水曲柳体胚发生过程中不同状态类型外植体的生理生化状态
2012-11-10丛建民沈海龙李玉花
丛建民,沈海龙,李玉花,张 鹏,杨 玲,黄 剑
(1 东北林业大学生命科学学院,黑龙江哈尔滨 150040;2 白城师范学院生物系,吉林白城 137000;3 东北林业大学林学院,黑龙江哈尔滨 150040;4东北林业大学,林木遗传育种国家重点实验室,黑龙江哈尔滨 150040)
植物的体细胞胚胎发生(Somatic embryogenesis)具有普遍性、两极性、遗传稳定性等特点,通过生产体胚苗可以为优良品系大规模快速无性繁殖和建立植物生物学研究提供高效试验体系[1].水曲柳Fraxinus mandshurica属于木犀科白蜡树属大乔木用材和观赏树种.建立其体胚发生体系,对快速扩繁其优良品系和进行其发育生物学、遗传转化研究均具有重要意义.我们的系列研究[2-8]证明,水曲柳体胚发生最适宜的外植体材料是刚进入子叶期的幼嫩合子胚的子叶;但是在水曲柳体胚诱导过程中,体胚的发生基本上都伴随着外植体的褐化,而且幼胚子叶外植体表现为多种状态类型,如褐化发生体胚、未褐化发生体胚等.体胚发生过程中外植体不同的状态类型是其各种内在的生理生化反应和基因表达调控的外在综合体现,发掘和认识这种内在变化与外在表观类型之间的内在联系,可以为解析体胚发生机理和改善体胚发生技术提供科学依据.本研究针对水曲柳体胚发生过程中各种状态外植体的一些生理生化指标进行检测,以期从生理生化角度对各种状态类型外植体与水曲柳体胚发生的关系进行解析,为提高水曲柳体胚发生技术提供理论依据,并为木本植物体胚发生的研究提供一个可能的新的切入点.
1 材料与方法
1.1 不同状态类型外植体材料的获得
水曲柳体胚发生的外植体采集和体胚诱导等按照本研究组前期研究确定的成熟方案[2-8]进行.诱导和继代培养基为MS1/2(将MS培养基所有成分都减半),添加水解酪蛋白(CH)400 mg·L-1,蔗糖70 g·L-1,BA 0.5 mg·L-1,NAA 1.5 mg·L-1,琼脂 6 g·L-1,添加琼脂前调节pH至5.8,然后120℃高压灭菌20 min.培养室的温度控制在25℃,湿度60% ~70%之间.
2009年7月14日,在东北林业大学实验林场内,从不同母树上采取水曲柳未成熟种子(受粉后9周左右,子叶形胚初期).将种子去翅,置于蒸馏水中浸泡12 h后,再在流水下冲洗2 h;在超净工作台中用φ为70%的乙醇处理10 s,20 mg·L-1的次氯酸钠消毒10 min,用无菌水冲洗4~5次.将种子胚根端切去1/3~1/2后挤出幼胚,取单片子叶接种到诱导培养基上(子叶内侧附于培养基上),每皿接种10个外植体.接种后的培养皿放入纸箱中暗培养.4周后将外植体转接到相同新鲜培养基上.培养45 d时取不同状态类型外植体用作生理生化检测试验材料.
外植体状态类型包括:褐化外植体发生体胚(HT)、未褐化外植体发生体胚(WT)、褐化外植体发生愈伤组织(HY)、未褐化外植体发生愈伤组织(WY)、褐化外植体长大(HD)、未褐化外植体长大(WD)、褐化外植体未长大(HW)、未褐化外植体未长大(WW).
1.2 生理生化指标测定方法
可溶性多糖含量的测定用蒽酮法[9].可溶性蛋白含量测定用考马斯亮兰G-250法[10],以牛血清蛋白做标准曲线,计算各样品的蛋白质质量比(mg·g-1).超氧化物歧化酶(SOD)活性测定用氯化硝基四氮哇蓝法[11].过氧化物酶(POD)活性测定用 Kochba法[12].过氧化氢酶(CAT)活性测定采用过氧化氢还原法,参考陈建勋[13]方法进行测定,多酚氧化酶(PPO)活性参照罗晓芳等[14]的方法测定.均重复3次,取平均值.
运用Spss 10.0软件对数据进行差异显著性分析,Excel软件作图.
