墨龙与红鲫的视网膜和视盖解剖结构比较
2013-04-19安安树青华田苗
万 安安树青华田苗
(1. 南京大学生命科学学院, 南京 210093; 2. 安庆师范学院资源环境学院, 安庆 246000; 3. 安徽师范大学生命科学学院, 芜湖 241000)
墨龙与红鲫的视网膜和视盖解剖结构比较
万 安1,2安树青1华田苗3
(1. 南京大学生命科学学院, 南京 210093; 2. 安庆师范学院资源环境学院, 安庆 246000; 3. 安徽师范大学生命科学学院, 芜湖 241000)
墨龙是一种由红鲫进化来的龙睛种金鱼(Carassius auratus)。随机取体长10—12 cm, 重约35 g的墨龙和红鲫各4尾, 解剖取出整个眼球及脑, 并常规石蜡切片, HE染色。在光学显微镜下观察墨龙和红鲫的视网膜、视盖系统的显微结构变化并比较各层厚度, 发现与红鲫相比, 墨龙视网膜的总厚度下降29.9%, 其中外网状层厚度增加2.5%、内网状层厚度增加11.8%; 而内核层厚度下降21.6%、外核层厚度降低35.6%, 神经节细胞层、杆锥层也变薄, 且后两者分层不规则; 墨龙视盖壁整体厚度增加28.9%, 其中除围脑室层厚度减少22.6%外, 中央纤维层厚度增加12.8%, 中央细胞层厚度增加30.6%, 表面纤维层厚度增加21.9%, 且纤维远较红鲫密集, 视神经层厚度增加91.7%, 边缘层厚度增加35.6%。结果表明长期的人工选择不但改变了墨龙的外形, 而且使其中枢神经组织结构也发生了较大变化, 并推测墨龙的眼球直径及视网膜面积较大, 从而导致自视网膜传入视盖的纤维增多, 是视网膜和视盖中的传递神经冲动的神经元、神经纤维所在层段增厚的主要原因; 同时墨龙视网膜中色素上皮层向杆锥层交错对插, 富含神经元的视网膜外核层、内核层以及视盖中的围脑室层厚度也降低, 可以减少因视网膜面积大而造成的强光伤害; 此外由于墨龙的围脑室层厚度降低, 导致其游动及平衡能力较红鲫差。
金鱼; 墨龙; 红鲫; 视网膜; 视盖
金鱼也称“金鲫鱼”, 属鲤形目, 鲤科, 鲤亚科,鲫属, 与鲫鱼同用一个学名(Carassius auratus), 研究表明,银灰色的野生鲫鱼变为红黄色的红鲫(Red crucian), 然后再经过不同时期的驯养和选择培养,由红鲫逐渐变成为不同品种金鱼, 各种金鱼的体态出现了与自然环境下的鱼类明显不同的特征[1]。除外貌特征的改变外, 有学者发现墨龙(Black dragoneye)眼球体积及视网膜整体面积较红鲫大, 而厚度较薄[2]。鉴于此, 通过实验观察, 本文对金鱼中墨龙和红鲫的视网膜和视盖系统的显微结构进行进一步比较,以期有所发现。
本研究采用石蜡切片和HE染色技术,通过光镜对红鲫和墨龙两种金鱼的视网膜和中脑视盖的组织学结构进行了观察, 为深入研究长期人工选择条件下金鱼的感觉器官和神经系统的生理结构变化提供基础资料。
1 材料与方法
实验用金鱼100尾, 购自安庆市花鸟鱼虫市场,实验前置于实验室禁食驯养48h,随机取体长10—12 cm, 重约35 g的各4尾置于4℃水中麻醉, 用游标卡尺测量体长, 眼球直径, 从第3、第4脊椎处剪断, 将头部转移到Olympus SZ1145型体视显微镜下进行解剖, 将整个眼球及脑取出后, 迅速用4%多聚甲醛的PBS溶液固定48h。用PBS清洗后, 按常规方法制作石蜡切片, 连续冠状切, 厚度6 μm。
将所得切片做HE染色, Olympus BX51型显微镜观察并拍照, 每张切片上随机选取8个视野测量视网膜、视盖总厚度及各层厚度(Unint: μm), 并计算平均值。所有数据采用平均值±标准差表示, 用t-test进行各指标的显著性检验。
2 结果
2.1 体貌特征
测量结果显示, 实验所用红鲫和墨龙的体长相似, 但墨龙的眼球直径明显大于红鲫(表1)。
2.2 视网膜厚度变化
