),探讨其生物量、生物量分配形式和植物形态对铵胁迫以及铵胁迫解除后的响应,为富营养化湖泊水体中沉水植物的修复提供参考.
1 材料与方法
1.1 实验材料
实验地点位于云南省大理市洱海湖滨带(25°52'N,100°06'E),实验场地顶部支架覆盖一层遮阳网.实验用狐尾藻和金鱼藻于2011年8月底采自洱海喜州镇码头附近的水域,选取长势良好且无损伤的植物顶端15 cm 长,保留顶部7 cm 以内的叶子,去掉其余的叶子和分枝.两种沉水植物分别挑选384 株,种植前用自来水洗净待用.将挑选的植株种植于的塑料杯中(直径×高=6.0 cm×7.5 cm),每杯种植1 株.杯子底部盛有6 cm 采自洱海的底泥,底泥顶部覆盖1 cm 的细沙,细沙购买自附近沙石场且用自来水清洗干净.随后将种有植株的塑料杯放于水深30 cm(水体积60 L)的种植箱中(长×宽×高=50 cm×40 cm×35 cm),每箱放置48 杯,每种植物8 箱,共16 箱.预培养时间为15 d.
1.2 实验设计
实验设置处理组(加铵态氮)和对照组,并分为铵胁迫和铵胁迫解除两个时间阶段.铵胁迫阶段:每天早上往每种植物其中4 个种植箱中加入确定浓度的氯化铵溶液使水体铵态氮(NH4+-N)浓度维持在2 mg/L,作为处理组;另外4 个种植箱不加氯化铵作为对照组,铵态氮处理维持4 d.每天测定水体铵态氮浓度,以确定氯化铵溶液的加入量.铵胁迫解除阶段:在铵态氮处理第4 d 对所有种植箱中水体统一用自来水置换,之后让植物持续生长10 d.该实验从2011年8月22日开始,持续29 d,期间每隔4 d 于中午测定1 次水表面光合有效辐射(photosynthetic active irradiation,PAR)、水温和水化指标.实验期间水温为19 ~22℃,PAR 为350 ~460 μmol/(m2·s),水体硝态氮(NO3--N)、NH4+-N 和正磷酸盐(-P)分别为 0.69 ±0.09、0.010 ±0.001和0.012 ±0.002 mg/L.每周用软毛刷轻轻擦拭植物叶片表面,以避免附着生物对植物生长的影响.
1.3 参数测定
实验开始后,每天早上(铵胁迫阶段为加铵之前)每个种植箱取3 株植物.每次取样时将植株洗净,再用蒸馏水清洗3 次,然后每株分别称重、测定株高,之后根、茎、叶分开分别称重.用根生物量/单株生物量、茎生物量/单株生物量和叶生物量/单株生物量3 个参数表征植株的生物量分配方式.由于金鱼藻为无根植物,文中提到两种植物根、茎、叶是指狐尾藻的根、茎和叶3 个部分,而金鱼藻仅为茎和叶2 部分.
1.4 数据分析
一般线性模型分析采用SPSS 13.0 软件进行统计分析,用于分析铵胁迫处理、取样时间和它们的交互作用对两种沉水植物生物量、株高和生物量分配方式的影响,其中铵盐处理为固定变量,取样时间为协变量.初步分析结果发现它们的交互作用对所有测定参数均无显著影响(P >0.05),所以本文中排除了交互作用的影响,未纳入结果分析.统计分析时,将每个种植箱3 个植株的测量数据平均处理,作为一个重复单元格数据.用0 d 表示铵胁迫前,1 ~4 d 为铵胁迫阶段,5 ~14 d 为铵胁迫解除阶段.
