南方四季杨高效遗传转化体系的建立
2012-09-08王红莉苏小曼李甫金罗建勋辜云杰
王红莉,苏小曼,李甫金,罗建勋,辜云杰,宋 鹏
(1.成都市林木种苗站,四川 成都 610031;2.内江市林业局,四川 内江 641000;3.四川省林业科学研究院,四川 成都 610081;4.平武县林业局,四川 平武 522550)
南方四季杨为杨柳科(Salicaceae)杨属(Populus L.)高大乔木,常绿/半常绿。其母本是美国密西西比州南方种源的美洲黑杨(P.deltoides Marsh.60/160),父本是智利黑杨(P.nigra cv.Chile)。是国内首选出的人工杂交雄性不育优良无性系。从2002年先后引种到云南、重庆、江苏南京、湖北、湖南、海南、上海、广东深圳及陕西的汉中地区,主要作为城乡园林绿化树种、短周期纤维工业用材林和大径级的胶合板材进行推广,2006通过四川省林木良种审定(认定)委员会审定(良种编号:川S-ETS-PDN-001-2006)。
该品种生长快、树干通直圆满、耐水湿、木材基本密度大、抗风折、落叶晚(比其他黑杨派杨树晚落叶30 d~40 d),在昆明、西昌、海南基本不落叶,翌年春天萌动早,不飞花粉,不会对引种发展地构成生态威胁。但是,南方四季杨在大量推广过程中遇到的较严重虫害问题,近年来造成了巨大的经济和生态损失,目前所采用的防治措施虽有一定的成效,但还是存在较大的局限性。加上雄性不育,不可能采用常规杂交育种进行改良,只寄希望通过转基因技术来改良其抗性。
本研究旨在建立南方四季杨组培体系的基础上优化南方四季杨遗传转化体系,拟将抗虫基因Cry3Aa导入南方四季杨的基因组中,减轻害虫对南方四季杨的危害,同时,为下一步通过基因工程手段改良该品种奠定技术基础。
1 材料与方法
1.1 实验材料
1.1.1 植物材料、菌株及化学试剂
植物材料为南方四季杨无菌苗。
菌株为以超表达载体pBI121为载体的短肽融合的Cry3Aa(Bt基因)(中国林科院林业所卢孟柱课题组馈赠)带有Cry3Aa目的基因的pBI121载体见图1。