2 结果与分析
2.1 水曲柳体胚发生的合子胚子叶外植体总体变化
培养中观察水曲柳合子胚子叶外植体状态,在3~5 d微增大,颜色变深,表面有褐色斑形成,10 d左右在外植体边缘或切口处有大量的愈伤组织产生,有少量的体胚出现在外植体褐化部位,这时的体胚多数处于球形胚状态.20 d开始,愈伤组织明显长大,而体胚的数量也开始显著增加.培养30 d后,有些愈伤组织呈现出粘稠状非胚性愈伤组织(即使是愈伤组织的状态,继续培养也不会出现体胚),有一些愈伤组织呈现疏松颗粒状,为胚性愈伤组织(经过继续培养,会有体胚出现).褐化的外植体占总状态的78.67%,体胚发生率为11.36%.其中,褐化外植体上体胚(HT)发生率为8.15%,明显高于未褐化外植体上体胚(WT)发生率3.21%的水平.
2.2 不同状态类型外植体的多糖含量变化
由图1A可以看出,水曲柳体胚发生过程中,不同状态类型外植体的多糖含量之间存在显著差异(P<0.05).未褐化未长大(WW)外植体中多糖含量最高,褐化长大(HD)外植体次高,褐化发生体胚(HT)外植体中多糖含量最低,其他长大或发生愈伤组织外植体居中.这个格局表明,外植体长大、褐化、发生愈伤组织和发生体胚等均消耗组织内的多糖,其中以褐化并发生体胚的外植体多糖消耗的数量最多.说明在水曲柳体胚发生过程中,外植体内部的多糖类物质被降解、转化、代谢,以便为生长分化提供能量.需要注意的是外植体褐化本身也会消耗多糖.
2.3 不同状态类型外植体的可溶性蛋白含量变化
图1B结果表明,不同状态类型的外植体的可溶性蛋白含量之间存在着显著差异(P<0.05).未褐化发生体胚(WT)外植体的可溶性蛋白含量最高,其次为未褐化长大(WD)和未褐化发生愈伤组织(WY)外植体.而褐化未长大(HW)、褐化长大(HD)、褐化发生体胚(HT)和褐化发生愈伤组织(HY)的外植体中可溶性蛋白的含量明显低于前三者未褐化的外植体.但是,未褐化未长大(WW)的外植体中可溶性蛋白的含量也很低,显著低于其他未褐化的类型.
2.4 不同状态类型外植体的SOD活性变化
图1C结果表明,水曲柳体胚发生过程中,不同状态类型的外植体中SOD活性是不同的,且差异显著(P<0.05).未褐化发生愈伤组织(WY)外植体中SOD活性最高,其次为褐化发生愈伤组织(HY)和未褐化发生体胚(WT)的外植体,褐化长大(HD)、褐化发生体胚(HT)和褐化未长大(HW)外植体的第3,未褐化未长大(WW)和未褐化长大(WD)外植体的SOD活性最低.由此可见,水曲柳体胚发生过程中,愈伤组织的发生与外植体中的SOD活性有直接的关系,而体胚发生与外植体SOD活性关联不大.
2.5 不同状态类型外植体的POD活性变化
图1D中可以看出,水曲柳体胚发生过程中,不同状态类型的外植体中POD活性存在显著差异(P<0.05).各个褐化外植体中的POD活性均显著低于各个未褐化外植体的POD活性.褐化发生体胚(HT)的外植体中POD活性略高于其他褐化的外植体的POD活性,而未褐化发生体胚(WT)的外植体中POD活性却小于其他未褐化的外植体的POD活性.未褐化长大(WD)的外植体中的POD活性最高,未褐化发生愈伤组织(WY)和未褐化未长大(WW)的外植体中POD活性次高,即未褐化的外植体中,单纯体积长大的类型中的POD活性最高,而发生愈伤组织或体胚,POD活性就降低;但是没有发生任何变化(WW)的外植体的POD活性也较低.
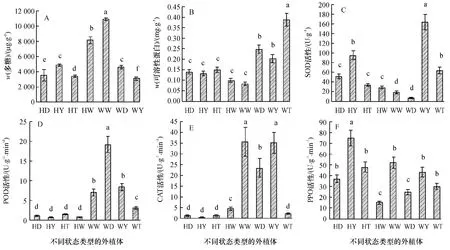
图1 水曲柳体胚发生过程中各状态类型外植体的不同生理指标变化Fig.1 Physiological index changes in different types of explants during somatic embryogenesis of Fraxinus mandshurica
2.6 不同状态类型外植体的CAT活性变化
图1E结果表明,水曲柳体胚发生过程中,不同状态类型外植体中的CAT活性有显著差异(P<0.05).除了未褐化发生体胚(WT)外植体的CAT活性极低外,其他未褐化的外植体中CAT活性均很高,其中未褐化未长大(WW)和未褐化发生愈伤组织(WY)外植体的CAT活性最高,未褐化长大(WD)的次之.褐化的外植体中CAT活性均很低,其中褐化长大(HD)、褐化发生愈伤组织(HY)和褐化发生体胚(HT)的外植体中CAT活性极低,显著低于褐化未长大(HW)的外植体.整体上来看,不管褐化与否,发生体胚的外植体的CAT活性均极低.