2 结果
2.1 铵态氮对生物量的影响
铵胁迫阶段,处理组2 种沉水植物的生物量与对照组无显著差别(表1),处理组和对照组狐尾藻单株生物量分别为 1.62 ±0.05 和 1.51 ±0.06 g,金鱼藻单株生物量分别为 2.03 ±0.06 和 2.10 ±0.04 g.铵胁迫解除阶段,处理组金鱼藻单株生物量较对照组显著增加(表1,图1),分别为2.53 ±0.05 和2.42 ±0.04 g;而狐尾藻单株生物量与对照组无显著差别(表1,图1),分别为1.91 ±0.05 和1.86 ±0.04 g.铵胁迫阶段和铵胁迫解除阶段,在不同取样时间狐尾藻单株生物量差异显著,而金鱼藻单株生物量仅在铵胁迫解除阶段差异显著(表1,图1).
2.2 铵态氮对株高的影响
铵胁迫阶段,与对照组相比,处理对狐尾藻株高无显著影响,然而金鱼藻株高显著增加(表1,图1),处理组和对照组中狐尾藻株高分别为25.35 ±0.43 和24.53 ±0.49 cm,金鱼藻分别为19.29 ±0.13 和18.68 ±0.18 cm.而在铵胁迫解除阶段,处理组2 种沉水植物株高均较对照组显著增高(表1,图1),狐尾藻株高分别为 29.01 ±0.38 和 27.67 ±0.39 cm,金鱼藻分别为 21.38 ±0.20 和 20.44 ± 0.17 cm.总体来看,2 种沉水植物株高均随取样时间增长而增高(表1,图1).
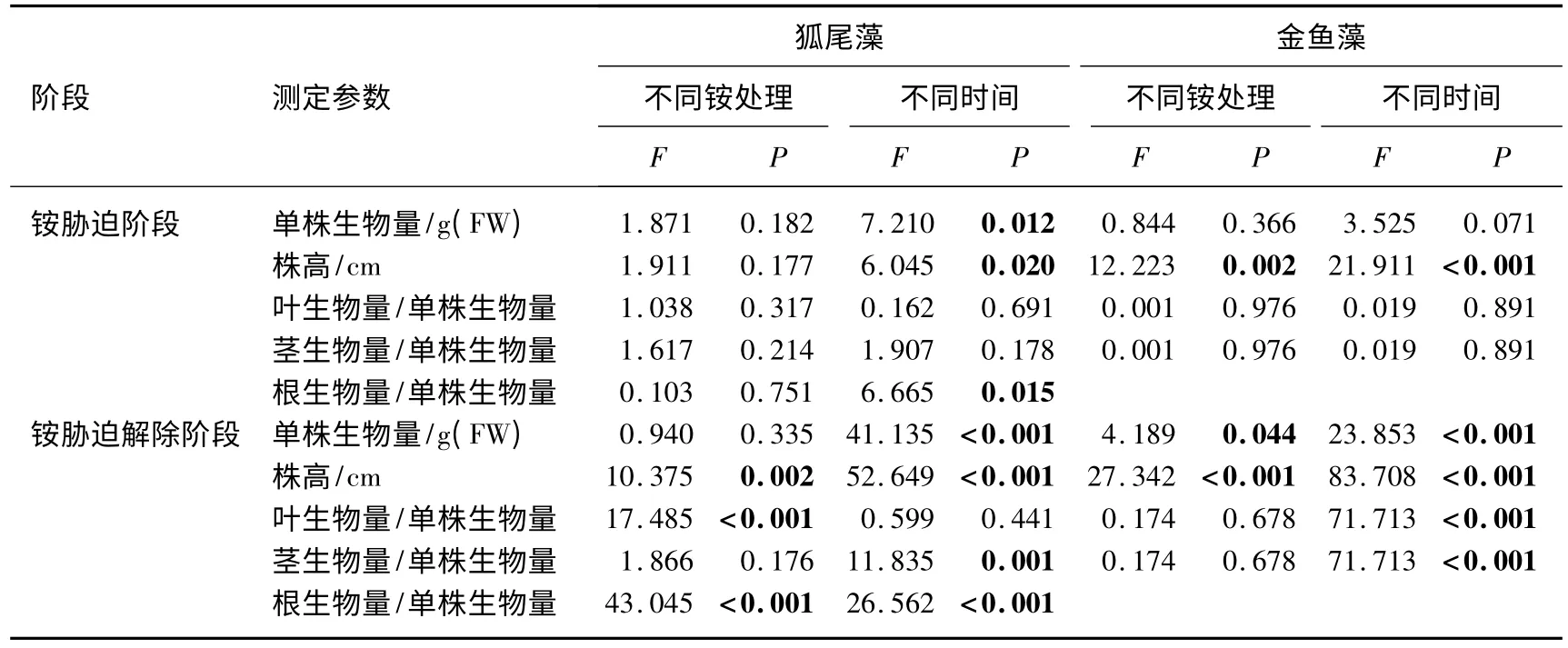
表1 狐尾藻和金鱼藻铵处理和取样天数的多变量二元方差分析结果*Tab.1 Multivariate two-way ANOVAs on ammonium enrichment treatment and sampling days for M.spicatum and C.demersum
2.3 铵态氮对生物量分配的影响
铵胁迫阶段,与对照组相比,处理对2 种沉水植物根生物量/单株生物量、茎生物量/单株生物量、叶生物量/单株生物量均无显著影响(表1,图2).在铵胁迫解除阶段,与对照组相比,处理组狐尾藻叶生物量/单株生物量显著增加,处理组和对照组分别为0.510 ±0.004 和0.490 ±0.004,而根生物量/单株生物量显著下降(表1,图2),分别为0.180 ±0.004 和0.210 ±0.004,茎生物量/单株生物量保持不变,分别为0.310 ±0.005 和0.300 ±0.004;金鱼藻茎生物量/单株生物量和叶生物量/单株生物量无显著差异.