组织染色显示红鲫的视网膜7层结构清晰易辨(图版Ⅰ-1), 而墨龙神经节细胞层(Ganglion cell layer, GCL), 杆锥层(Photoreceptor layer, PL)和色素上皮层(Pigment epithelium layer, PEL)各层界限不分明, 厚度不易分辨, 其中色素上皮层向杆锥层延伸并交错对插(图版Ⅰ-2)。
选择两种鱼的视网膜总厚度以及较清晰的4层:外核层(Outer nuclear layer, ONL)、外网状层(Outer plexiform layer, OPL)、内核层(Inner nuclear layer, INL)和内网状层(Inner plexiform layer, IPL)进行统计比较。与红鲫相比, 墨龙的视网膜出现一些明显的变化(表2、表3): (1)视网膜总厚度平均下降29.9%,与红鲫相比具有极显著差异(P<0.001); (2)其中外核层厚度平均下降35.6%, 具有显著差异(P<0.05); (3)外网状层厚度平均增加2.5%; (4)内核层厚度平均下降21.6%, 具有显著差异(P<0.05); (5)内网状层厚度平均增加11.8%; (6)外网状层和内网状层占视网膜总厚度的比例较红鲫均出现显著增加; (7)神经节细胞层, 色素上皮层和杆锥层, 各层厚度不易分辨。

表1 红鲫与墨龙体貌比较Tab. 1 Comparison of physical appearance of black dragoneye and red crucian (n=4)
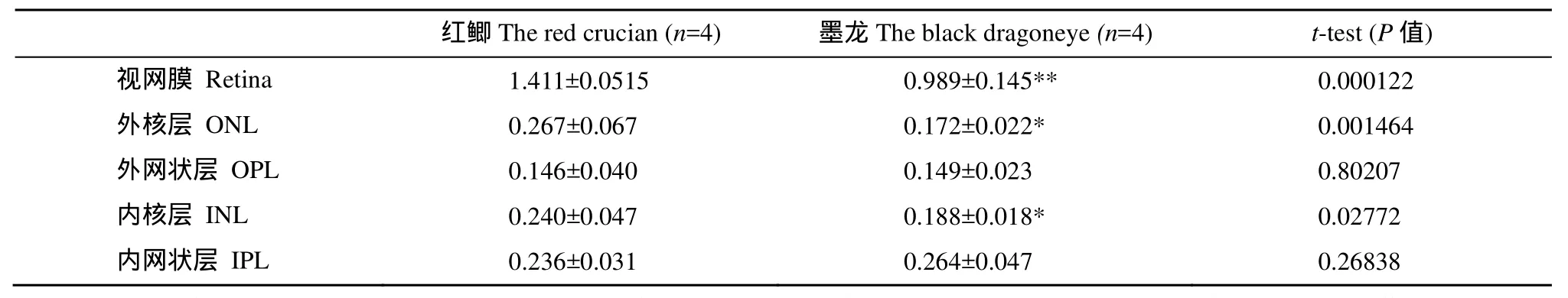
表2 红鲫、墨龙视网膜各层厚度比较Tab. 2 Comparison of the thicknesses of retina layers in black dragoneye and red crucian (μm)

表3 红鲫、墨龙视网膜各层在总厚度比值(相对厚度)比较Tab. 3 A comparison between the relative thickness of retina layers in black dragoneye’s and red crucian’s (%)
2.3 视盖各层厚度变化
组织染色显示两种金鱼的视盖结构清晰易辨,依据林浩然等[3,4]分6层(图版Ⅰ-3、4), 而与红鲫相比,墨龙的视盖出现一些明显的变化(表4、表5): (1) 视盖总厚度平均增加28.9%, 与红鲫相比具有极显著差异(P<0.001); (2) 围脑室层(Stratum periventriculare, SPV)较红鲫薄, 厚度减少22.6%, 与红鲫相比具有极显著差异(P<0.001); 围脑室层占视盖总厚度的百分比较红鲫显著降低; (3) 中央纤维层(Stratum album central, SAC) 较红鲫厚, 厚度增加12.8%; (4) 中央细胞层(Stratum griseum central, SGC) 较红鲫厚, 厚度增加30.6%, 与红鲫相比具有显著差异(P<0.05); (5) 表面纤维层(Stratum fibrosum et griseum superficiale, SFGS)较红鲫厚, 厚度增加21.9%, 与红鲫相比具有显著差异(P<0.05); 且两者结构质地明显有差异, 红鲫此层纤维密度较稀疏(图版Ⅰ-3、4)。(6) 视神经层(Stratum opticum, SO) 较红鲫厚, 厚度增加91.7%, 与红鲫相比具极显著差异(P<0.001); 视神经层占视盖总厚度的百分比出现显著增加。(7) 边缘层(Stratum marginal, SM)较红鲫厚, 厚度增加35.6%, 与红鲫相比具有显著差异(P<0.05)。
3 讨论
墨龙全体漆黑, 身体短而肥, 尾鳍分叉大, 喜静, 不善游动, 由红鲫驯化而来[1], 属龙睛类金鱼,其眼球过分膨大而突出于眼眶之外, 眼径明显大于红鲫。鱼类眼进化的特点: 一般白天活动靠视觉摄食的鱼类, 有突出的眼, 其直径大约相当于头长的l/5—1/6; 而黄昏或夜间活动、主要用视觉捕食的鱼,或至少部分靠视觉摄食的深海鱼类, 眼径可达头长的1/3—1/2[5]。墨龙眼径大, 推测其视网膜和视盖也应随之而发达。
