班氏跳小蜂个体发育研究
2012-11-22许再福
何 佳,许再福
(华南农业大学昆虫学系,广州510642)
扶桑绵粉蚧Phenacoccus solenopsis Tinsley是一种危害园林、蔬菜、大田作物的害虫,对棉花等多种作物的生产构成威胁,是我国重要检疫对象(Hodgson et al.,2008;朱艺勇等,2011)。班氏跳小蜂Aenasius bambawalei Hayat属膜翅目跳小蜂科Encyrtidae,是扶桑绵粉蚧的重要寄生性天敌,是一种内寄生蜂,于2008年首次在印度被发现,主要分布于中国 (湖南、广东、海南)、印度和巴基斯坦 (Tanwar et al.,2008;Hayat,2009;陈华燕等,2010;黄玲等,2011)。班氏跳小蜂对不同寄主上的扶桑绵粉蚧均有较高自然寄生率,即使在扶桑绵粉蚧发生水平较低时,仍能有较高的寄生率 (Tanwar et al.,2008;Dhawan A.K.et al.,2011)。目前国内外对班氏跳小蜂的生物学研究较少,主要是关于寄主龄期选择、发育起点温度、发育历期等方面内容,很少有关该蜂的个体发育研究报道 (黄玲等,2011;Poorani et al.,2011;Zain-ul-Abdin et al.,2012)。为了更好地繁殖和利用该寄生蜂,我们在实验室条件下系统观察了班氏跳小蜂的个体发育情况。现将结果报道如下。
1 材料与方法
1.1 试验材料
扶桑绵粉蚧Phenacoccus solenopsis Tinsley用马铃薯Solanum tuberosum幼苗来培养繁殖。
班氏跳小蜂Aenasius bambawalei Hayat以扶桑绵粉蚧雌成虫为寄主进行繁殖。用规格为15 mm×100 mm的玻璃试管收集并保存蜂,并用20%的蜂蜜水给蜂补充营养。
以上试验均在温度27±1℃,相对湿度70±5%,光周期L12∶D12的人工气候箱中进行。
1.2 试验方法
选取12株长势相同并分别接有100头扶桑绵粉蚧雌成虫的马铃薯幼苗放入繁蜂箱 (长40cm×宽40 cm×高40 cm)中,接入650头已交配的雌性班氏跳小蜂,寄生12 h后,取出带有粉蚧的马铃薯幼苗。将带有扶桑绵粉蚧的马铃薯幼苗放在温度27±1℃,相对湿度 70±5%,光周期为L12∶D12的人工气候箱中继续培养。每隔12 h解剖一次,每次解剖被寄生的扶桑绵粉蚧雌成虫30头,直至羽化。在体视镜下观察被寄生的粉蚧雌成虫体色以及外部形态特征的变化,记录寄生蜂各虫态的形态特征及发育情况;用测微尺测量寄生蜂虫体大小;选择典型虫态进行显微拍照。
2 结果与分析
2.1 班氏跳小蜂的发育历期
在试验条件下,班氏跳小蜂从卵开始,发育到成虫,整个过程均在扶桑绵粉蚧体内完成,其发育经历卵、幼虫、蛹和成虫4个阶段,其历期为14.5 d。其中,卵的发育历期为1.5 d,1龄幼虫历期1 d,2龄幼虫历期1 d,3龄幼虫历期为2 d,4龄幼虫历期为2.5 d,预蛹的历期为1 d,蛹的历期为5.5 d。
2.2 班氏跳小蜂的形态特征
卵:卵 (图1)为辣椒形,长0.19~0.27 mm,宽0.10~0.13 mm;卵壳褐色,有细长卵柄;卵柄末端颜色较浅,弧形弯曲,突出于寄主体壁 (图2),起到呼吸的作用。卵壳与卵柄相连处颜色较深。初产的卵卵体小、半透明,大部分被卵壳包被;随着卵裂的进行,卵体不断增大,显著突出于卵壳外,此时剥除卵壳可见卵为椭圆形。
