夏秋用家蚕品种7 个数量性状的配合力与遗传力分析
2012-07-15艾均文司马杨虎朱勇何行健薛宏黄平叶学林钟天生
艾均文,司马杨虎,朱勇,何行健,薛宏,黄平,叶学林,钟天生
(1.湖南省蚕桑研究所,湖南 长沙 410127;2.苏州大学 基础医学与生物科学学部,江苏 苏州 215123;3.西南大学 生物技术学院,重庆 400715;4.云南省农业科学院 蚕桑蜜蜂研究所,云南 蒙自 661101;5.广东省茂名市蚕业技术推广中心,广东 化州 525100)
家蚕一代杂交种的推广使用是农业生产上利用杂交优势的典范[1]。随着优良杂交种的大面积推广,“遗传侵蚀”导致家蚕品种同质化的问题日显突出,因此,对不同地理来源品种资源进行引进与评价,充分挖掘多样性家蚕地方品种资源的潜力,对目前进行新品种选育与种质创新具有十分重要的现实意义[2]。在家蚕品种资源的评价指标中,除了其自身的丰产性与强健性指标外,配合力也是衡量其育种价值的重要指标。笔者在一批地方代表性品种资源中选取来自不同生态区域的8 个中系与8 个日系夏秋蚕品种,针对丰产性和强健性方面的主要数量性状进行配合力与遗传力分析,旨在对这些新蚕品种的育种潜力进行初步评价,并为纯系选育与一代杂交种组配提供参考依据。
1 材料与方法
1.1 材料
选取来自不同家蚕生态区域的夏秋用家蚕品种16 个,其中有8 个中系亲本(芙蓉、试抗、云1、夏芳A、试6、932、C7、C5)和8 个日系亲本(湘晖、日3、3087、7532、日丰、0802B、秋白B、湘T)。
1.2 方 法
2011年春,根据家蚕品种选育过程中普遍采用的中、日两系品种互交型式[3],利用不完全双列杂交试验设计(NCⅡ),以8 个日系品种为母本(P1组),以8 个中系品种为父本(P2组),配制成64 个杂交组合。2011年夏季在相同饲养条件下进行比较饲养,将4 龄起蚕饲养24 h 后分成3 区,每区200 头,按随机区组设计,共计192 个小区。
1.3 数据处理
按照《2011年国家桑蚕品种实验室鉴定实施方案》,评定、统计各试验小区的丰产性状(全茧量、茧层量、茧层率、万蚕产茧量、万蚕茧层量)与强健性状(死笼率、虫蛹统一生命率)。将茧层率、死笼率与虫蛹统一生命率数值进行反正弦转换后再进行方差分析。
用DPS 软件进行统计分析。
2 结果与分析
2.1 各数量性状的配合力方差
