草鱼钙网蛋白的克隆与组织表达
2012-09-25李旭威王海英吴锐全谢骏郁二蒙吴垠余德光王广军龚望宝
李旭威,王海英,吴锐全,谢骏,郁二蒙,吴垠,余德光,王广军,龚望宝
(1.中国水产科学研究院 珠江水产研究所 农业部热带亚热带水产资源利用与养殖重点实验室,广东 广州 510380;2.大连海洋大学 水产与生命学院,辽宁 大连116023)
钙网蛋白(Calreticulin,CRT)是一种高度保守的分子伴侣蛋白,是内质网(ER)中主要的钙结合蛋白之一。Ostwald等[1]于1974年首次从骨骼肌肌浆网中提纯得到钙网蛋白,之后Fliegel等[2]和Smith等[3]同时获得编码这种蛋白的cDNA,并将此蛋白命名为高亲和力Ca2+结合蛋白 (high affinity calcium binding protein,HACBP)。直到1991年,Opas等[4]发现 HACBP是一种存在于细胞内质网中的蛋白,并将其重新命名为钙网蛋白。
钙网蛋白在细胞受到应激如炎症、热休克、病毒感染时分泌到细胞外[5],从而起到保护细胞的作用。Asgari等[6]研究发现,菜粉蝶Pierisrapae血细胞表面的钙网蛋白具有吞噬作用;在中国明对虾Fenneropenaeuschinensis体内注射白斑综合征病毒(WSSV)6 h后,检测发现钙网蛋白的表达量显著增加,结果证实了钙网蛋白参与病毒侵袭后的免疫过程[7]。
钙网蛋白位于大多数有核细胞的内质网中,本身具有凝集素样功能[8-9],在结构上亦与补体C1q受体类似,补体C1q能与钙网蛋白结合,启动补体经典激活途径。同时,吞噬细胞膜上的LDL受体相关蛋白(LDL—receptor related protein,LRP)可识别凋亡细胞上钙网蛋白的功能受体。钙网蛋白与LRP相互作用可活化吞噬细胞内的相关信号传递系统,启动细胞的内吞过程[10]。
钙网蛋白基因目前已在斑马鱼Daniorerio、虹鳟Oncorhynchusmykiss[11]、牙鲆Paralichthysolivaceus、中国明对虾Fenneropenaeuschinensis[12]、斑节对虾Penaeusmonodon[13]、斑点叉尾鮰Ictaluruspunctatus[14]中克隆获得了钙网蛋白基因序列,在虹鳟、斑点叉尾鮰的钙网蛋白的研究中也初步了解了鱼类钙网蛋白的特性及其作用。Visudtiphole等[13]的研究表明,钙网蛋白可作为斑节对虾热应激的生物标记物。
作为中国淡水养殖的四大家鱼之一的草鱼Ctenopharyngodonidellus,在养殖过程中病害频发,由此造成的经济损失巨大[15-16]。本试验中,作者通过克隆和RACE技术获得草鱼钙网蛋白基因全长,并研究了钙网蛋白在草鱼不同组织中的表达规律,以期为进一步深入研究钙网蛋白的功能以及在鱼类免疫中的应用奠定基础。
1 材料与方法
1.1 材料
试验草鱼为3尾,购自黄沙水产品市场,体质量为(1.451±0.352)kg。解剖后取肌肉、肠道、肝胰脏、鳃、皮肤、鳍条、肾脏和脾脏,立即放入液氮中保存备用。
Trizol总RNA提取试剂盒购自Nitrogen公司;反转录试剂盒、pMD-19T载体、DL2000 DNA和Taq DNA聚合酶购自大连TaKaRa公司;SMARTTM RACE cDNA Amplification Kit和Advantage-2 PCR Kit均购自Clontech生物工程公司;琼脂糖凝胶DNA回收试剂盒和DH5α购自Tiangen公司。
1.2 方法