幼虫:幼虫分4龄。1龄幼虫 (图3)体长0.23~0.32 mm,宽0.13~0.17 mm;体透明,末端组织紧连卵壳;体躯分为头部 (1节)和胸腹部 (7节)共8个体节,分节从腹面易识别,背面不明显。初孵幼虫体内可见淡黄色卵圆形的内容物,发育到1龄后期,内容物颜色变深。2龄幼虫(图4)体长0.51~0.60 mm,宽0.18~0.23 mm;体透明,尾端连接卵壳;头部和体躯相接处缢缩,头部明显可辨,可见黄色硬化的上颚;胸部3节,腹部11节;体躯内部消化道清晰可见,占据较大体积;体侧有白色的气管纵干和侧支出现,连接卵壳。3龄幼虫体长1.58~1.84 mm,宽0.47~0.65 mm。体半透明,浅乳白色,尾端与卵壳相连;体分节明显,分为头部、胸部 (3节)和腹部 (11节);腹末腹面可见1对黄色的排泄组织;体侧可见明显的气管纵干、侧支和分支气管,气管纵干与卵壳相连;虫体可在寄主体内的一定范围移动。4龄幼虫体长2.50~3.17 mm,宽1.00~1.33 mm;体粗壮,半头型,上颚硬化程度高;4龄幼虫 (图5)初期体躯半透明,分为头部、胸部(3节)和腹部 (11节);腹末气管连接卵壳,但随着气管系统的逐渐完善,虫体渐渐脱离卵壳直至腹末钝圆 (图6);后期气管系统已遍及整个幼虫体躯,4龄后期寄主的组织已经消耗殆尽,寄主体壁开始硬化,逐渐形成僵蚧。
预蛹:预蛹 (图7)体长1.79~2.17 mm,宽0.96~1.08 mm。体乳白色,半透明,口器褐黄色;消化道内的食物已经排泄完毕;体明显缩短,但仍可分为头部、胸部和腹部;胸部三节发达,较宽,约占体长的一半;腹部体节紧凑,分节不明显。
蛹:体长2.00~2.32 mm,宽1.06~1.29 mm。初期蛹体白色透明,但复眼、触角和足等清晰可辨 (图8)。根据触角类型可以辨别其雌雄两性。随着发育进行,体色渐深。首先是复眼的颜色发生变化,接着是胸部和腹部的颜色变化;复眼由浅黄色到褐黄色、褐色、黑褐色变化;伴随着复眼颜色的变化,胸部和腹部的体色也逐渐变为黑色;胸部由背面中央变为黑色,逐渐向胸部侧面和腹面发展;腹部由前端背面中央逐渐向腹部侧面、后端和腹面变黑;直到胸部和腹部全部变为黑色 (图9)。
成虫:雌蜂 (图10)体黑色,体长1.62~2.23 mm。触角棒状。复眼大,卵圆形,具稀疏的白毛。前翅从前翅基部到无毛斜带外缘的区域为褐色,其余部分无色透明;后翅无色透明。小盾片大,呈U字形。足黑色,但胫节端部和第1~4跗节为黄褐色。腹部稍短于胸部。产卵器伸过腹末。雄蜂 (图11)体长0.83~1.37 mm。触角黑褐色;索节短而横宽;棒节长且不分节,中部稍弯,密被绒毛;前翅和后翅均无色透明。
2.3 寄主的形态变化
自班氏跳小蜂在寄主体上产卵至其幼虫发育至3龄期间,被寄生的扶桑绵粉蚧雌成虫外部形态未见出现明显变化。但当粉蚧被寄生5.5 d后,班氏跳小蜂幼虫发育开始进入4龄时,粉蚧出现行动迟缓,体色加深,胸部和腹部背面黑斑开始褪去,体背逐渐隆起,体形开始由扁平变成半椭球形;当粉蚧被寄生6.5 d后,外部已无头部、胸部与腹部的分段,变成被有蜡粉的半椭球形僵蚧,粘附于寄主植物表面,此时僵蚧体壁较软,体内物质尚未被完全消耗;当粉蚧被寄生7 d后,跳小蜂进入4龄幼虫发育后期,僵蚧体壁变硬;当粉蚧被寄生8 d后,跳小蜂完成卵和幼虫的发育,粉蚧体内物质被消耗殆尽,蜡粉依然粘附于体壁上,但可被轻易拭去,露出褐色光滑的体壁,体壁半透明,可见内部正在发育的蛹 (图12)。
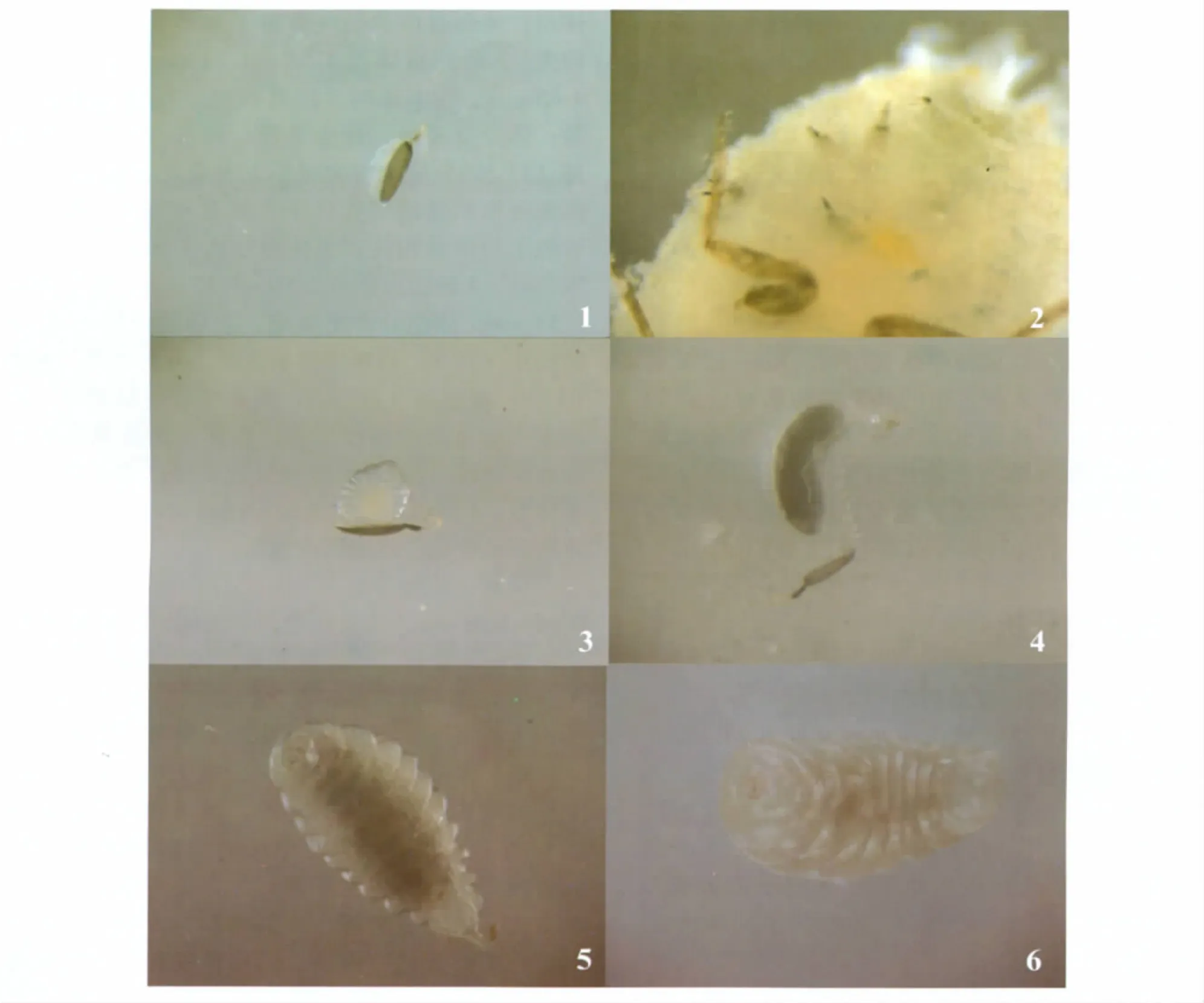
图版I 班氏跳小蜂Plate I Aenasius bambawalei Hayat

图版II 班氏跳小蜂及僵蚧PlateⅡ Aenasius bambawalei Hayat and mummy
3 结论与讨论
Poorani等 (2009)曾报道在温度27±5℃,相对湿度65%的实验条件下,以2~3龄扶桑绵粉蚧幼虫为寄主,班氏跳小蜂在寄主体内从卵发育到蛹的历期、蛹发育到成虫的历期以及生命周期分别为8.85 d、7.35 d和16.10 d,其研究结果比本实验室的研究结果分别推后0.85 d、0.85 d和1.6 d。这种差异的产生可能与实验温度和寄主龄期的不同有关,相对于扶桑绵粉蚧幼虫,成虫可提供更多班氏跳小蜂发育所需的营养物质。
班氏跳小蜂的卵和幼虫均需要通过卵柄与外界进行气体交换,因此卵柄末端突出于寄主体外。幼虫在寄主体内依靠卵柄与外界进行气体交换直至4龄后期。在这过程中,幼虫与卵壳的连接组织逐渐萎缩,气管系统逐渐发达直至最后遍及躯体。目前已研究过的跳小蜂Diaphorencyrtus aligarhensis和阔柄跳小蜂Metaphycus flavus等均无卵柄这一构造,与班氏跳小蜂幼虫的呼吸方式截然不同。