对64 个组合7 个性状的调查数据进行方差分析(表1)。各性状重复区组间的差异不显著,而组合间的差异均达到极显著水平。将组合间遗传方差分为中系、日系亲本的一般配合力方差及日系×中系组合的特殊配合力方差,对其进行分析可知:各性状的一般配合力方差均达极显著水平,日系×中系组合所有性状的特殊配合力方差也达显著或极显著水平,表明这些性状的遗传除了受加性遗传控制外,还在一定程度上受到亲本间互作效应的影响。

表1 家蚕7 个数量性状的配合力方差Table 1 Variance analyses of combining abilities for seven quantitative characters of silkworm
2.2 各品种资源的一般配合力效应
从表2 可以看出,同一亲本各性状间的一般配合力(GCA)相对效应值均存在明显差异,表明亲本在不同性状上的GCA 效应值是不同的,其中,死笼率效应值的负向、其他性状的正向为家蚕育种工作的追求方向。中系品种的C5、日系品种的日3在各自不同地理品系中5 个丰产性状的GCA 效应值均为最高;中系品种试抗、日系品种湘晖强健性状的GCA 效应值最高。中系亲本932 的5 个丰产性状的效应值均为负,且死笼率的效应值与育种目标方向相反;日系亲本3087 的2 个强健性状与4 个丰产性状的效应值均与育种方向相反。从综合表现来看,中系夏芳A、C5、云1和日系日3的丰产性状及强健性状表现优良。
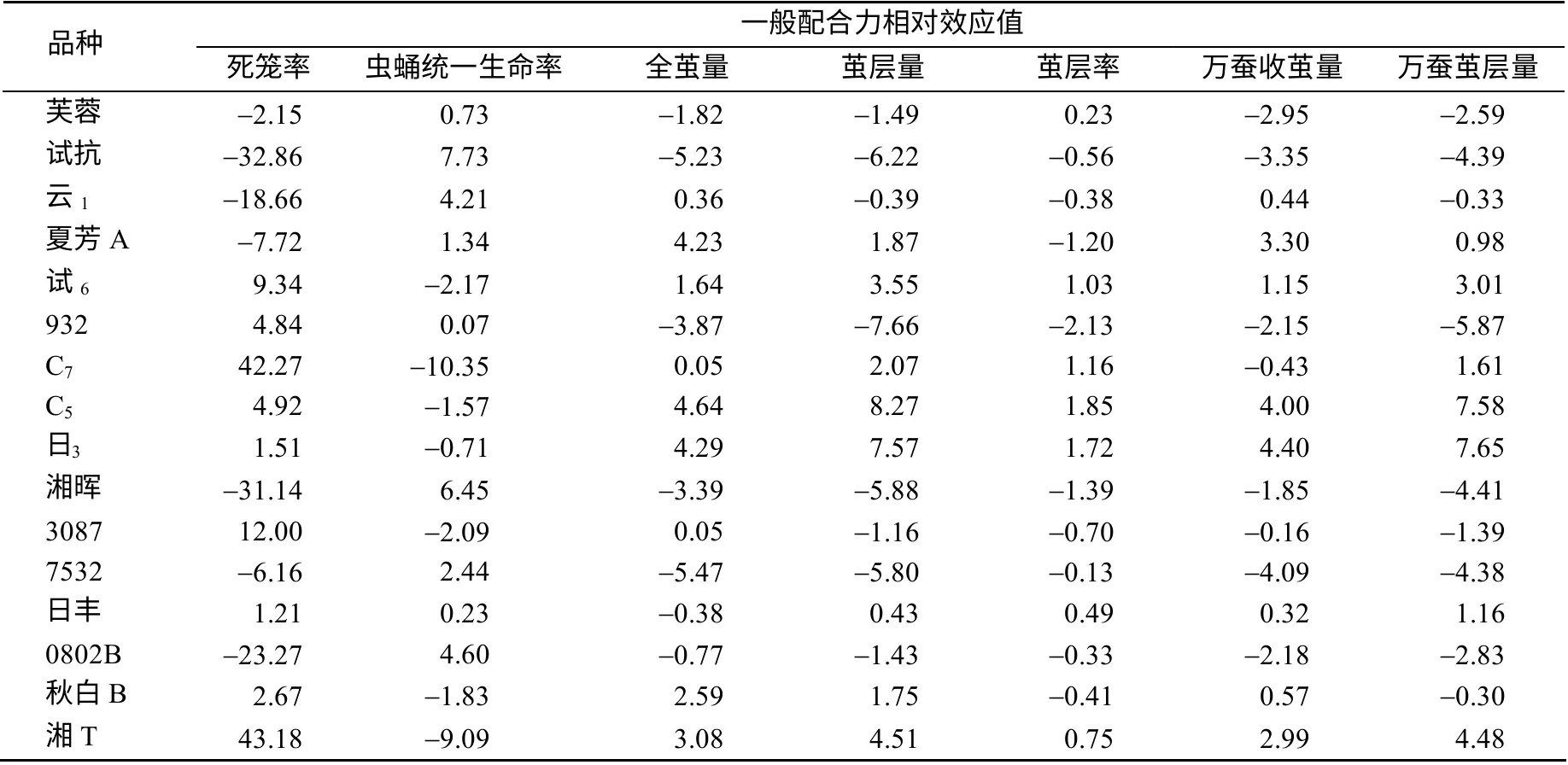
表2 各亲本7 个数量性状的一般配合力相对效应值Table 2 Estimates of the relative general combining ability effects for seven quantitative characters in parents
2.3 不同组合的特殊配合力效应
特殊配合力(SCA)是亲本品种在特定杂交组合中对杂交后代某一性状平均值的偏离。亲本品种的非累加效应(显性、超显性和上位效应)要依据具体组合而定。由表3 可知,组合间各性状的SCA 效应值差异很大,同一性状不同组合间SCA 效应值差异也很大,其中,日丰×夏芳A、3087×C7、0802B×C7、日3×C5、湘T×芙蓉、秋白B×试抗等6个组合各性状的SCA 效应值均在家蚕育种方向上有不同程度的提高;0802B×试抗、秋白B×云1、秋白B×C5、3087×芙蓉、日3×夏芳A、7532×试6、湘晖×C7等组合多个性状的SCA 也较高。
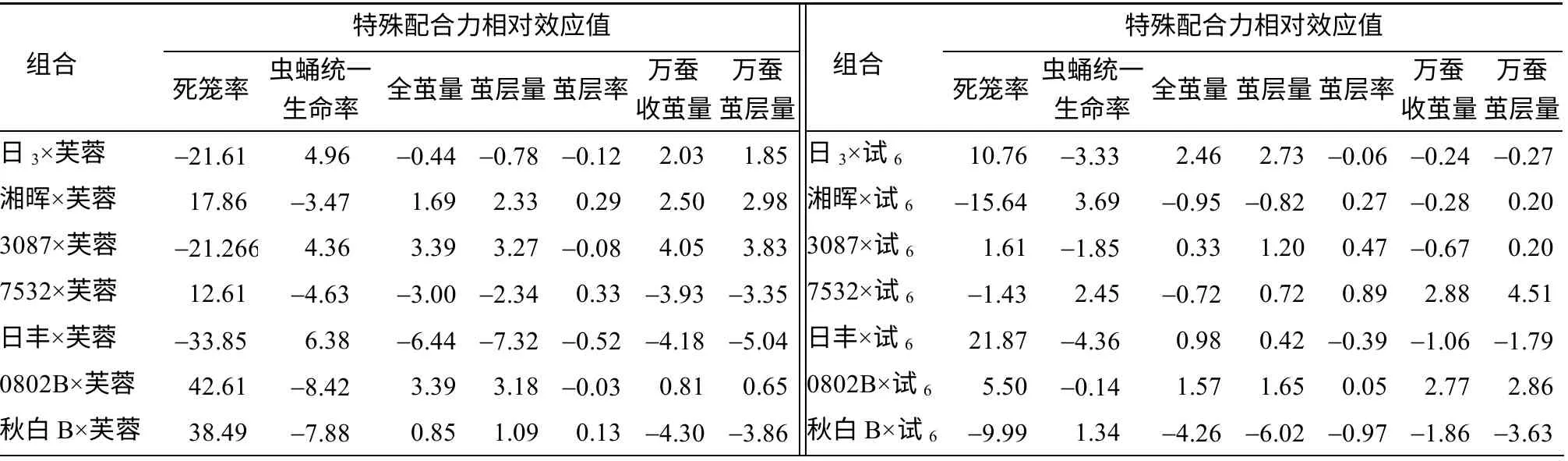