实验发现, 与红鲫相比, 墨龙的眼径较红鲫大,视网膜面积也较大, 其视网膜却出现小眼径鱼类具有的特征[3,9—12]: (1)视网膜壁层总厚度较红鲫平均下降29.9%; 杆锥层(PL)、视神经节层(GCL)变薄,分层不规则; 感光细胞胞体所在的外核层(ONL)较薄, 推测其视网膜的感光能力较弱; (2)具有传递神经冲动作用的双极细胞、水平细胞所在的内核层(INL)[3], 含大量传导纤维的外网状层(OPL)、内网状层(IPL)较红鲫均显著增厚, 说明墨龙的视网膜整体输出的神经冲动较红鲫多。
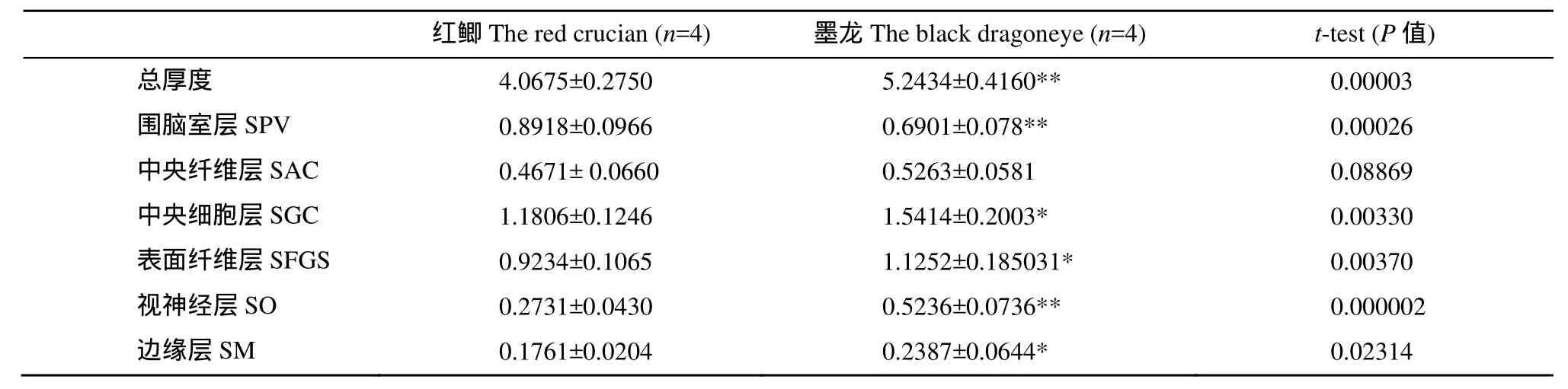
表4 红鲫与墨龙视盖各层厚度比较Tab. 4 A comparison of the optic tectum layers in black dragoneye and red crucian (μm)
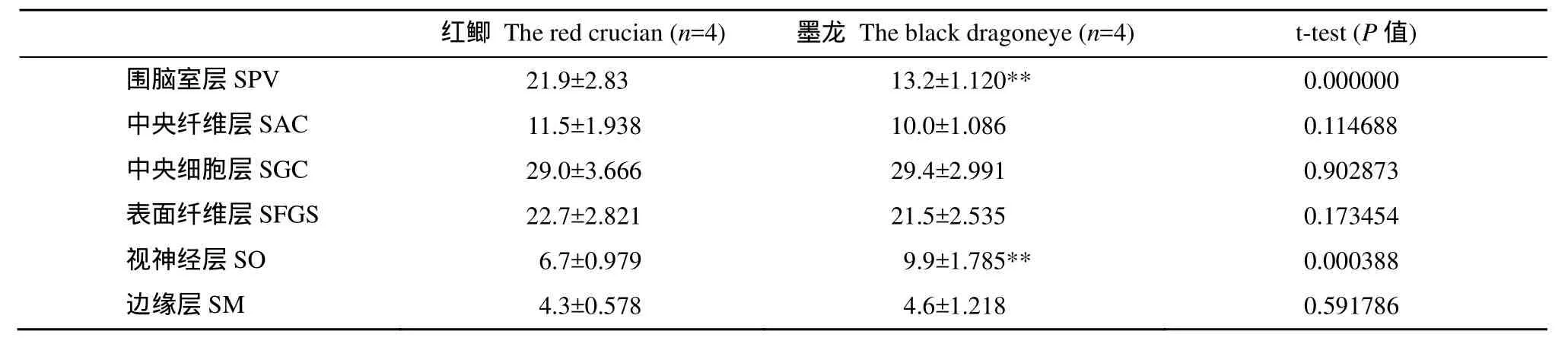
表5 红鲫与墨龙视盖各层与总厚度比值(相对厚度)比较Tab. 5 A comparison between the relative thickness of the optic tectum layers in black dragoneye and that red crucian (%)
可见与红鲫相比, 墨龙视网膜上感光细胞的减少, 造成对外界光线的敏感度和颜色分辨能力下降,但墨龙的眼球直径较大, 因此导致视网膜传入视盖的纤维增多, 并可能是导致其视网膜和视盖中的传递神经冲动的神经元、神经纤维所在层段增厚的主要原因。
与红鲫相比, 墨龙的视盖壁整体增厚28.9%,除围脑室层厚度减少22.6%外, 其余各层均明显变厚: (1)表面纤维层(SFGS)含有接受传入联系的神经元和神经纤维、视神经的前枝纤维,以及一些水平细胞和较大的锥体神经细胞, 视神经层(SO)含有来自中央和两侧视束的视神经纤维[3—8], 观察到表面纤维层(SFGS)的纤维分布远较红鲫密集, 且这两层均较红鲫增厚, 其中负责传入视觉神经冲动的视神经层(SO)厚度增达91.7%, 在视盖中相对厚度也较红鲫增大, 表明自视网膜输入视盖的神经纤维数量确实增多。(2)除含少数零星的水平细胞外, 主要由传出神经纤维组成的中央纤维层(SAC); 含有锥体神经细胞的树突和胞体位于中央细胞层(SGC)的上行神经纤维的边缘层(SM)[3—9]各层厚度均增加, 说明墨龙自视盖中发出的上行及下行神经纤维数量增加。(3)视盖的中央细胞层(SGC)内含有多极神经元、大梭状神经元和梨形神经元等多种类型的中间神经元[3—8], 墨龙的此层厚度增加说明其视盖各层间信息交换的能力增加。(4)墨龙围脑室层(SPV)由围脑室神经元和视盖深层纤维所组成, 神经元发出的神经纤维向外穿过视盖各层, 广泛地与视盖各层发生联系, 并可能是视觉信息传出的主要途径[3,7,9]。而墨龙的围脑室层的实际厚度和在视盖中的相对厚度较红鲫均显著减少, 表明其信息处理能力较红鲫差。此外由于视盖是鱼类的视觉中枢[3,6,9,12—14], 有将刺激从视网膜传递入脑形成视觉, 同时还具有控制鱼类位置和移动的功能, 墨龙的围脑室层厚度降低, 推测其游动及平衡能力也较差。
此外, 与前人观察结果相似[2], 也发现墨龙视网膜色素上皮层向杆锥层运动并交错对插, 推测是墨龙的色素上皮层可以防止因视网膜面积增大而导致杆锥层产生强烈光感。
总之, 人工选择不但改变了金鱼的外形, 而且使得金鱼的中枢神经组织结构也发生了较大变化,墨龙的眼径较红鲫大, 视网膜面积也较大, 导致传递神经冲动的神经纤维和神经元增多。推测墨龙视网膜色素上皮层向杆锥层运动并交错, 外核层、神经节细胞层厚度降低, 并且视盖中处理信息的围脑室层厚度也降低, 可以减少因视网膜面积增大而产生的强烈光感。