图1 处理组和对照组中狐尾藻和金鱼藻的株高和单株生物量Fig.1 Individual biomass and shoot height of M.spicatum and C.demersum in ammonium enrichment group and control group
3 讨论
对沉水植物的生长研究发现,经过半个月的生长,不同沉水植物生物量积累存在明显差异[34].而本研究中生长14 d,狐尾藻处理组和对照组单株生物量无显著差异,说明2 mg/L 的铵态氮对狐尾藻生长没有影响;而金鱼藻单株生物量在铵胁迫解除阶段显著增加.说明较高浓度铵态氮处理4 d 对2 种沉水植物的生物量积累没有显著的抑制作用.
以往野外调查研究结果认为,铵态氮浓度大于0.56 mg/L 时就会抑制沉水植物苦草生长[5].本研究与其结果不同.这两种研究结果的不同可能与时间尺度和生长环境有关.有研究发现较高光照能有效缓解铵胁迫对沉水植物的影响,在强光照环境下菹草(Potamogeton crispus)体内可溶性碳水化合物含量较低光照环境显著增加[11].同样高铵态氮浓度条件下,金鱼藻中淀粉含量在实验环境光照条件下要显著高于遮光处理[33].本研究中尽管进行了遮光处理,但水表面PAR 仍然较其他实验系统高出2 倍多[11,33],这促进了碳水化合物的积累,从而为沉水植物对铵的大量吸收提供能量基础,并有效缓解了铵态氮对2 种沉水植物的毒性效应.
铵胁迫解除阶段,处理组金鱼藻生物量积累显著增加,狐尾藻生物量也有所增加,这可能是由于在铵胁迫阶段植物体内积累了大量的铵态氮和由铵态氮转化合成的游离氨基酸[9,24,33],当铵胁迫解除后,充足的光照条件使植物体内碳资源不断得到补充,有利于体内积累的铵态氮继续向游离氨基酸转化,最终被植物吸收利用.相反,对照组中可能由于氮资源缺乏,生物量积累反而较处理组低.在铵胁迫解除阶段,处理组金鱼藻和狐尾藻株高均显著增加,这可能与铵处理组较高的生物量积累有关.