表3 64 个组合的7 个数量性状的特殊配合力相对效应值Table 3 Estimates of the relative specific combining ability effects for seven quantitative characters in 64 hybrid combinations
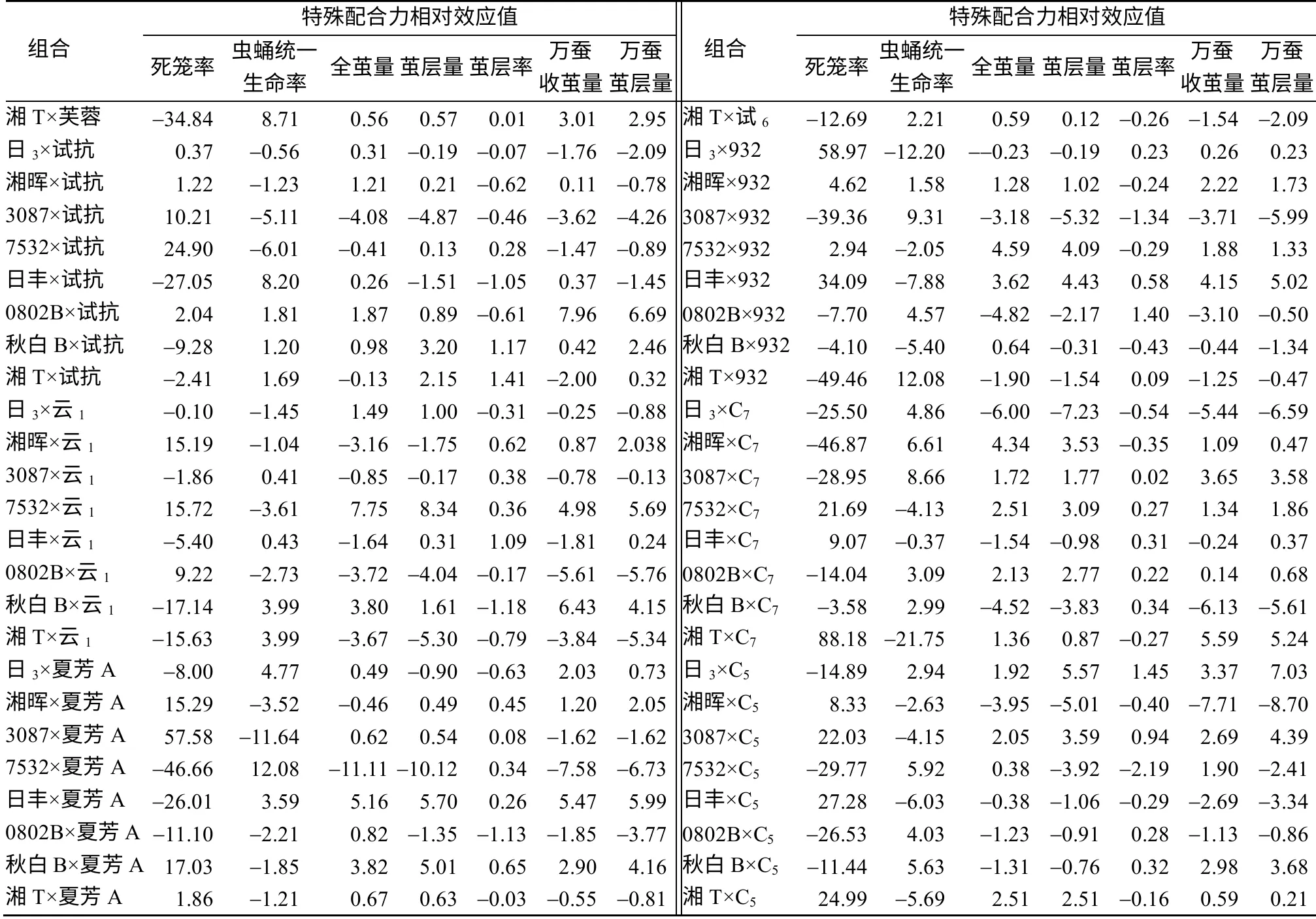
续 表
2.4 各性状的总配合力效应值
家蚕杂交组合的各性状表现既受双方亲本GCA 效应的影响,又受特定组合SCA 效应的制约。将父、母本的GCA 效应和杂交组合的SCA 效应按线性累加为总配合力(TCA)效应,可更直观、准确地判定杂交组合的优劣[4]。通过计算发现,日3×C5、日3×芙蓉、日3×云1、0802B×试6、0802B×C5、秋白B×C5等6 个组合各性状的TCA 效应值在育种目标方向上均有不同程度的提高(表4),秋白B×云1、日3×夏芳A、日3×试抗、日丰×夏芳A、日丰×云1、0802B×试抗、7532×云1 等组合多个性状的TCA 效应值较高,综合表现优良,其中,日3×C5丰产性状的TCA 效应值最高,0802B×试抗强健性状的TCA效应值最高。进一步对这些组合的亲本进行分析,发现利用GCA 表现优良的亲本进行杂交育种能明显提高子代的遗传增益,较易得到表现突出的强优组合,如日3、C5、云1等。综合分析强健性与丰产性的表现,日3×C5是最具有育成前途的强优组合。

表4 优良组合7 个数量性状的总配合力效应值Table 4 Estimates of the relative total combining ability effects for seven quantitative characters in the excellent hybrid combinations
2.5 各数量性状的群体配合力方差及其对杂交种的贡献率
为了进一步明确双亲及互作对杂交后代的影响,根据随机模型估算各性状的亲本基因型GCA 方差(V1P,V2P)与SCA 方差(V12P),进而估算出各 性状的GCA 方差与SCA 方差及其在基因型总方差(VT)中的份量,同时估算了在GCA 方差中父、母本所占的份量[5]。从数量遗传角度来看,GCA 主要由基因加性效应决定,遗传较为稳定,而SCA 是由基因的显性效应、上位效应及超显性效应等非加性效应决定。