CR1基因多态性与汉族人迟发性阿尔茨海默病关系
2012-11-21田燕卢瑞春郁金泰谭兰
田燕,卢瑞春,郁金泰,谭兰
(青岛市市立医院神经内科,山东 青岛 266071)
阿尔茨海默病(AD)是一种病因不明的进行性神经系统变性疾病,根据发病年龄(以65岁为界)分为早发性AD和迟发性AD(LOAD)。AD的发病与遗传变异有关的观点已经得到广泛认可[1]。载脂蛋白E(Apo E)ε4等位基因是目前已经被证实的AD的独立危险因素之一,但携带此等位基因的病人仅占38%[2]。因此,发现更多的遗传基因标记均对诊断和治疗AD是非常有必要的。淀粉样β蛋白诱导补体系统激活在AD发病中发挥重要作用。补体受体1(CR1)被认为有助于清除淀粉样β蛋白。最近有研究结果表明,CR1的基因多态性是否与高加索人的LOAD有关[3]。但是,CR1基因多态性与汉族人LOAD有关尚需研究证实。本研究旨在探讨CR1基因多态性与LOAD发病的关系。
1 对象与方法
1.1 研究对象
LOAD组254例为2006年9月—2010年10月我科的住院痴呆病人,其中男118例,女136例;平均年龄(78.4±6.7)岁;发病年龄为(72.6±6.8)岁。LOAD诊断均符合美国国立神经病、语言机能紊乱和卒中研究所及阿尔茨海默病和相关疾病协会(NINCDS-ADRDA)的标准。均排除了其他神经系统退行性疾病,无AD及精神病家族遗传史。对照组357例为来自我院体检门诊的健康人,其中男161例,女196例;平均年龄(78.7±6.4)岁。均无记忆和智能障碍,MMSE评分>28;临床及实验室检查均正常;均无自身免疫疾病、糖尿病、心肌梗死、哮喘及脑卒中病史。两组均为中国北方汉族人。此项研究获得研究对象的同意及伦理委员会批准。
1.2 研究方法
1.2.1 基因提取 采集外周血5 mL,ACD抗凝,采用苯酚-氯仿法提取基因。
1.2.2 CR1基因多态性检测 引物的设计和合成由上海生工公司完成。rs6656401引物序列:上游引物,5′-ACGTTGGATGGCTTGTAGATGCATCATTTCC-3′;下游引物,5′-ACGTTGGATGGACAGAAGAGCAAAGGACAC-3′;扩增引物,5′-ACACACAGAGGAGAAGGCGA-3′。rs3818361引物序列:上游引物,5′-ACGTTGGATGAAAGGACAGTTCCAGAGCAC-3′;下游引物,5′-ACGTTGGATGTTTTAAGCCCTCTGGTAAGC-3′;扩增引物,5′-CCTCTGGTAAGCATAAGATATA-3′。PCR参数为95℃15 min;95℃5 s、56℃20 s、72℃30 s,共50个循环;72℃30 s[4]。
1.2.3 Apo E基因型检测 引物序列为:5′-ACAGAATTCGCCCCGGCCT GGTACAC-3′和5′-TAAGCTTGGCACGGCTGTCCAAGGA-3′。PCR 参数为95 ℃ 5 min;95 ℃ 30 s、62 ℃ 30 s、70 ℃1 min,共30个循环;70℃5 min[5]。非变性聚丙烯酰胺凝胶电泳分离酶切产物,银染后Gel-Doc 2000下照相分型。
1.3 统计学处理
用H WJ软件进行Hardy-Weinberg平衡估计,其他统计分析用SPSS 11.5软件完成。用直接计数法计算等位基因和等位基因型频率,各组间基因型和等位基因频率比较采用χ2检验。
2 结 果
2.1 各基因组Hardy-Weinberg平衡吻合度检测
除LOAD组rs6656401以外,对照组和LOAD组病人中其他SNP位点的基因型分布均符合Hardy-Weinber g平衡法则(P>0.05)。
2.2 两组临床特征比较
LOAD组和对照组的年龄、性别无明显差异(P>0.05)。LOAD 组携带 Apo Eε4等位基因者120例,对照组携带Apo Eε4等位基因者84例,携带Apo Eε4等位基因可增加LOAD的发病风险(χ2=5.52,P<0.05,OR=2.91,95%CI=2.06~4.12)。
2.3 两组rs6656401和Apo E等位基因和基因型频率分布
LOAD组与对照组CR1基因rs6656401的基因型和等位基因频率差异有显著意义(χ2=6.56、7.16,P<0.05),等位基因A显著增加了LOAD的发病风险(OR=2.60,95%CI=1.13~5.98)。这一显著性差异在非Apo Eε4携带者中依然存在(χ2=11.78、11.58,P<0.05),而在 Apo Eε4携带者中则未发现两组间rs6656401基因型和等位基因频率差异有显著性(P>0.05)。见表1~3。
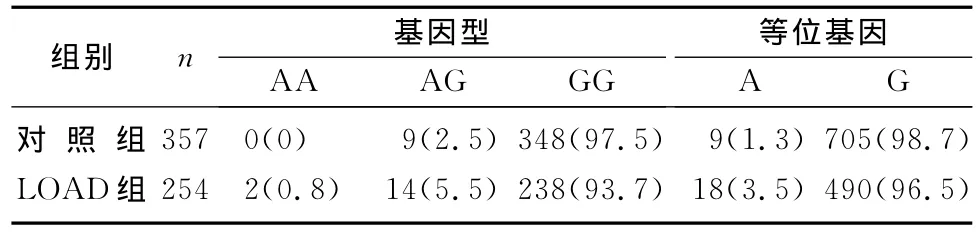
表1 两组rs6656401等位基因和基因型频率分布(例(χ/%))