[1] Zhuang Y H, Zhou Y, Niu Y D, et al. Molecular phylogenetic relation ships of different varieties of goldfishes (Carssius auratus Var) and crucian carps (Carssius auratus) [J]. Acta Laser Biology Sinica, 2009, 18(1): 46—49 [庄远红, 周毅,牛艳东, 等. 不同品种金鱼和鲫鱼的分子系统发育关系研究. 激光生物学报, 2009, 18(1): 46—49]
[2] Ma D M, Zhu H P, Gui J F. Ectopic Six3 expression in the dragon eye goldfish [J]. Comparative Biochemistry and Physiology (Part B), 2008, 149(2): 303—313
[3] Lin H R. Physiology of Fish [M]. Guangzhou: Guangdong Higher Education Press. 1999, 247—275 [林浩然. 鱼类生理学. 广州: 广东高等教育出版社. 1999, 247—275]
[4] Estuardo R, Stephen J S, Herwig B. Characterization of genetically targeted neuron types in the zebrafish optic tectum [J]. Frontiers in Neural Circuits, 2011, 5: 1—14
[5] Chen R, Rao Y Z, Yang S J, et al. The histological study of the optic tectum in tilapia mossambica [J]. Journal of Zhanjiang Normal College, 2006, 27(6): 58—61 [陈蓉, 饶颖竹,杨素娇, 等. 莫桑比克罗非鱼中脑视顶盖组织学结构观察.湛江师范学院学报, 2006, 27(6): 58—61]
[6] Trygve E B, Charles F S. Visual system scaling in teleost fish [J]. The Journal of Comparative Neurology, 2012, 520(1): 142—153
[7] Kalman M. Astroglial architecture of the carp (Cyprinus carpio) brain as revealed by immunohistochemical staining against glial fibrillary acidic protein (GFAP) [J]. Anatomy and Embryology, 1998, 198(5): 409—433
[8] Barbara R, Heinrich R, Mario F W. The zebrafish brain: a neuroanatomical comparison with the goldfish [J]. Anatomy and Embryology, 1996, 194(2): 187—203
[9] Guerra A, Urbina M, Lima L. Modulation of taurine uptake in the goldfish retina and axonal transport to the tectum: effect of crushing the optic nerve or axotomy [J]. Amino Acids, 2000, 19(3—4): 687—703