在铵胁迫解除阶段,与对照组相比狐尾藻生物量分配方式发生改变,将更多的生物量分配给叶子,而相对减少了对根的生物量分配.实验观察发现在铵胁迫解除阶段,处理组狐尾藻叶子边缘出现明显的发黄现象.这说明铵胁迫阶段积累的铵态氮在生长后续阶段仍然可能存在毒害作用,为了获取更多的碳资源,将体内的氮资源进行转化吸收利用,维持体内的碳氮代谢平衡,狐尾藻将更多的生物量分配给叶子部分,而相对减少了对根的生物量分配.
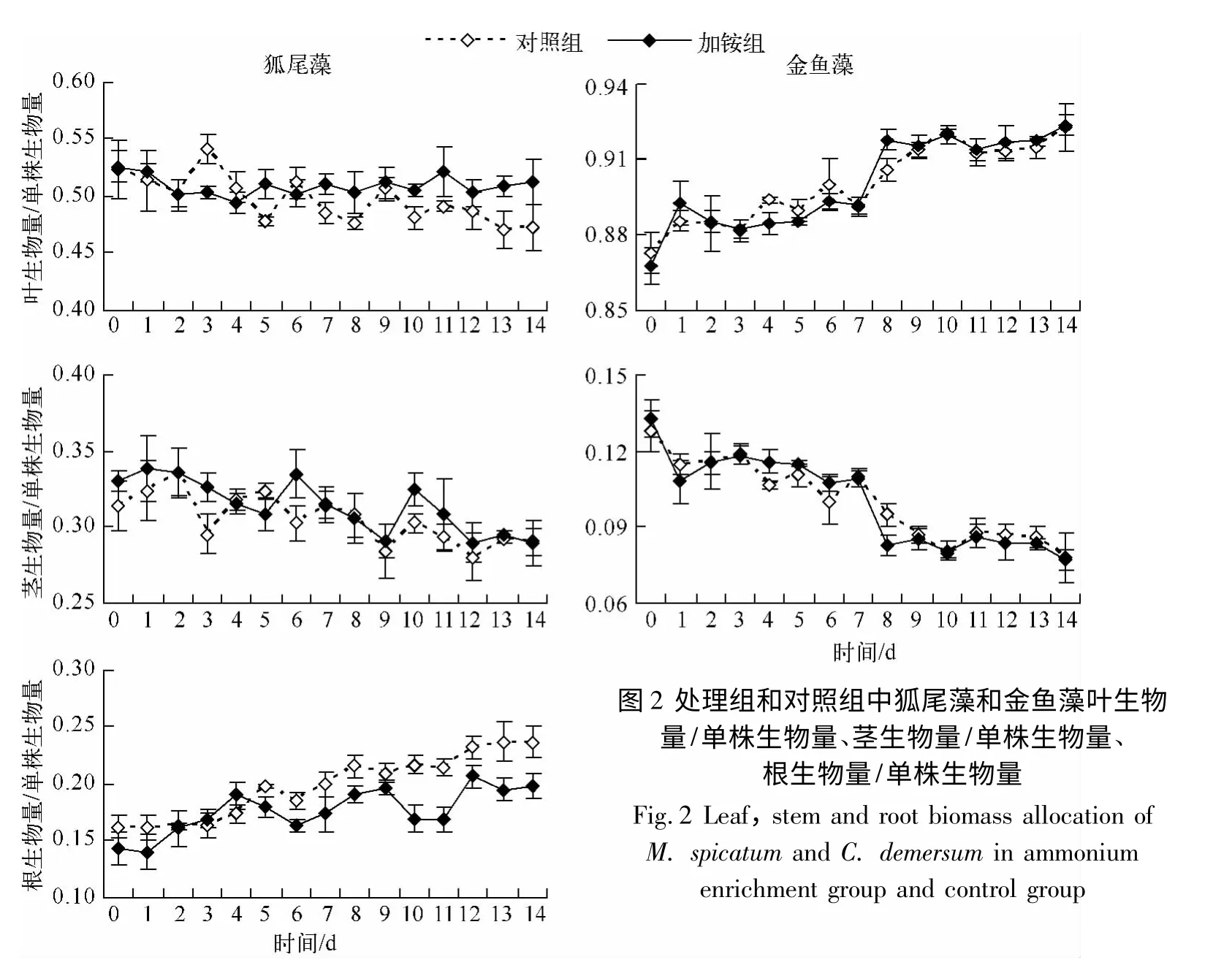
图2 处理组和对照组中狐尾藻和金鱼藻叶生物量/单株生物量、茎生物量/单株生物量、根生物量/单株生物量Fig.2 Leaf,stem and root biomass allocation of M.spicatum and C.demersum in ammonium enrichment group and control group
用0.16 mg/L 铵态氮处理并持续两个月的实验研究发现,金鱼藻积累了更多的淀粉,而狐尾藻积累了更多的氮[33].在水体氮含量充足的情况下,较多的碳资源积累更有利于维持体内的碳氮代谢平衡,说明金鱼藻较狐尾藻能更好地适应高铵态氮浓度的环境[33].本文研究结果与其一致,即金鱼藻生物量积累在处理组较对照组显著增加,而狐尾藻生物量积累增加不显著.