2.7 不同状态类型外植体的PPO活性变化
图1F结果表明,水曲柳体胚发生过程中,各不同状态类型外植体的PPO活性存在显著差异(P<0.05),但整体上来看,褐化和未褐化外植体的PPO活性均较高.其中,褐化发生愈伤组织(HY)的外植体的PPO活性明显高于其他各个类型的外植体.褐化未长大(HW)和未褐化长大(WD)的外植体中PPO活性最低,其他各种状态类型外植体的PPO活性居中.
3 讨论与结论
多糖类物质在体细胞胚胎发生过程中的作用主要是提供能量和参与形态建成.糖类化合物的积累与植物体细胞胚发生潜能的实现是密切相关的.水曲柳中褐化外植体的体胚发生能力较未褐化外植体高,也可能与其褐化外植体中糖含量有关.
可溶性蛋白含量与生物体适应环境的调控有关,杨和平等[15]在马唐 Digitaria sanguinalis胚状体发生的研究中发现,当胚胎发生受阻时,胚性愈伤组织中蛋白质合成较旺盛.但在愈伤组织生长过程中可溶性蛋白质含量在一段短暂的上升之后有一下降趋势,这与愈伤组织生长高峰期到来以前所需的蛋白及随后多转化为贮藏蛋白和结构蛋白有关.未褐化产生体胚的外植体中可溶性蛋白含量明显高于未产生体胚外植体,可能是由于胚性细胞的代谢较非胚性细胞旺盛,因此在从培养基中获得了有机与无机成分以后及时将其转化成能量和细胞物质,积累了蛋白质,以备细胞分化之用[16].但是在未褐化的4种外植体中,却是未褐化未长大的外植体蛋白含量最低,褐化外植体间的蛋白含量变化差异不大,推测可能是由于调控外植体褐化与否的细胞内部基因表达发生变化,从而影响了蛋白的合成.
植物体细胞胚发生和发育过程中抗氧化酶活性研究已有报道[17],研究枸杞Lycium barbarum体细胞胚发生过程中发现,SOD、POD和CAT相互配合来调节胚性细胞的分化和发育,这3种酶在胚胎发生与发育过程中具有特异性变化,表明体细胞胚胎发生与超氧化物对自由基的清除有关,认为体细胞胚发生及发育是大量酶特异性合成及参与代谢的结果.体细胞胚发生过程中活性氧的变化与体细胞胚的发生发育密切相关,因此其SOD活性也会发生相应的变化.据报道,SOD活性的增加与细胞分化频率成正比,对胚性细胞的分化以及胚胎发育有促进作用[18-19].有关植物体细胞胚胎发生过程中SOD活性的报道很有限,而且其中大多也是有关胚性和非胚性组织的.崔凯荣等[18]认为在胚性愈伤组织转入分化培养基后,随着胚性细胞的形成,SOD活性逐渐升高,当胚性细胞进一步分裂形成体细胞胚时,SOD活性达到最高峰,表明SOD活性升高对胚性细胞的分化及胚胎发育具有促进作用.这与本试验结果有相似之处.此外,0berle等[19]也认为,具分生组织的茎尖和根尖的SOD活性较高,即只有SOD活性升高,分化才会发生[20].H2O2是一种比较强的氧化剂,在植物体中存在和积累可导致破坏性的氧化作用,能将生物大分子的有关活性基团氧化,使生物大分子钝化,甚至完全失去生物活性.一般认为,体内的CAT活性与该器官的代谢强弱有密切关系,代谢强,产生的H2O2等副产物较多,CAT活性就高.过氧化氢酶是植物体内重要的活性氧消除系统成分之一,可以清除H2O2.据报道,CAT活性与植物抗逆性有关,与植物老化也有一定的关系,在老化的组织中其活性较强[21].很多学者都以CAT活性变化来研究植物的生长发育和进行植物的选育.