表2 两组ApoEε4携带者rs6656401等位基因和基因型频率比较(例(χ/%))

表3 两组非ApoEε4携带者rs6656401等位基因和基因型频率比较(例(χ/%))
2.4 两组CR1基因rs3818361基因型和等位基因频率分布
LOAD组与对照组CR1基因rs3818361基因型和等位基因频率差异无显著性(P>0.05)。然而,将该位点经是否携带Apo Eε4分层后,发现非Apo Eε4携带者中两组等位基因频率差异有显著性(χ2=4.45,P<0.05,OR=1.38,95%CI=1.02~1.87)。见表4~6。

表4 两组CR1基因rs3818361基因型和等位基因频率比较(例(χ/%))
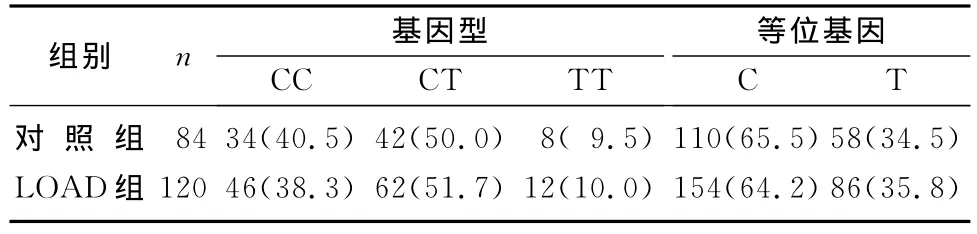
表5 两组ApoEε4携带者CR1基因rs3818361基因型和等位基因频率比较(例(χ/%))
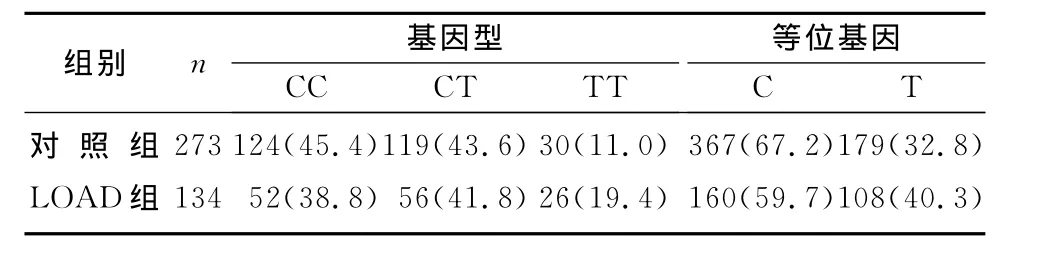
表6 两组非ApoEε4携带者CR1基因rs3818361基因型和等位基因频率比较(例(χ/%))
2.5 LOAD多因素Logistic回归分析
经过性别、年龄和Apo E调整以后,CR1基因rs6656401等位基因A携带者(基因型AA和AG)较基因型GG纯合子导致LOAD的风险高2.40倍(OR=2.40,95%CI=1.00-5.73)。而rs3818361基因型TT和CT与CC间的差异则无显著性(P>0.05)。
2.6 对这两个位点可能组合的单倍型分析
AT单倍型在LOAD组出现的频率高于对照组(χ2=4.70,P<0.05,OR=2.44,95%CI=1.06~5.62)。见表7。

表7 两组单倍型频率比较(例(χ/%))
3 讨 论
3.1 Apo E基因与LOAD
业已证实,Apo Eε4等位基因被认为是LOAD发病的易感基因[6],本研究结果也证实了这一点。但是,Apo Eε4等位基因仅占LOAD发病独立危险因素的40%~50%[7]。国内外的研究不仅证实此等位基因是LOAD的遗传易感基因,而且证实其与其他一些遗传易感基因存在相互作用[8]。目前,Apo Eε4等位基因已经成为LOAD遗传研究的基本参量[9]。
3.2 CR1基因与LOAD
Aβ是一种异质的分子结构,能够激活补体系统并与C3b结合。曾经有研究认为,β蛋白诱导补体激活是AD的重要发病机制[10]。已经有研究发现转基因鼠过度表达C3能减少Aβ的沉积和病理作用[11]。相反,抑制C3转化酶的表达则加速Aβ的沉积和神经退行性改变[12]。Aβ通过C3b介导与红细胞表面的CR1相结合,并最终被红细胞清除出循环系统。而CR1作为补体的调节剂主要存在于外周血细胞的细胞膜中。CR1还存在于外周神经有髓鞘纤维的施万细胞内,并推测其通过限制补体激活而阻止有髓鞘纤维脱髓鞘改变[13]。有研究发现,可溶性的CR1具有治疗自身免疫性疾病和炎症性疾病的作用。因此,我们推测CR1在AD发病中具有保护作用。
虽然本研究中rs3818361和rs6656401位点均位于CR1的内含子中不能直接参与调整蛋白的表达,但是此内含子基因的突变可能通过影响CR1基因的表达而导致LOAD的发病。
最近 LA MBERT 等[3]的研究结果显示,CR1的基因多态性是高加索人患AD的重要基因危险因素,rs6656401和rs3818361的等位基因是LOAD发病的危险因素,尤其是rs6656401。另外一项基因组研究显示,rs3818361是欧洲和美洲人群发生AD的危险因素[14]。本研究结果显示,rs6656401基因多态性与LOAD发病有关,rs6656401的A等位基因与G等位基因联合能增加LOAD的发病风险,此结论与LA MBERT等的研究结论相一致。但本研究未显示rs3818361的等位基因与LOAD发病有关。LOAD组与对照组CR1基因rs3818361基因型和等位基因频率分布也没有显著性差异。用是否携带Apo Eε4分层后,发现非Apo Eε4携带者中LOAD组rs6656401的A等位基因和rs3818361的T等位基因出现的频率较对照组高。这些情况均与LA MBERT等的研究结果不同,考虑可能与种族不同有关。本研究结果还表明,AT单倍型是LOAD发病的危险因素。
综上所述,CR1基因突变可能通过影响Aβ的清除而导致LOAD发病,rs6656401的A等位基因是汉族人患LOAD的重要危险因素。
[1]BERTRA M L,TANZI R E.Thirty years of Alzhei mer’s disease genetics:the i mplications of systematic meta-analyses[J].Nat Rev Neurosci,2008,9:768-778.