综上所述,在高光照条件下,铵态氮浓度的短时间增加对沉水植物的生长不会产生抑制作用,反而可能促进植物的生长,这表明自然水体中来自外源或内源沉积物库的脉冲式营养输入有可能促进富营养化初期沉水植物扩增.由于不同沉水植物碳氮代谢差异,在铵态氮增加时对其利用程度也不同而表现出种间差异和优势变化.然而,需要关注的是,在富营养化湖泊中,由于弱光胁迫导致植物体内碳水化合物短缺,这可能使沉水植物对环境变化的敏感程度增加.
[1]Blindow I.Decline of charophytes during eutrophication:comparison with angiosperms.Freshwater Biology,1992,28:9-14.
[2]Jones JI,Young JO,Eaton JW et al.The influence of nutrient loading,dissolved inorganic carbon and higher trophic levels on the interaction between submerged plants and periphyton.Journal of Ecology,2002,90:12-24.
[3]Middelboe AL,Markager S.Depth limits and minimum light requirements of freshwater macrophytes.Freshwater Biology,1997,37:553-568.
[4]Phillips GL,Eminson D,Moss B.A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters.Aquatic Botany,1978,4:103-126.
[5]Cao T,Xie P,Ni L et al.The role of NH4+toxicity in the decline of the submersed macrophyte Vallisneria natans in lakes of the Yangtze River basin,China.Marine and Freshwater Research,2007,58:581-587.
[6]Li H,Cao T,Ni L.Effects of ammonium on growth,nitrogen and carbohydrate metabolism of Potamogeton maackianus A.Benn.Fundamental and Applied Limnology,2007,170:141-148.
[7]Cramer MD,Lewis OAM.The influence of nitrate and ammonium nutrition on the growth of wheat (Triticum aestivum)and maize (Zea mays)plants.Annals of Botany,1993,72:359-365.
[8]高镜清,燕启社,黄五星等.富营养化水体中铵态氮对金鱼藻生长的影响.长江流域资源与环境,2011,20:611-616.
[9]Cao T,Xie P,Ni L et al.Carbon and nitrogen metabolism of an eutrophication tolerative macrophyte,Potamogeton crispus,under NH4+stress and low light availability.Environmental and Experimental Botany,2009,66:74-78.
[10]Li W,Zhang Z,Jeppesen E.The response of Vallisneria spinulosa (Hydrocharitaceae)to different loadings of ammonia and nitrate at moderate phosphorus concentration:a mesocosm approach.Freshwater Biology,2008,53:2321-2330.
[11]Zhang M,Cao T,Ni L et al.Carbon,nitrogen and antioxidant enzyme responses of Potamogeton crispus to both low light and high nutrient stresses.Environmental and Experimental Botany,2010,68:44-50.