1.2.1 总RNA的提取与cDNA第一链的合成 RNA的提取参照Trizol说明书进行,利用紫外分光光度计和琼脂糖凝胶电泳检测RNA的纯度和质量。按照TaKaRa RNA PCR Kit试剂盒推荐的方法,以Oligo(dT)为引物合成cDNA第一链。具体步骤如下:取Oligo dT-接头引物 1 μL、dNTP mixture 1 μL、总RNA 4 μL,加ddH2O至10 μL,于65 ℃下水浴5 min,然后在冰上急速冷却;取该反应液10 μL、5×PrimeScript II Buffer 4 μL、RNase inhibitor 0.5 μL、PrimeScript I RTase 1 μL,用RNase Free ddH2O加至20 μL。在PCR仪中进行如下反应:42 ℃下变性30 min,99 ℃下退火5 min,5 ℃下延伸 5 min。
1.2.2 草鱼钙网蛋白基因cDNA核心序列克隆 根据GenBank中斑马鱼、虹鳟、小家鼠、人的钙网蛋白基因mRNA序列设计并合成1对简并引物——P1和P2(表1),产物预计长度为495 bp。以草鱼肌肉的cDNA为模板, 采用PCR扩增目的基因片段。PCR反应体系为25 μL,包括ddH2O 19.8 μL、Buffer(含MgCl2) 2.5 μL、dNTP Mix 1 μL、cDNA 1 μL。PCR反应条件为: 94 ℃下预变性3 min;94 ℃下变性1 min,49 ℃ 下退火30 s,72 ℃下延伸 1 min,共进行35个循环;最后在72 ℃下再延伸10 min。PCR扩增产物经15 g/L的琼脂糖凝胶电泳检测、回收后,与pMD19-T载体连接,并转化至大肠杆菌DH5α感受态细胞中。挑选阳性克隆,最后通过PCR反应检测得到阳性克隆,阳性克隆由上海英俊生物技术有限公司进行测序,所得cDNA片段与GenBank数据库中的已知序列进行Blast同源性分析。

表1 试验用引物序列
1.2.3 草鱼钙网蛋白基因cDNA 3′RACE的克隆 3′RACE的操作按照TaKaRa RNA PCR Kit(AMV)Ver 3.0试剂盒方法进行。首先以试剂盒提供的Oligo dT-Adaptor primer为引物进行逆转录反应,用上述的逆转录反应液、5×PCR Buffer 10 μL、ddH2O 28.75 μL、TaKaRa Ex Taq®HS 0.25 μL、上游特异性引物P3(表1)0.5 μL和M13 Primer M4 0.5 μL进行首次PCR反应。具体反应条件为:94 ℃下预变性3 min;94 ℃下变性30 s,55 ℃下退火30 s,72 ℃下延伸1 min,共进行30个循环。巢式PCR所用引物为M13 Primer M4和P4(表1)。PCR产物在15 g/L琼脂糖凝胶中电泳检测,并回收、克隆后测序。
1.2.4 草鱼钙网蛋白基因cDNA 5′RACE的克隆 根据克隆得到的草鱼钙网蛋白基因的cDNA核心片段,以SMARTTMRACE cDNA Amp lification Kit (Clotech)引物设计原则,设计2个下游引物P5与P6(表1),上游引物由试剂盒提供。5′RACE cDNA合成用试剂盒提供的5′-CDS Primer A与SMART II A Oligo引物。取总RNA 2 μL,合成5′RACE cDNA方法按照试剂盒说明进行。得到5′RACE cDNA后进行首次PCR扩增,反应条件为:94 ℃下变性30 s,65 ℃下退火30 s,72 ℃下延伸 3 min,共进行25个循环。取首次扩增产物5 μL,加入245 μL的Tricine-EDTA buffer,然后取5 μL作为巢式PCR的cDNA。扩增条件为:94 ℃下预变性30 s,65 ℃下退火30 s,72 ℃下延伸3 min,共进行20个循环。RACE产物纯化、克隆和测序方法同上。