班氏跳小蜂在1头寄主体内可产卵多达5粒,平均2粒。但是,1头被寄生的扶桑绵粉蚧体内最多只能羽化出1头班氏跳小蜂。这表明班氏跳小蜂有过寄生现象。因此,在同一寄主体内的班氏跳小蜂个体间存在竞争。Fisher等 (1961)指出,过寄生造成寄生蜂子代部分个体死亡的原因主要有寄主资源不足、物理性竞争、生理性抑制等3种。Tena等 (2011)发现,阔柄跳小蜂Metaphycus flavus在4龄幼虫时通过发达的上颚来咬杀同一寄主体内的其他个体,进行种内竞争,直至最后留下1头幼虫。通常情况下,产于扶桑绵粉蚧体内的卵都能顺利孵化,但是同一寄主体内只有1头幼虫能够进入2龄或3龄,其余个体则在1龄或2龄时就被竞争力强的幼虫完全吸收。由此可见,班氏跳小蜂种内竞争出现于上颚尚未高度硬化的2龄幼虫前,此时的上颚尚未具有咬杀的功能。因此,难以用物理性种内竞争来解释。Vinson等 (1998)认为,先发育的寄生蜂会改变寄主血淋巴成分,改变后的血淋巴会抑制其他寄生蜂的发育。班氏跳小蜂是否通过生理性抑制的机制来进行种内调节,有待进一步的研究。
References)
Chen HY,Cao RX and Xu ZF,2010.First record of Aenasius bambawalei Hayat(Hymenoptera:Encyrtidae),a parasitoid of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)from China.Journal of Environmental Entomology,32(2):280 - 282.[陈华燕,曹润欣,许再福,2010.扶桑绵粉蚧寄生蜂优势种Aenasius bambawalei Hayat记述.环境昆虫学报,32(2):280-282]
Dhawan AK,Kamaldeep S,Sarika S,Anand A,Jasbir S,2011.Parasitizing potential of parasitoid (Aenasius bambawalei)on mealybug(Phenacoccus solenopsis)in cotton(Gossypium spp.)and weed plants.Indian Journal of Agricultural Sciences,81(1):97-99.
Fisher RC,1961.A study in insect multiparasitism.Ⅱ.The mechanism and control of competition for possession of the host.Journal of Experimental Biology,38:605-628.
Hayat M,2009.Description of a new species of Aenasius Walker(Hymenoptera:Encyrtidae), parasitoid of the mealybug,Phenacoccus solenopsis Tinsley(Homoptera Pseudococcidae)in India.Biosystematica,3(1):21-26.
Hodgson C,Abbas G,Arief MJ,Saeed S,Karar H,2008.Phenacoccus solenopsis Tinsley(Sternorrhyncha:Coccoidea Pseudococcidae),an invasive mealybug damaging cotton in Pakistan and India,with a discussion on seasonal morphological variation.Zootaxa,1913:1-35.