从表5 可以看出,7 个数量性状的GCA效应值均大于50%,说明这些性状中亲本基因的加性效应对杂种一代性状的形成起着主导作用。特别是茧层率的基因加性效应值达86.22%,两亲本的互作效应十分低。父、母本效应对7 个性状在杂交种F1中的表现是不同的,虫蛹统一生命率表现为母本P1的GAC 方差大,而茧层率表现为父本P2的GCA方差大,全茧量表现为父本P2的GCA 方差略大于母本P1,其他性状的GCA 方差大致相等。这与文献[6]的试验结果有一定的差异,这可能与本研究中的试验材料和试验环境与文献[6]中的试验材料(春用限性品种)及试验环境不同有关。

表5 7 个数量性状的基因型方差与父、母本及其互作对杂交种F1的遗传贡献率Table 5 The contribution ratio of the male,female and their interaction to the genotypic variance of the seven quantitative characters in F1 hybrids
2.6 各数量性状的遗传力估算
根据配合力方差分析结果,估算了各性状的广义遗传力与狭义遗传力(表6)。各性状的广义遗传力从大到小依次为茧层量、全茧量、茧层率、万蚕茧层量、万蚕收茧量、死笼率、虫蛹统一生命率;各性状的狭义遗传力从大到小依次为茧层率、茧层量、全茧量、万蚕茧层量、万蚕收茧量、死笼率、虫蛹统一生命率。除茧层率外,各性状广义遗传力与狭义遗传力的变化趋势基本相同。虫蛹统一生命率、死笼率、万蚕收茧量的广义遗传力低于50%,其环境方差较大,这些性状易受环境因素影响;全茧量、茧层量、茧层率、万蚕茧层量的广义遗传力达到60%以上,说明这些性状受环境因素影响较小。茧层率、茧层量、全茧量的狭义遗传力高于50%,其中茧层率与茧层量的均在70%以上,表明其性状由亲代遗传给子代的传递能力强;死笼率、虫蛹统一生命率的狭义遗传力低于30%,这与育种工作中生命力易受环境影响的实际相符。

表6 7 个数量性状的群体遗传力Table 6 Heritability of seven quantitative characters in the colony
3 结论与讨论