1.3 钙网蛋白基因的氨基酸序列相似性分析
用NCBI中Blast软件将获得的cDNA序列进行拼接,得到钙网蛋白基因的全长cDNA序列。用Vector 8.0程序搜索开放阅读框并预测氨基酸序列,采用ClustalX 1.83 对不同物种的钙网蛋白基因的氨基酸序列进行比对,然后用Mega 3.1软件构建NJ进化树。
1.4 不同组织中Ci-CRT的表达
用“1.2”中的方法得到不同组织的cDNA模板,根据目的基因的全长cDNA序列,设计1对RT-PCR特异引物P7与P8(表1),以草鱼的β-actin基因序列设计内参引物P9与P10(表1)。PCR反应体系同上。PCR反应条件为:94 ℃下预变性3 min;94 ℃下变性30 s,49 ℃下退火30 s,72 ℃下延伸1 min,共进行32个循环;最后在72 ℃下再延伸10 min。试验重复3次。
2 结果
2.1 钙网蛋白基因全长cDNA序列的特征分析
草鱼钙网蛋白扩增产物经Vector NTI Suite 8.0软件包中的Contig Express程序进行重复序列的识别和拼接,得到钙网蛋白全长cDNA为1 389 bp,包含1 263 bp的开放阅读框,编码为421个氨基酸,长度为69 bp的5′UTR和长度为57 bp的3′UTR。利用ProtParam程序(http://www.expasy. org/)对氨基酸序列进行基本的物理化学性质分析,预测该蛋白的相对分子质量为48 700,理论等电点为4.33。经SignalP软件预测,氨基酸N-端是由21个氨基酸残基构成的疏水信号肽( MQIITLFPFVSAALLALAANA ),且发现两个保守的钙网蛋白家族标签(KHEQKIDCGGGYVKVF与IMFGPDICG),以及另外两个钙网蛋白家族保守重复序列A和B(PXXIXDPDAXKPEDWDE与GXWXPPXIXNPXYX)(图1)。
2.2 序列比对以及系统进化分析
选择哺乳类、鱼类、虾蟹类、昆虫等物种的钙网蛋白的氨基酸序列(图2)进行比对分析,结果发现,草鱼钙网蛋白基因功能域具有较高的保守性,在C-端都有保守K/HDEL标签,且氨基酸序列与斑马鱼(NM_131047.1)、虹鳟(NM_001124478.1)的相似性分别为86.26%、76.78%,与中国明对虾(DQ323054.1)的相似性为63.83%,与人(BT007448.1)、牛(BC140582.1)、果蝇(NP_524293)和小家鼠(NM_007591.3)的相似性分别为69.67%、70.38%、62.17%和69.19%。
对钙网蛋白基因的系统进化分析显示,构建的进化树与传统的生物界的划分比较一致。构建的分子进化树可以简单的将基因聚类为植物、脊椎动物和无脊椎动物。已获得的草鱼钙网蛋白核苷酸序列与小家鼠、人、牛、非洲爪蟾、其它的鱼类聚为一类;家蚕、果蝇与中国明对虾、太平洋牡蛎等无脊椎动物聚为一类;而小麦等植物位于进化树的最下方(图3)。
2.3 草鱼钙网蛋白基因的组织表达
通过半定量RT-PCR分析显示,钙网蛋白基因在草鱼的肌肉、肠道、肝胰脏、鳃、皮肤、鳍条、肾脏和脾脏中均有表达(图4)。以钙网蛋白基因与β-actin电泳带吸光度的比值绘制了钙网蛋白在草鱼各组织的表达相对丰度(图5)。从图5可见:钙网蛋白基因在肝胰脏、鳍条的表达量均较高,显著高于肌肉、肠道、鳃、肾脏、脾脏(P<0.05),但与皮肤无显著差异(P>0.05);钙网蛋白基因在皮肤中的表达量次之。
3 讨论