Huang L,Liu H,Xiao GC,Zhou SW,2011.Biology of Aenasius bambawalei.Chinese Journal of Applied Entomology,48(6):1734-1737.[黄玲,刘慧,肖铁光,周社文,2011.扶桑绵粉蚧寄生性天敌班氏跳小蜂生物学研究.应用昆虫学报,48(6):1734-1737]
Poorani J,Rajeshwari SK,Gupta A,2009.Notes on diagnosis and biology of Aenasius bambawalei Hayat(Hymenoptera:Encyrtidae),a parasitoid of the invasive mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Sternorrhyncha:Pseudococcidae).Journal of Biological Control,23(4):463 -466.
Rohrig E,Shirk PD,Hall DG,Stansly PA,2011.Larval development of Diaphorencyrtus aligarhensis (Hymenoptera:Encyridae),an endoparasitoid of Diaphorina citri(Hemiptera:Psyllidae).Arthropod Biology,104(1):50-58.
Tanwar RK,Bhamare VK,Ramamurthy VV,Hayat M,Jeyakumar P,Singh A,Bambawale OM,2008.Record of new parasitoids on mealybug,Phenacoccus solenopsis.Indian Journal of Entomology,70(4):404-405.
Tena A,Kapranas A,Walker GP,Garcia-MaríF,Luck RF,2011.Larval morphology of Metaphycus flavus and its role in host attachment and larval cannibalism.Bulletin of Entomological Research,101:365-372.
Vinson SB,Hegazi EM,1998.A possible mechanism for the physiological suppression of conspecific eggs and larvae following superparasitism by solitary endoparasitoids.Journal of Insect Physiology,44:703 -712.
Zainul-Abdin AMJ,Gogi MD,Muhammad A,Fiaz H,Abbas SK,Hoor S,Atif M,2012.Biological characteristics and host stage preference of mealybug parasitoid Aenasius bambawalei Hayat(Hymenoptera:Encyrtidae).Pakistan Entomologist,34(1):47 -50.
Zhu YY,Huang F,Lv YB,2011.Bionomics of mealybug Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)on cotton.Acta Entomologica Sinica,54(2):246-252.[朱艺勇,黄芳,吕要斌,2011.扶桑绵粉蚧生物学特性研究.昆虫学报,54(2):246-252]