图1 带有Cry3Aa目的基因的pBI121载体
抗生素选择卡那霉素(Kan)(用无菌水溶解,过滤灭菌),利福平(Rif)(用二甲基甲酰胺溶解)、特美汀(TMT)(用无菌水溶解,过滤灭菌),-20℃保存。
1.1.2 培养基
(1)细菌固体培养基(LB培养基):
(2)细菌液体培养基为不添加琼脂的LB培养基;
(3)植物培养基(各培养基中均添加琼脂5 g·L-1,蔗糖30 g·L-1,调节 pH 至 5.8 ~6.0)
预培养、共培养培养基MS+6-BA0.25 mg·L-1+NAA0.09 mg·L-1;
筛选分化养基:MS+6-BA0.25 mg·L-1+NAA0.09 mg·L-1+Kan30 mg·L-1+TMT50 mg·L-1;
生根培养基:1/2MS+IBA0.05 mg·L-1+IAA0.1 mg·L-1+Kan30 mg·L-1+TMT50 mg·L-1。
1.2 实验方法
1.2.1 菌株的活化
(1)菌株的活化
取保存于-80℃的菌株,冰上融化,用枪头蘸取少量菌液在含有相应抗生素的LB培养基平板上划线,27℃倒置培养至长出单菌落。
(2)菌液的制备
从平板上挑取一个生长饱满的菌落,接种到含有相应抗生素(Kan 30 mg·L-1,Rif(利福平)15 mg·L-1)的100 ml液体 LB 培养基中,27℃、230 rpm振荡培养12 h,即可用于浸染。
1.2.2 外植体的选取
选取生长健壮、颜色亮绿的组培苗,从顶端开始的第3片~4片叶子,无菌条件下用手术刀片将叶片划伤数刀。
1.2.3 实验正负对照的设置
对照是指在既定条件下应该出现实验预期结果的实验参照,对照包括正对照和负对照。针对本实验,正对照是指外植体不经过农杆菌浸染,也无卡那霉素等选择压力,直接放置于分化培养基上,让其正常生长;负对照是指外植体不经农杆菌浸染,但是置于含有卡那霉素等抗生素的培养基(即和转化外植体一样的培养基)上生长。
1.2.4 遗传转化程序
遗传转化程序包括以下5个主要步骤:预培养、浸染、共培养、筛选培养、抗性芽生根。
1.2.5 实验设计
根癌农杆菌介导的遗传转化效率受多个因子的影响,但要设计一个试验,把所有的因子及其交互作用对转化效率的影响都包括在内是一项巨大的工作。因此试验采取不考虑交互的实验设计,每次固定其它因子,只研究单因子变量对转化效率的影响。
根据转基因研究的一般情况,单因子变量为:预培养3 d,浸染时间5 min,浸染后26℃暗处共培养3 d。本实验共研究5个因子对转化效率的影响。最终将5个因子的最佳条件组合,得出基本转化条件。具体实验程序如下:
(1)选择压的筛选
卡那霉素浓度设置 0 mg·L-1,10 mg·L-1,20 mg·L-1,30 mg·L-1,40 mg·L-1,50 mg·L-1等 6个梯度。在南方四季杨不定芽分化培养基中加入上述浓度的卡那霉素,将无菌苗苗叶片切为0.5 cm大小,置于培养基中,每个处理接种3个培养皿,接种量以铺满培养皿空间为宜,均重复3次。半个月后观察,确定南方四季杨叶片对卡那霉素的抗性浓度。
(2)影响遗传转化效率的主要因素培养条件的筛选
①预培养时间:设置 0 d,1 d,2 d,3 d,4 d 等 4个梯度。
②浸染时间:设置 5 min,10 min,15 min,20 min,25 min等5个梯度。
③共培养时间:设置 0 d,1 d,2 d,3 d,4 d 等 5个梯度。
④共培养温度:设置22℃,25℃,28℃3个梯度。
⑤乙酰丁香酮的影响:设置0 umol·L-1,100 umol·L-1,200 umol·L-1,300 umol·L-1,400 umol·L-15 个梯度。
以上实验每个处理接种5个培养皿,根据培养皿的大小,每个培养皿接种6~12不等叶片数,均重复3次,分化率(%)=产生抗性芽的叶片数/接种的叶片总数×100%。
1.2.6 数据统计分析
采用Excel2007和SPASS软件分析。
2 结果与分析
2.1 选择压的选择(抗生素Kan浓度的筛选)
转基因过程中一个重要步骤就是阳性植株的初步筛选,本实验目的基因构建在载体pBI121上,根据该载体质粒的特点,其上有对Kan有抗性的NPTⅡ(新霉素磷酸转移酶基因),因此,表达该基因的细胞对抗生素Kan会表现出抗性,在含有该抗生素的培养基上,能够存活的再生芽初步被确定为转化体。但是,不同植株对抗生素的敏感度不同,浓度过高或者过低都会影响到转化效率,抗生素Kan的添加,既要能有效抑制非转化细胞的生长,使之慢慢变黄、死亡,又不能影响转化细胞的正常生长[1]。由于杨树叶片对Kan十分敏感,而美洲黑杨及其杂种的不定芽再生非常困难[2,3],因此,本实验梯度设置较小,选择压选择的实验结果见表1。

表1 不同卡那霉素(Kan)浓度对南方四季杨叶片分化的影响
由表1可以看出:实验过程中,随着Kan浓度的增加,叶片分化率下降,褐化明显急剧增加,20 mg·L-1时只有小部分分化,部分发黄,浓度≥30 mg·L-1时没有分化芽产生。从上表可以看出,南方四季杨对Kan比较敏感,30 mg·L-1可以作为其临界值。实验情况见图2。
2.2 影响转化效率的因素
2.2.1 预培养时间
关于浸染前预培养的必要性,因植物种类不同而异,大多数研究认为:预培养是有必要的,主要原因包括两点:(1)减少外源污染。如果外植体被污染,在预培养中有可能被淘汰[1];(2)有助于整合外源基因。预培养可使伤口周围的细胞得以修复,伤口愈合过程中分泌的酚类物质诱导农杆菌的Vir基因启动;预培养可促进细胞分裂,而处在分裂状态的细胞更易整合外源DNA;在高渗透压培养基上进行外植体预培养还可以减少伤口处的细胞伤流液,使细胞内的液泡减少并使细胞发生一定程度的质壁分离,使农杆菌易于接触伤口周围的细胞,从而提高转化率[4]。不同预培养时间对南方四季杨抗性芽分化的实验结果见表2,影响结果见表3。