钙网蛋白是内质网上主要的Ca2+结合蛋白,具有调控细胞钙稳态及内质网钙储存功能的作用,并且参与细胞黏附、伤口愈合、调节免疫应答、参与病理变化等过程[17-18]。随着对钙网蛋白的深入研究,发现人类和小家鼠还具有不同基因型(2型钙网蛋白基因,calreticulin-2),在大鼠、牛中也相继发现2型钙网蛋白基因[19]。本研究中的草鱼钙网蛋白基因是根据NCBI数据库中虹鳟、牙鲆、斑马鱼等的钙网蛋白序列设计的简并引物克隆获得的,基因全长为1 389 bp,由基因推导的氨基酸序列在结构上与哺乳类的钙网蛋白一样,分为N-、P-和C- 3个功能域。经同源性比对发现,其与斑马鱼、虹鳟具有较高的同源性,序列含有两个高度保守的钙网蛋白家族标签(KHEQKIDCGGGYVKVF 100~116和IMFGPDICG 132~140)以及内质网回收信号四肽(H/KDEL)区域[20]。而所知的人类和小家鼠的calreticulin-2与calreticulin相比,其在C末端只具有-EL区域,而不具有钙网蛋白的回收信号四肽序列[19]。由此推知,本试验中从肝胰脏克隆的基因应为草鱼calreticulin。Liu等[14]发现,斑点叉尾鮰钙网蛋白具有calreticulin和calreticulin-like 2。草鱼钙网蛋白是否存在不同的基因型还有待进一步研究。

注:双下划线表示信号肽,方框表示两个保守的钙网蛋白家族标签,粗体下划线表示重复序列,星号表示终止密码子。Note:The predicted signal peptide is indicated with double underlined,CRT family signature motifs are boxed, repeat sequences are indicated with underlined bold text, and stop codon is indicated by asterisk.图1 草鱼钙网蛋白基因cDNA全长序列及推导的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of full-length Calreticulin(CRT) cDNA in grass carp
本试验中,利用RT-PCR方法检测钙网蛋白在草鱼肝胰脏、脾、肠道、鳃、肌肉、鳍条、皮肤、肾脏中的表达分布情况,结果表明,在草鱼的8个组织器官中均可检测到钙网蛋白的表达,其中在肝肝胰脏和鳍条中的表达量较高,且两者表达量都高于其它6个组织。Kales等[11]研究了虹鳟的钙网蛋白,在虹鳟的肌肉、头肾、肝胰脏、中肾、鳃、脾脏、心脏、脑和肠道中均检测到其表达,其中在肝胰脏中的表达量高于其它组织,这与本试验结果相一致。在Liu等[14]的研究中,选取斑点叉尾鮰的脾脏、肌肉、中肾、头肾、脑、心脏、鳃、胃、肠道和皮肤10个组织器官进行试验,结果表明,钙网蛋白在各组织中均能表达,亦发现其在肝胰脏中的表达量较高。肝胰脏作为免疫系统中的重要器官,由于其中存在大量的巨噬细胞等先天性免疫组分,在抑制肿瘤转移、修复损伤中发挥重要作用,许多抗病毒相关基因在肝胰脏中的表达量都较高[21]。
在体外试验中,钙网蛋白与HSP70、HSP90、gb96一样能够直接激活免疫细胞[22]。钙网蛋白在应激状态下也会发生变化,如钙网蛋白在热应激情况下,为了维持细胞内稳态,其表达呈上调的趋势,而在低氧、缺氧的情况下,钙网蛋白的表达也都呈上调趋势[23-24]。草鱼是中国淡水池塘养殖的主要种类之一,在养殖过程中,草鱼的发病率高且难治愈。鉴于钙网蛋白在免疫和应激方面存在的潜在作用,加强对草鱼钙网蛋白功能的进一步研究是十分必要的。