[10] Ying M C. Ecology of Fish [M]. Beijing: Chinese Agricultural Press. 1995, 152—159 [殷名称. 鱼类生态学. 北京:中国农业出版社. 1995, 152—159]
[11] Wang D Q. The observation on the morphology and organization of the brain of eight freshwater cyprinids [J]. Journalof Fisheries of China, 1986, 10(1): 96—105 [王典群. 八种鲤科鱼类脑的形态构造观察, 水产学报, 1986, 10(1): 96—105]
[12] Wan A, Hua T M, Kong X J, et al. A morphology observation on the loach brain [J]. Journal of Biology, 2009, 26(6): 15—18 [万安, 华田苗, 孔学军, 等. 泥鳅脑的形态构造观察. 生物学杂志, 2009, 26(6): 15—18]
[13] Bing Z, Carp Organization [M]. Beijing: Science Press. 1983, 48—51 [秉志. 鲤鱼组织. 北京: 科学出版社. 1983, 48—51]
[14] Fang J, Fan J D, Chen Y. The immunocytochemical study on the distribution of 5-hydroxytryptamine in the brain of Schizothorax prenanti [J]. Acta Hydrobiologica Sinica, 2012. 36(1): 143—147 [方静, 樊均德, 玥陈. 齐口裂腹鱼脑内5-羟色胺免疫组织化学的定位观察, 水生生物学报, 2012. 36(1): 143—147]
A NEUROANATOMICAL STUDY OF THE BLACK DRAGONEYE’S RETINA AND OPTIC TECTUM IN COMPARISON WITH THE RED CRUCIAN’S
WAN An1,2, AN Shu-Qing1and HUA Tian-Miao3
(1. School of Life Science, Nanjing University, Nanjing 210093, China; 2. School of Resources and environment, Anqing Normal University, Anqing 246000, China; 3. School of Life Science, Anhui Normal University, Wuhu 241000, China)
The black dragoneye, a kind of dragoneye goldfish (Carassius auratus), has evolved from the red crucian. Four black dragoneyes and four red crucians, each being 10—12 cm long and weighing 35 g, were taken randomly and dissected. Their brains and eye-balls were observed using routine paraffin section and HE staining. The microscopic structures of the retinas and optic tectums of black dragoneye and the red crucian were studied under an optical microscope. We found that, compared