图2 Kan 浓度筛选(依次为 0,10,20 mg·L-1浓度)
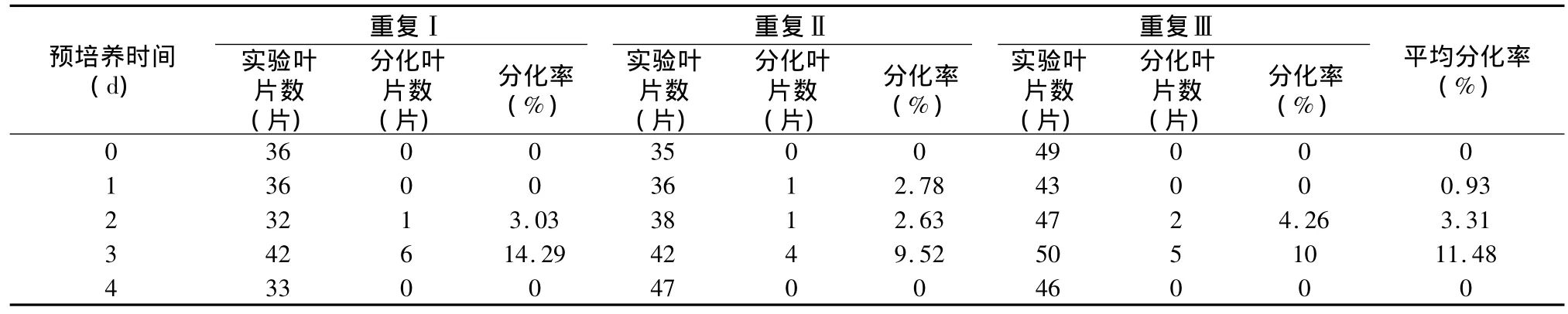
表2 不同预培养时间条件下南方四季杨抗性芽分化的实验结果

表3 不同预培养时间对南方四季杨抗性芽分化的影响
从表3可以看出,对于南方四季杨而言,没有经过预培养的外植体产生抗性芽的概率为0,预培养时间与抗性芽分化出现的频率呈极显著性相关,并且当预培养3 d时,抗性芽产生频率最高,达到11.48%。
2.2.2 浸染时间
浸染时菌液OD值和浸染时间都会影响转化效率,农杆菌浸染时间过短,附着于外植体切口处的农杆菌不足,抗性芽的产生频率明显降低,有时甚至无抗性芽产生,将大大降低转化效率;但如果时间过长,褐化情况也会随之严重[5,6]。
本实验将OD600统一为约0.4,研究浸染时间对转化率的影响。不同浸染时间对南方四季杨抗性芽分化的实验结果见表4,影响结果见表5。