注:人Homo sapiens(BT007448.1),小家鼠Mus musculus(NM_007591.3),牛Bos taurus(BC140582.1),果蝇Drosophila melano gaster(NM_079569.4),中国明对虾Fenneropenaeus chinensis(DQ323054.1),草鱼Ctenopharyngodon idellus, 斑马鱼Danio rerio(NM_131047.1),虹鳟Oncorhynchus mykiss(NM_001124478.1)。图2 草鱼钙网蛋白基因氨基酸序列的比对Fig.2 Amino acid sequence alignment of Calreticulin (CRT) in grass carp
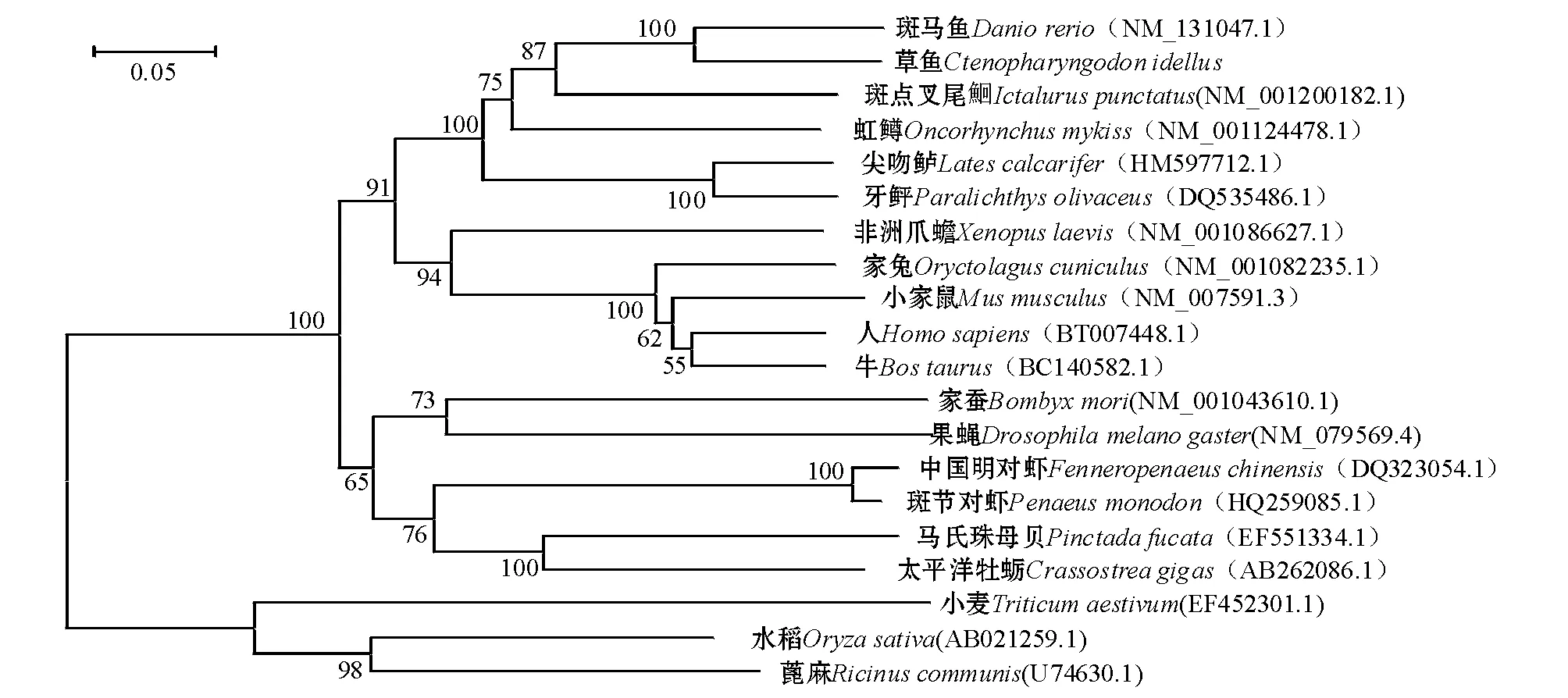
图3 钙网蛋白的系统进化分析Fig.3 Phylogenetic analysis of Calreticulin (CRT)

注:M为DL 2000 marker;1~8、9~16、17~24为3组重复,依次对应肌肉、肠道、肝胰脏、鳃、皮肤、鳍条、肾脏、脾脏。Note:M,DL 2000 marker;1-8,9-16,17-24 triplication;1,muscle;2,intestine; 3,hepatopancreas; 4,gill; 5,skin; 6,ray; 7,kidney; 8, spleen.图4 Calreticulin mRNA 在草鱼不同组织中的表达电泳图谱Fig.4 Expression of Calreticulin mRNA in different tissues of grass carp by RT-PCR

图5 Calreticulin mRNA在草鱼不同组织中表达量的分析Fig.5 The significance tests of Calreticulin mRNA expression in different tissues of grass carp
参考文献:
[1] Ostwald T J,Maclennan D H.Isolation of a high affinity calcium-binding protein from sarcoplasmic reticulum[J].The Journal of Biologlcal Chemist,1974,249:974-979.
[2] Fliegel L,Burns K,MacLennan D H,et al.Molecular cloning of the high affinity calcium-binding protein (calreticulin) of skeletal muscle sarcoplasmic reticulum[J].The Journal of Biologlcal Chemist,1989,264:21522-21528.
[3] Smith G L,Koch E.Multiple zones in the sequence of calreticulin (CRP55,calregulin,HACBP),a major calcium-binding ER/SR protein[J].EMBO Journal,1989,8(12):3581- 3586.
[4] Opas M,Dziak E,Fliegel L,et al.Regulation of expression and intracellular distribution of calretieulin,a major calcium binding protein of nonmusele cells[J].Journal of Cellular Physiology,1991,149 (1):160-171.