外植体褐化的机制是外植体在受损伤时,液泡中的酚类物质与细胞质中的PPO接触后,被氧化成相应的醌,再进一步反应形成黑色或褐色的色素沉淀[22].单一的多酚氧化酶活性和多酚含量并不能说明褐变的机理,多酚和多酚氧化酶在细胞内存在区域化分布,形成酚和酶的区域化分布的膜系统才是褐变发生的关键[14].组织培养过程中的褐变现象已经成为影响组培成功的重要因素.褐变现象主要是由多酚氧化酶(PPO)作用于天然底物酚类物质而引起的.组织培养中外植体酚类物质的含量与其褐变率成正相关,与成活率成负相关[20].现在许多报道认为POD是一种诱导酶,外界因素通过POD系统影响器官的生理变化和个体发育,其酶活力和同工酶的变化可能与一些高等植物的发育进程密切相关[21].在胚胎发生发育中相应的基因在时间和空间上顺序表达,有旧酶的消失和新酶的产生.POD同工酶也是较好的遗传信息表达的标志,它的合成受一系列基因所调控[22].
本文以水曲柳幼嫩合子胚子叶为外植体诱导水曲柳体胚发生,检测这些不同状态类型外植体的多糖含量、可溶性蛋白质含量及 SOD、POD、CAT、PPO活性等生理生化指标.结果发现,不同状态类型外植体的生理生化状态与体胚发生、愈伤组织发生、外植体褐化和生长发育存在非常有趣的相关关系,值得进一步通过分子生物学方法加以验证、解析.
[1]沈海龙.植物组织培养[M].北京:中国林业出版社,2005:107-110.
[2]孔冬梅,沈海龙,冯丹丹,等.水曲柳体细胞胚与合子胚发生的细胞学研究[J].林业科学,2006,42(12):130-133.
[3]李楠,孔冬梅,沈海龙.低温预处理对水曲柳体细胞胚胎发生的影响[J].植物生理学通讯,2009(6):579-582.
[4]孙桂君,孔冬梅,沈海龙.取材时期和母树来源对水曲柳体细胞胚诱导的影响[J].东北林业大学学报,2010,38(1):28-30.
[5]冯丹丹.外植体的母树来源对水曲柳体细胞胚胎发生的影响[D].哈尔滨:东北林业大学,2006.
[6]张宇.水曲柳体细胞胚胎发生的同步化控制[D].哈尔滨:东北林业大学,2007.
[7]张丽杰.取材时期对水曲柳合子胚外植体体细胞胚胎发生的影响[D].哈尔滨:东北林业大学,2006.
[8]刘春苹.水曲柳体胚发生伴随外植体褐化的生理机制及差异蛋白研究[D].哈尔滨:东北林业大学,2009.
[9]李合生.现代植物生理学[M].北京:高等教育出版社,2000.
[10]张龙翔,张庭芳,李令媛.生化实验方法和技术[M].北京:高等教育出版社,1997.
[11]杨和平,程井辰,周吉源,等.石刁柏体细胞胚胎发生过程中超氧物歧化酶活性的变化[J].植物学报,1993,3(6):190-193.
[12]张志良.植物生物学实验指导[M].北京:高等教育出版社,1990:154-155.
[13]陈建勋.植物生理学实验指导[M].广州:华南理工大学出版社,2002:120-121.
[14]罗晓芳,田砚亭,姚洪军.组织培养过程中PPO活性和总酚含量的研究[J].北京林业大学学报,1999,21(1):92-95.
[15]杨和平,程井辰.马唐体细胞胚胎发生过程中生理变化的研究[J].武汉植物学研究,1991,9(4):306-312.
[16]李付广,李秀兰,李凤莲.棉花体细胞胚胎发生及主要物质生化代谢机制[J].河南农业大学学报,1994,28(3):313-316.
[17]DUTTA GUPTA S,DATTA S.Antioxidant enzyme activities during in vitro morphogenesis of gladiolus and the effect of application of antioxidants on plant regeneration[J].Biologia Plantarum,2003,47(2):179-183.
[18]崔凯荣,任红旭,邢更妹,等.枸杞组织培养中抗氧化酶活性与体细胞胚发生相关性的研究[J].兰州大学学报,1998,34(3):93-99.
[19]OBERLEY L W,OBEBLEY T D.Role of antioxidant enzymes in cell immortalization and transformation[J].Molecular and Cellular Biochemistry,1988,84(2):147-153.
[20]St.CLAIR D K,OBERLEY T D,MUSE K E,et al.Expression of manganese superoxide dismutase promotes cellular differentiation[J].Free Radical Biology and Medicine,1994,16(2):275-282.
[21]胡家恕.超干红花种子抗老化作用及其机理[J].植物生理学报,1999,25(2):171-177.
[22]李斌,黎秋华,杨宏伟,等.植物多酚氧化酶分子生物学研究进展[J].广东茶业,2003,21(增刊1):8-11.