表4 不同浸染时间条件下南方四季杨抗性芽分化的实验结果

表5 不同浸染时间对南方四季杨抗性芽分化的影响
如表5所示,浸染时间与抗性芽分化出现的频率呈极显著性相关,当侵染时间为10 min~15 min时,抗性芽产生频率较高,当侵染时间为20 min时,抗性芽产生频率急剧下降,故应将侵染时间该控制在15 min。
2.2.3 共培养时间
经过农杆菌浸染后,外源基因不会立即整合进植物细胞,如果把外源基因插入T-DNA中,就有可能携带进入受体植物并整合到染色体上。因此需要通过共培养为该过程提供充足的时间。但是,共培养所使用的培养基不添加任何抗生素,时间过短,后期引起污染的可能性较小,但是转化率也会随之降低;时间过长,可以提高整合外源基因效率,但是后期的农杆菌污染可能无法控制。不同共培养时间对南方四季杨抗性芽分化的实验结果见表6,影响结果见表7。
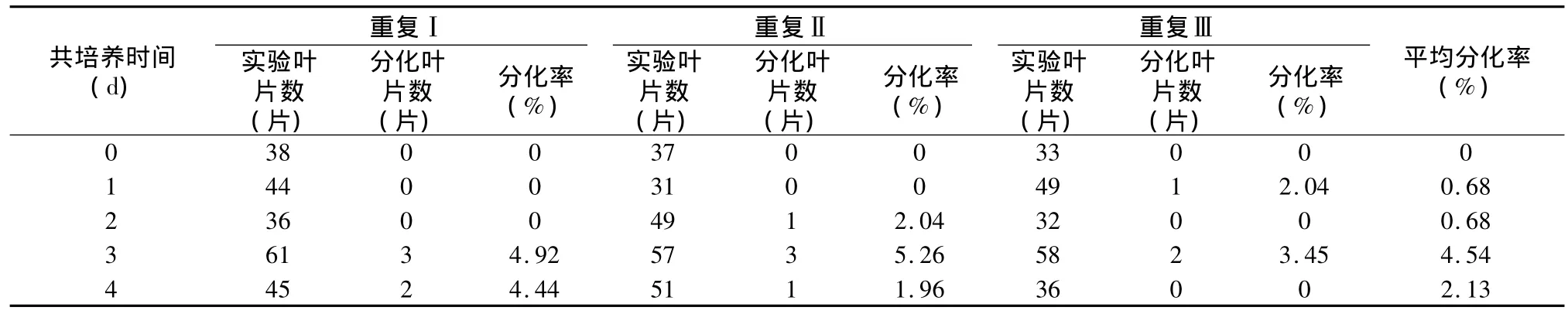
表6 不同共培养时间条件下南方四季杨抗性芽分化的实验结果
从表7可以看出:共培养时间与抗性芽的分化呈显著性关系,0 d~2 d的时间产生的分化芽很少,3d的培养时间,分化率达到最高,周围有少量农杆菌菌落,共培养4 d时有较大幅度的下降,并且叶片边缘有明显的农杆菌产生。因此,初步认定南方四季杨遗传转化共培养时间为3 d。
2.2.4 共培养温度
农杆菌正常生长的适宜温度为28℃,本实验设置3个共培养温度,实验结果显示:只有22℃和25℃有抗性芽产生,并且数量很少,说明共培养温度在一定范围内(22℃ ~25℃),对转化效果影响不大。不同共培养温度对南方四季杨抗性芽分化的实验结果见表8,影响结果见表9。
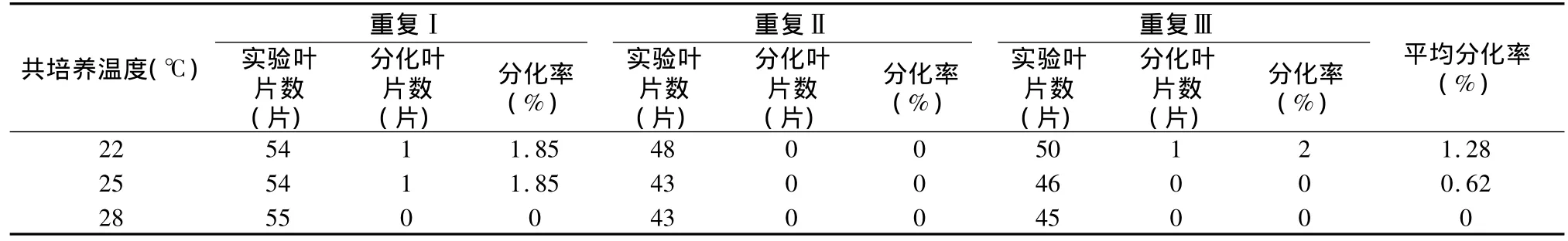
表8 不同共培养温度条件下南方四季杨抗性芽分化的实验结果

表9 不同共培养温度对南方四季杨抗性芽分化的影响
从表9可以看出:本实验得出的结果在一定范围内(22℃ ~25℃),但同时也确定了南方四季杨转化过程中,共培养温度不宜过高。
2.2.5 乙酰丁香酮对转化效率的影响
乙酰丁香酮(AS)的作用原理是其可诱导农杆菌Vir基因活化,从而促进外源基因的整合。本实验将乙酰丁香酮添加在浸染的菌液中,每隔5 min摇匀一次,使其与菌液中混合均匀,半个小时后浸染。菌液中添加不同浓度的AS对南方四季杨抗性芽分化的实验结果见表10,影响结果见表11。
从表11可以看出:实验结果AS的添加与抗性芽的分化关系不显著,虽从数据来看,添加300umolL-1时,一定程度上可以提高转化效率,但影响不大。
3 结论与讨论
3.1 结论
确定了30 mg·L-1的Kan浓度作为南方四季杨遗传转化过程中的选择压;