with the red crucian, the retina of black dragoneye was 29.9% thinner, of which the outer plexiform layer increased 2.5%, and the inner plexiform layer increased 11.8%, while the inner nuclear layer decreased 21.6%, and the outer nuclear layer decreased 35.6%. Besides, the ganglion cell layer and the photoreceptor layer were thinner and irregular. However, optic tectum in black dragoneye was 28.9% thicker than that of the red crucian, except that the stratum periventriculare layer decreased 22.6%, all layers were incrassate, with the stratum album central layer increased 12.8%, the stratum griseum central layer increased 30.6%, the stratum fibrosum et griseum superficiale layer whose fibers were 21.9% denser than that of the red crusian, the stratum opticum layer was 91.7% thicker and the stratum marginal layer was 35.6% thicker. The results showed that the central nervous system of black dragoneye varied widely due to artificial selection and breeding. It can be inferred that the larger eyeball diameter and retina area of the black dragoneye resulted in an increase of fibers transmitted from the retina to the optic tectum, which probably was the main cause for the thicker layers of neurons and nerve fibers that transmit nervous impulse. To reduce the damage caused by strong light, the pigment epithelium layer moved to the photoreceptor layer interlacedly, and the inner nuclear layer of and outer nuclear layer the retina as well as the stratum periventriculare layer in its optic tectum became thinner. In addition, the thickness of the stratum periventriculare layer decreased, which can help to explain why the black dragoneye is not so good at swimming and balancing as the red crucian.