[12]许秋瑾,金相灿,王兴民等.不同浓度铵态氮对镉胁迫轮叶黑藻生长及抗氧化酶系统的影响.应用生态学报,2007,18:420-424.
[13]焦立新,王圣瑞,金相灿.穗花狐尾藻对铵态氮的生理响应.应用生态学报,2009,20:2283-2288.
[14]高镜清,黄五星,黄 宇等.铵态氮胁迫下金鱼藻的过氧化损伤和抗氧化能力.武汉大学学报:理学版,2010,56:590-596.
[15]马剑敏,靳同霞,贺 锋等.菹草对铵态氮和硝氮急性胁迫的响应.环境科学与技术,2009,32:26-30.
[16]宋 睿,姜锦林,耿金菊等.不同浓度铵态氮对苦草的生理影响.中国环境科学,2011,31:448-453.
[17]Cao T,Xie P,Li Z et al.Physiological stress of high NH4+concentration in water column on the submersed macrophyte Vallisneria natans L..Bulletin of Environmental Contamination and Toxicology,2009,82:296-299.
[18]郭洪涛,曹 特,倪乐意.中等实验规模下不同营养环境对苦草(Vallisneria natans)生长的影响.湖泊科学,2008,20:221-227.
[19]范成新,张 路,秦伯强.太湖沉积物-水界面生源要素迁移机制及定量化——1.铵态氮释放速率的空间差异及源-汇通量.湖泊科学,2004,16:10-16.
[20]逄 勇,颜润润,余钟波等.风浪作用下的底泥悬浮沉降及内源释放量研究.环境科学,2008,29:2456-2464.
[21]李一平,逄 勇,吕 俊等.水动力条件下底泥中氮磷释放通量.湖泊科学,2004,16:318-324.
[22]Rudolph H,Voigt JU.Effects of NH4+-N and NO3--N on growth and metabolism of Sphagnum magellanicum.Physiologia Plantarum,1986,66:339-343.
[23]Vines HM,Wedding RT.Some effects of ammonia on plant metabolism and a possible mechanism for ammonia toxicity.Plant Physiology,1960,35:820-825.
[24]Cruz C,Bio AFM,Domínguez-Valdivia MD et al.How does glutamine synthetase activity determine plant tolerance to ammonium?Planta,2006,223:1068-1080.
[25]Magalhäes JR,Huber DM.Ammonium assimilation in different plant species as affected by nitrogen form and pH control in solution culture.Nutrient Cycling in Agroecosystems,1989,21:1-6.
[26]Magalhães JR,Ju GC,Rich PJ et al.Kinetics of15NH4+assimilation in Zea mays.Plant Physiol,1991,94:647-656.
[27]Mark S.Nitrate regulation of metabolism and growth.Current Opinion in Plant Biology,1999,2:178-186.
[28]Britto DT,Kronzucker HJ.Nitrogen acquisition,PEP carboxylase,and cellular pH homeostasis:new views on old paradigms.Plant,Cell and Environment,2005,28:1396-1409.
[29]Britto DT,Siddiqi MY,Glass ADM et al.Futile transmembrane NH4+cycling:A cellular hypothesis to explain ammonium toxicity in plants.Proceedings of the National Academy of Sciences,2001,98:4255-4258.
[30]Kronzucker HJ,Britto DT,Davenport RJ et al.Ammonium toxicity and the real cost of transport.Trends in Plant Science,2001,6:335-337.
[31]McCarthy MC,Enquist BJ.Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation.Functional Ecology,2007,21:713-720.
[32]Cao T,Ni L,Xie P.Acute biochemical responses of a submersed macrophyte,Potamogeton crispus L.,to high ammonium in an aquarium experiment.Journal of Freshwater Ecology,2004,19:279-284.
[33]Cao T,Ni L,Xie P et al.Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability.Freshwater Biology,2011,56:1620-1629.
[34]Fu H,Yuan G,Cao T et al.Relationships between relative growth rate and its components across 11 submersed macrophytes.Journal of Freshwater Ecology,2012,27:471-480.