表10 菌液中添加不同浓度的AS条件下南方四季杨抗性芽分化的实验结果
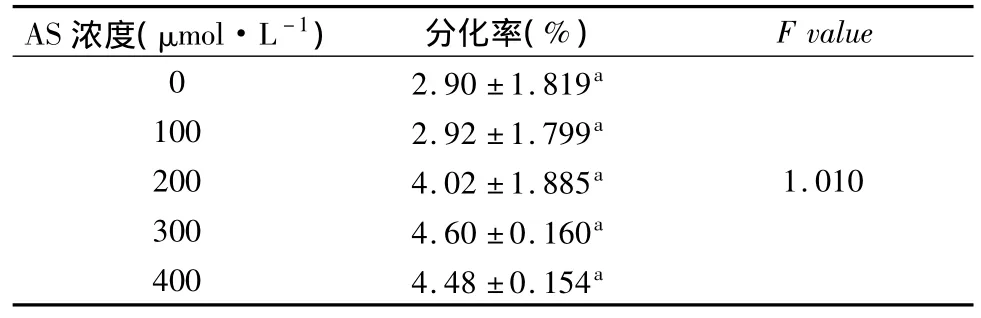
表11 菌液中添加不同浓度的AS对南方四季杨抗性芽分化的影响
南方四季杨遗传转化过程中优化后条件是:叶盘法,预培养3 d;OD600=0.4的菌液浸染10 min~15 min;浸染前菌液中加入 AS300 μmol·L-1;22℃~25℃条件下暗处共培养3 d。
3.2 讨论
一直以来,杨树作为木本植物中的模式植物进行遗传转化研究。但是同一属的不同种影响表现还是有很大区别的。本实验研究的是南方四季杨,选取了遗传转化过程中的5个主要因素进行研究。
Confalonieri等转化欧洲黑杨(P.nigra L.)实验结果证明:无预培养或预培养1 d对其转化率没有显著影响[7];Confalonieri等研究欧美杨(P.euramericana Neva.)和美洲黑杨(P.deltoids Marsh.)时预培养24 h[8]。本实验结果得到的预培养天数为3 d,与前人研究同一派杨树的结果差异较大,并且不同浓度间差异极显著,有待于进一步观察了解预培养阶段植物体内部发生的微小变化,分析总结出具体原因所在。
浸染时菌液OD值和浸染时间亦会影响转化效率,Leple等用OD600=0.3的菌液浸染杨树杂种(P.tremula L. × P.alba L.)16 min[9];郝贵霞认为用OD600=0.2~0.4的菌液浸染究毛白杨(P.tomentosa Carr.)时间应保持在10 min ~20 min[10]。本实验研究结果为15 min,与前人相关方面的研究结果基本一致。
在欧洲黑杨遗传转化研究中,Confalonieri等认为共培养48h最佳[7]。本实验研究结果显示:共培养2 d时,分化芽很少,但3 d,分化芽迅速增多。
有研究报道:用农杆菌介导转化菜豆和烟草的实验中,当温度超过22℃时,报告基因的表达大大下降,27℃时特别低,29℃时就完全检测不出了[11]。但也有报道显示水稻转愈伤组织转化中共培养温度在37℃时转化率较高[12]。南林895的转化实验中,得出的结果是转化频率最高的28℃,说明较高的温度更有利于农杆菌的转化[13]。因此,不同植物共培养过程中对温度的需求差异是较大的,本实验得出的结果与 Dillenw[11]的研究基本一致。但是,本实验设置的温度范围较小,更低的温度是否更有利于提高转化效率,有待于进一步研究。
新疆杨的研究中,添加乙酰丁香酮的培养基转化率都明显高于对照,最适浓度为 80 μmol·L-1,浓度过高,则转化率反而下降[14]。杨属内常用的乙酰丁香酮添加浓度为 200 μmol·L-1[5,11],浓度较低,其原因有三:首先乙酰丁香酮是一种植物受伤后分泌的酚类物质;其次杨树是农杆菌的天然宿主[9],细胞受伤后本身就有酚类物质的分泌;最后,乙酰丁香酮对农杆菌的生长可能不利。本实验结果与理论结果存在一定偏差,可能是由于植物本身的特殊性,也有可能是乙酰丁香酮添加的时间所造成的,前人大多数是添加进培养基中,而本实验添加于浸染的菌液中,至于哪种添加效果明显还有待于进一步对比研究。
实验结果表明,对于黑杨派杨树而言,本研究结果与以往的研究结果差异表现较大的是预培养天数和乙酰丁香酮的添加。其中,乙酰丁香酮的添加与大多数研究结论相反,关于乙酰丁香酮的作用原理有待于进一步研究。
另外,影响遗传转化效率的因素还有很多:乙酰丁香酮的添加时机、菌液制备方法、培养基pH值以及叶片放置方法等。要想取得最高转化率,实验过程中各个因素都要进行优化,并且这些因素各自不是单独存在的,需要考虑个因素之间的交互作用,这样一来,要设计一个实验,工作量是相当大的。因此,本实验选取几个重要的因素进行优化,得出了较优遗传转化体系。
叶盘法转化过程中,抗性芽的筛选是一个重要环节,能够在含有一定剂量的抗生素培养基中良好生长的转化体被初步确定为阳性转化体。本实验所有生根的抗性芽中,阳性植株所占的比例很低,很大原因是在抗性芽生根的过程中,采用的选择压太小。在最优条件下,南方四季杨遗传转化结果见图3、图4和图5。