[5] Park B J,Lee D J,Yu J R,et al.Calreticulin,a calcium-binding molecular chaperone,is required for stress response and fertility inCaenorhabditiselegans[J].Molecular Biology of the Cell,2001(9):2835-2845.
[6] Asgari S,Schmidt O.Is cell surface calreticulin involved in phagocytosis by insect hemocytes?[J].Journal of Insect Physiology,2003,49(6):545-550.
[7] Wang B,Li F H,Dong B,et al.Discovery of the genes in response to white spot syndrome virus (WSSV) infection inFenneropenaeuschinensisthrough cDNA microarray[J].Marine Biotechnology,2006,8(5):491-500.
[8] Kasper G,Brown A,Eberl M,et al.A calreticulin-like molecule from the human hookwormNecatoramericanusinteracts with Clq and the cytoplasmic signaling domains of some integrins[J].Parasite Immunology,2001,23(3):141-152.
[9] Michalak M,Corbett E F,Mesaeli N,et al.Calreticulin:one protein,one gene,many functions[J].Biochem J,1999,344(2):281-292.
[10] Gardai S J,McPhillips K A,Frasch S C,et al.Cell-surface calreticulin initiates clearance of viable or apoptotic cells through trans-activation of LRP on the phagocyte[J].Cell,2005,123(2):321-334.
[11] Kales S,Fujiki K,Dixon B.Molecular cloning and characterization of calreticulin from rainbow trout(Oncorhynchusmykiss)[J].Immunogenetics,2004,55 (10) :717-723.
[12] Wei L N,Li F H,Xiang J H,et al.Molecular characteristics and expression analysis of calreticulin in Chinese shrimpFenneropenaeuschinensis[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2007,147(3): 482-491.
[13] Visudtiphole V,Watthanasurorot A,Klinbunga S,et al.Molecular characterization of Calreticulin:A biomarker for temperature stress responses of the giant tiger shrimpPenaeusmonodon[J].Aquaculture,2010,308(1):100-108.
[14] Liu H,Peatman E,Wang W Q,et al.Molecular responses of calreticulin genes to iron overload and bacterial challenge in channel catfish (Ictaluruspunctatus)[J].Developmental and Comparative Immunology,2011,35(3):267-272.
[15] 张春雨.池塘草鱼病害的发生与防治[J].现代农业科技,2008,22:240-242.
[16] 王方华,李安兴.草鱼病毒性出血病研究进展[J].南方水产,2006,2(3):66-71.
[17] Kwon M S,Park C S,Choi K,et al.Calreticulin couples calcium release and calcium influx in integrin-mediated calcium signaling[J].Molecular Biology of the Cell, 2000,11:1433-1443.
[18] Johnson S,Michalak M,Opas M,et al.The in and outs of calreticulin:from the ER lumen to the extracellular space[J].Trends Cell Biol,2001,11 (3):122-129.
[19] Persson S,Rosenquist M,Sommarin M.Identification of a novel calreticulin isoform (Crt2) in human and mouse[J].Gene,2002,297(1-2):151-158.
[20] Michalak M,Parker J M R,Opas M.Ca2+signaling and calcium binding chaperones of the endoplasmic reticulum[J].Cell Calcium,2002,32(5-6):269-278.
[21] Pigeon C,Ilyin G,Courselaud B,et al.A new mouse liver-specific gene,encoding a protein homologous to human antimicrobial peptide hepcidin,is over expressed during iron over load[J].Biol Chem,2001,276(11):7811-7819.
[22] Hong Ch,Qiu X, Li Y, et al.Functional analysis of recombinant calreticulin fragment 39-272:implications for immunobiological activities of calreticulin in health and disease[J].The Journal of Immunology, 2010,185(8):4561-4569.
[23] Nguyen T Q,Capra J D,Sontheimer R D. Calreticulin is transcriptionally upregulated by heat shock,calcium and heavy metals[J].Molecular Immunology,1996,33(4-5):379-386.
[24] Kawabe S,Yokoyama Y.Molecular cloning of calnexin and calreticulin in the Pacific oysterCrassostreagigasand its expression in response to air exposure[J].Marine Genomics,2010,3(1) 19-27.
