南阳黄牛肌肉发育过程中的MyoD和MEF2基因的表达变化研究
2012-07-14祁艳霞张小辉庞有志王玉琴
祁艳霞,张小辉,庞有志,王玉琴,廖 辉
(河南科技大学动物科技学院,河南洛阳471003)
牛肌肉的发育状况是影响其经济价值的重要因素,选育肌肉发育快、肉质好的牛品种是家畜育种的目标之一.肌肉发育受一系列基因的调控,因此,肌肉发育的基因表达调控一直受到广泛的关注.肌肉组织是胚胎发育过程中最早形成的组织之一.以肌肉发生调控因子(Myogenic regulatory factors,MRFs)为核心的信号传导网络在肌细胞分化过程中发挥关键的调控作用[1].MyoD(Myogenic differentiation 1)为MRFs家族成员之一,在骨骼肌细胞中表达,可激活下游肌肉特异表达基因的转录,抑制细胞生长,促进细胞分化,使很多非肌细胞转化为肌细胞[2].MEF2(Myocyte enhancer factor2)属于MADS(MCM1 agamous deficiens serum response factor)框转录因子家族,脊椎动物MEF2蛋白由 4种亚基组成,分别由 MEF2A,MEF2B,MEF2C和MEF2D基因编码.转录因子MEF2的DNA结合位点是一段保守的序列,该序列广泛存在于肌肉组织特异性表达基因的调控区,MEF2与MyoD基因家族成员具有协同作用,共同调节胚胎期的肌肉发育[3].动物出生后的肌肉发育以肌纤维肥大为主[4],同时,肌间脂肪的沉积速度逐渐加快[5].因此,本研究利用荧光实时定量PCR技术分析了南阳黄牛出生后3月龄到24月龄之间背最长肌组织中MyoD和MEF2基因的表达变化模式,为进一步深入研究MyoD和MEF2基因在动物出生后的肌肉发育调控过程中的作用提供基础.
1 材料与方法
1.1 组织样品的采集
选择发育正常的3,6,12,18,24 月龄的南阳黄牛各4头,在无菌条件下分别采集背最长肌组织,立即投入液氮中保存备用.
1.2 总RNA的提取及检测
用Takara公司的RNAiso plus试剂盒提取各组织总RNA.利用A260:A280确定RNA的纯度,通过0.8%的琼脂糖凝胶电泳检测RNA的完整性,根据18S rRNA的亮度计算总RNA的浓度,用DEPC处理的双蒸水稀释至1 g·L-1备用.
1.3 荧光实时定量PCR分析
分别将 3,6,12,18,24 月龄南阳牛背最长肌组织的总RNA各300 ng用DNAse(Promega)处理,以去掉残存的基因组DNA,用Takara公司的PrimeScriptTM1st strand cDNA Synthesis Kit试剂盒合成cDNA第1链,以此 cDNA第1链为摸板,用Takara公司生产的SYBR®Premix Ex TaqTMⅡ试剂盒进行荧光定量分析(Stratagene Mx3005P),总的反应体系为20 μL,其中模板 cDNA 第1链1 μL,10×SYBR®Premix Ex TaqTM2 μL,上、下游引物各0.7 μL,ddH2O 7.6 μL.反应程序为 95 ℃ 预变性1 min;然后94℃变性 30 s,退火 30 s,72 ℃延伸30 s,42个循环.对所有样本进行3个重复测定,并在每次试验时设阴性对照.本研究用到的引物序列信息见表1.

表1 荧光实时定量分析的引物信息表Table 1 The information of primers for real time PCR analysis
1.4 基因表达数据分析
参照 WINER 等[6]方法,用 2-ΔCt法测定目的基因mRNA的相对表达水平,以GAPDH基因为内标进行标准化.通过方差分析对不同发育时期背最长肌组织的基因表达进行显著性检验,采用BON法进行多重比较.
2 结果与分析
2.1 MyoD基因在牛肌肉发育过程中的表达变化
随着南阳黄牛月龄的增加,MyoD基因在背最长肌组织中的表达表现出先升高后降低的趋势(表2).MyoD基因在3月龄时表达量最低,随后表达量显著升高(P<0.05);在18月龄时表达量达到最高值,与其它月份之间差异均显著(P<0.05);24月龄时表达量显著降低(P<0.05).