图3 叶片分化前

图4 抗性芽分化

图5 抗性芽生根
[1]王关林,方宏筠主编.植物基因工程(第二版)[M].北京:科学出版社,2002.
[2]Sellmer J C,McCown B H,Haissig B E.Shoot culture dynamics of six Populus clones.Tree Physiol,1989,5:219 ~ 227.
[3]Coleman GD,Ernst SG.Shoot induction competence and callus determination in Populus deltoids.Plant Sci,1990,71:83 ~92.
[4]林荣呈,陈龙清.提高根癌农杆菌介导的香石竹遗传转化效率的研究.[J].林业科学研究,2003,16(21):23 ~128.
[5]Confalonieri M,Balestrazzi A,Biosffi S,Cella R.Factors affecting Agrobacterium tumefaciens-mediated transformation in several black poplar clones.Plant Cell Tissue Organ Cult,1995,43,215 ~222.
[6]Noel N,Leple J C,Pilate G.Optimization of in vitromicropropagation and regeneration for Populus x interameramericana and Populus x euramericana hybrids(P.deltoids,P.trichocarpa,and P.nigra)[J].Plant Cell Report,2002,20:1150 ~ 1155.
[7]Confalonieri M,Allegro G.Genetic transformation of Populus nigra by Agrobacter tumeraciens[J].Plant Cell Reports,1994,13(7):256~261.
[8]Confalonieri M,Balestrazzi A,Cella R.Genetic transformation of Populus deltoids and P.euramericana clones using Agrobacterium tumeraciens[J].Plant Cell Tissue and Organ Culture,1997,48(1):53~61.
[9]Leple J C,Brasileiro A C M,Michel M F,et al.Transgenic poplars:expression of chimeric genes using four different constructs[J].Plant Cell Report,1992(11):137 ~141.
[10]郝贵霞,朱桢,朱之悌.毛白杨遗传转化体统优化的研究[J].植物学报,1999,4(9):936 ~940.
[11]Dillen W,Cleroq J D.Kapila J,et al.The effect of temperature on Agrobacterium tumefaciens-mediated gene transfer to plants[J].The Plant Journal,1997,12(6):1459 ~1463.
[12]刘学志,张旭.用LBA4404/pCAMBIA系列转化水稻的最佳条件[J].复旦学报,1999,38(4):439 ~443.
[13]房丹.南林895杨抗虫转基因研究,南京林业大学硕士论文,2005.
[14]诸葛强,王婕琛,等.新疆杨高效遗传转化体系的建立[J].植物资源与环境学报,2003,12(4):6 ~10.