a.家蚕主要数量性状的遗传特点。本研究中有关家蚕强健性与丰产性的7 个主要数量性状的GCA 和SCA 方差均达到显著或极显著差异水平,表明这些性状的遗传受亲本基因的加性效应与非加性效应共同控制,但各性状的GCA 方差均大于SCA 方差,因此,这些性状的遗传效应主要是基因的加性效应。通过对群体的配合力方差及其对杂交种的贡献率进行分析,发现虫蛹统一生命率表现出了一定程度的母本效应,而茧层率表现出了较强的父本效应,这与夏秋蚕品种选育过程中以生命力强的多化性品种为母本,以丝量多、茧层厚的二化性品种为父本的亲本选择经验[7]相吻合;全茧量也表现出了较弱的父本效应,这与冯家新[8]关于家蚕选育的正反交试验结论一致,但他的研究中茧层率没有父本效应。遗传力是进行育种选择的重要参数[6]。本研究中,家蚕的茧层率、茧层量、全茧量的遗传力高,宜在选育的早期世代进行(这与文献[3]结果一致);死笼率、虫蛹统一生命率的狭义遗传力低,宜在显性和上位等效应消退后,遗传力有所提高的中后期世代进行单蛾饲养与定向选择,早期选择时应适当降低选择强度,扩大入选个体数和下一代参与收蚁蚕蛾数。本试验结果可作为目前家蚕品种经典选育方法[3]的参考依据。
b.配合力对家蚕遗传育种的利用价值。GCA 是评价亲本潜在育种能力的重要指标[10]。本研究中发现不同亲本在同一性状、同一亲本在不同性状上的GCA 效应差异很大。前人在家蚕或其他作物研究中也得到相似的结果[11-14]。本试验中的材料选择具有一定的地域代表性与遗传差异性。在各自不同的地理品系中,中系品种C5、日系品种日3的5 项丰产性状的GCA 效应值均最高,日3×C5、秋白B×C5、日3×夏芳A 等杂交组合也属于GCA 效应和SCA 效应均高的优良亲本。中系品种试抗、日系品种湘晖的强健性状的GCA 效应值最高,因此,对现行品种中生命力低的品种进行改造具有重要意义。中系品种932、日系品种7532 已作为成对亲本材料或单一亲本材料被广泛应用,并衍生出了一大批实用性家蚕品种。目前国内几乎所有日系育成品种均含有7532 的血缘[15],但在本次试验中,它们多个性状的GCA 效应明显偏低,表明这些骨干亲本通过各地深度改造,已形成了一批更加优良的地方品种,或各地通过自主创新获得了一批GCA 效应更高的基础材料。通过SCA 效应分析还发现,中系品种932、日系品种的7532 与其他大部分地方品种的互作效应并不突出,表明这些地方品种与当前普遍应用家蚕品种的遗传差异较大,具有育成遗传成分更加丰富、综合表现更加优良组合的潜力。
[1]司马杨虎,钱荷英,徐世清,等.家蚕对交系间的遗传距离及其一代杂交种的杂种优势研究[J].蚕业科学,2008,34(3):429-434.
[2]代方银,鲁成.家蚕基因资源持续保存的重要意义与我国的任务[J].中国蚕业,2001,22(3):5-12.