[2]BERTRA M L,MCQUEEN M B,MULLIN K,et al.Association studies:the Alz Gene database[J].Nat Genet,2007,39:17-23.
[3]LAMBERT J C,HEATH S,EVEN G, et al.Geno me-wide association study identifies variants at CLU and CR1 associated with Alzhei mer’s disease[J].Nat Genet,2009,41:1094-1099.
[4]BUETOW K H,EDMONSON M.High-t hroughput development and characterization of a geno mewide collection of genebased single nucleotide poly mor phis m mar kers by chip-based matrix-assisted laser desor ption/ionization time-of-flight mass spectro metry[J].Natl Acad Sci USA,2001,98(2):581-584.
[5]DONOHOE,SALOMKI,LEHTI MKI,et al.Rapid identification of Apolipopr otein E genotypes by multiplex amplification refractory mutation system PCR and capillary gel electr ophoresis[J].Clin Chem,1999,45:143-146.
[6]WILLIA MSON J,GOLDMAN J,MARDER K S,et al.Genetic aspects of Alzhei mer disease[J].Neur ologist,2009,15(2):80-86.
[7]BERTRA M L,TANZI R E.Thirty years of Alzhei mer’s disease genetics:the i mplications of systematic meta-analyses[J].Nat Rev Neur osci,2008,9:768-778.
[8]YU J T,TAN L.Poly mor phisms at theβ2-adrenergic receptor gene infl uence Alzhei mer’s disease susceptibility[J].Brain Research,2008,1210:216-222.
[9]韩彬,张生林.Apo E、A2 M、ACE基因与汉人Alzhei mer病的相关性研究[J].山西医科大学学报,2008,39(8):692-697.
[10]BONIFATI D M,KISHORE U.Role of co mplement in neur odegeneration and neur oinflammation[J].Mol Immunol,2007,44:999-1010.
[11]ZHOU J,FONSECA M I,PISALYAPUT K,et al.Co mplement C3 and C4 expression in C1q sufficient and deficient mouse models of Alzhei mer’s disease[J].Neur ochem,2008,106:2080-2092.
[12]WYSS-CORAY T,YAN F,LIN A HT,et al.Pro minent neurodegeneration and increased plaque for mation in co mplementinhibited Alzhei mer’s mice[J].Proc Natl Acad Sci USA,2002,99:10837-10842.
[13]VEDELER C A,MATRE R.Peripheral ner ve CR1 express in sit u cofactor activity for degradation of C3b[J].Neuroi mmunol,1990,26:51-56.
[14]HAROLD D,ABRAHA M R,HOLLINGWORT H P J.Geno me-wide association study identifies variants at CLU and PICAL M associated with Alzhei mer’s disease[J].Nat Genet,2009,41:1088-1093.