Carassius auratus; The black dragoneye; The red crucian; Retina; Optic tectum
Q111.2+2
A
1000-3207(2013)01-0100-06
10.7541/2013.100
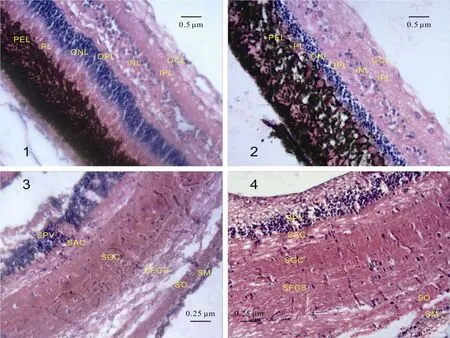
图版Ⅰ Plate Ⅰ
1.红鲫视网膜; 2. 墨龙视网膜; 3. 红鲫视盖; 4. 墨龙视盖; 如图1, 2所示, 红鲫视网膜结构清晰, 而墨龙视网膜的色素上皮层(PEL)向杆锥层(PL)运动, 并交错对插, 与红鲫相比, 墨龙视网膜外核层(ONL)、内核层(INL)厚度降低, 外网状层(OPL)和内网状层(IPL)厚度增加;如图3, 4所示, 红鲫和墨龙的视盖各层结构均清晰可辨, 与红鲫相比, 墨龙围脑室层(SPV)厚度降低, 其余各层厚度均有所增加
1. The Black Dragoneye’s Retina; 2. The Black Dragoneye’s Retina; 3. The Red Crucian’s Optic Tectum; 4. The Black Dragoneye’s Optic Tectum. As Picture 1 and Picture 2 show, the retina of red crucian presents a legible structure, but in the retina of black dragoneye the pigment epithelium layer (PEL) moves to the photoreceptor layer (PL) interlacedly. Besides, the inner nuclear layer (INL) and outer nuclear layer (ONL) of black dragoneye were thinner, while its outer plexiform layer (OPL) and inner plexiform layer (IPL) were thicker; As Picture 3 and Picture 4 show, the optic tectum of red crucian and black dragoneye are legible. Compared with the red crucian, the stratum periventriculare (SPV) layer in optic tectum was thinner in the black dragoneye, and other layers were thicker. PEL: 色素上皮层Pigment epithelium layer; PL: 杆锥层Photoreceptor layer; ONL: 外核层Outer nuclear layer; OPL:外网状层Outer plexiform layer; INL: 内核层Inner nuclear layer; IPL: 内网状层Inner plexiform layer; GCL: 神经节细胞层Ganglion cell layer; SPV: 围脑室层Stratum periventriculare; SAC: 中央纤维层Stratum album central; SGC: 中央细胞层Stratum griseum central; SFGS: 表面纤维层Stratum fibrosum et griseum superficiale; SO: 视神经层Stratum opticum; SM: 边缘层Stratum marginal
2011-12-27;
2012-10-17
国家自然科学基金(31171082); 南京大学重点学科经费; 安庆师范学院硕士科研启动基金(No.044-K05000000195); 校级教研项目(No.044-J10004000024)资助
万安(1980—), 男, 安徽安庆人; 博士研究生; 研究方向为生态学。E-mail: wanan@aqtc.edu.cn
安树青(1963—), E-mail: anshq@nju.edu.cn