[3]向仲怀.家蚕遗传育种学[M].北京:农业出版社,1994:113-262.
[4]罗俊,周会,张木清,等.能源甘蔗主要经济和光合性状的遗传分析[J].应用与环境生物学报,2004,10(3):268-273.
[5]顾万春.统计遗传学[M].北京:科学出版社,2004:280-298.
[6]杨峻.家蚕数量性状的配合力和遗传力的分析[J].安徽农学院学报,1988(1):62-70.
[7]江苏省蚕业研究所蚕种研究室夏秋组.关于夏秋蚕品种选育工作的意见[C]//冯家新.家蚕育种选集.杭州:浙江大学出版社,2002:616-687.
[8]冯家新.家蚕育种方法的探索[J].蚕业科学,1980,6(3):159-166.
[9]杨明观.家蚕数量性状配合力与杂种优势研究[J].蚕业科学,1982,8(4):193-198.
[10]Griffing B.Concept of general and specific combining ability in relation to diallel crossing sestems[J]. Australian Journal of Biological Science,1956,9:463-493.
[11]Seidavi A.Analysis of combining ability for some parameters in iranian lines of silkworm Bombyx mori L.(Lepidoptera:Bombycidae) [J].Annals of Biological Research,2011,2(2):158-163.
[12]Darvishzadeh R,Alavi R.Genetic analysis of chloride concentration in oriental tobacco genotypes[J].Journal of Plant Nutrition,2011,34(7):1070-1078
[13]朱勇,向仲怀.家蚕主要数量性状的配合力与遗传力分析[J].蚕学通讯,1992(1):15-19.
[14]张玲,杨国涛,谢崇华,等.几个籼型杂交水稻光合特性的配合力研究[J].南京农业大学学报,2009,32(2):5-9.
[15]姚福广,卢筱芬.家蚕夏秋用新品种朝霞(7532)的育成[J].广西蚕业,1995(1):49-51.