2.2 MEF2基因在牛肌肉发育过程中的表达变化
脊椎动物MEF2蛋白由4种亚基组成,分别由 MEF2A,MEF2B,MEF2C,MEF2D 基因编码.随着南阳黄牛月龄的增加,各编码基因在背最长肌组织中的表达趋势也存在一定差异,见表2.MEF2A基因在南阳黄牛背最长肌组织中的表达也表现出先增加后降低的趋势,该基因在3月龄时表达量最低,在12月龄和18月龄时表达量显著升高(P<0.05),随后表达量下降,但差异不显著(P>0.05).MEF2B基因在背最长肌组织中的表达呈持续的上升趋势,24月龄时表达量最高.MEF2C基因在肌肉组织中的表达量相对较低,在肌肉发育过程中的表达变化幅度比较小,MEF2C基因在3月龄时表达量最低,18月龄时表达量最高,差异显著(P<0.05).MEF2D基因的表达情况与MyoD基因一致,表现出明显的先升高后降低趋势,18月龄和24月龄时表达量比3月龄时显著高(P <0.05).
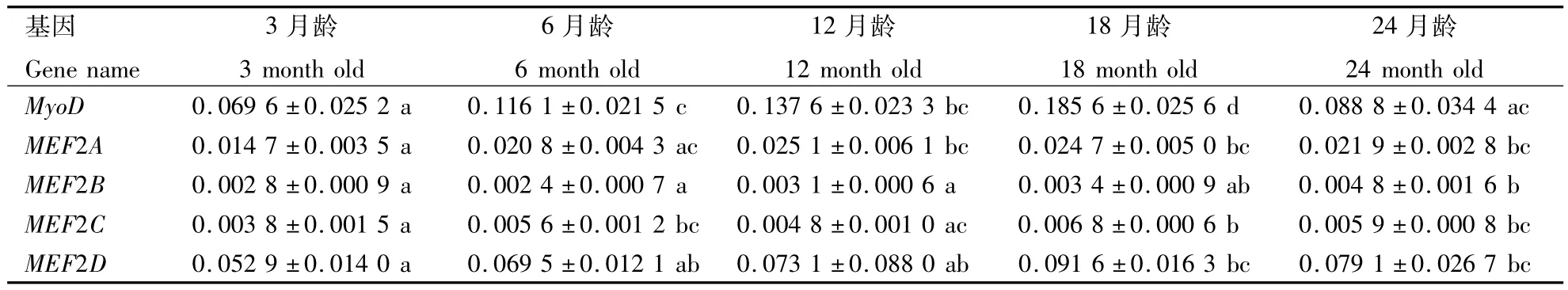
表2 MyoD和MEF2基因在南阳黄牛背最长肌发育过程中的表达变化Table 2 The expression of MyoD and MEF2 during the development of longissimus dorsi muscle in Nanyang cattle
3 小结与讨论
1)MyoD是调控肌肉生成的重要基因,其表达对维持肌细胞分化有重要作用,胚胎期MyoD基因缺失可导致成肌细胞的增殖和分化无法进行[7].肌细胞属于去分化能力的细胞,其数量在动物出生后基本恒定,肌肉细胞一旦受伤,则组织中的卫星细胞又可以再次分裂,生成新的肌肉细胞,MyoD在此过程中也发挥了重要的作用[8].MyoD基因在剧烈运动后的小鼠骨骼肌组织中表达水平上升,即表明该基因参与了损伤后骨骼肌细胞的修复过程[9].MyoD基因在出生后的动物肌肉组织中高表达,其表达水平与肌肉发育的丰满程度存在一定关系,长白猪肌肉组织中MyoD基因的表达水平极显著高于八眉猪[10].此外,MyoD基因的某些SNPs也影响肌肉的发育和胴体品质[11,12].这些研究表明,MyoD基因在多个方面对肌肉的发育进行调节,不仅是mRNA的表达量,而且蛋白的表达量和活性对肌肉发育也能发挥调控作用.
2)脊椎动物中目前已经发现4种类型的MEF2基因,分别由4个基因编码,而且这4个基因分别位于不同染色体上,每一个MEF2基因都有其独特的时空表达模式[13].MEF2基因通过启动子的选择及mRNA的选择性拼接可产生多种不同的MEF2蛋白质.许多骨骼肌和心肌中特异性表达基因的调控区都存在MEF2的结合位点,说明该基因在调节骨骼肌和心肌细胞系的发育过程中发挥重要的作用[14].MEF2转录因子主要促进I型肌纤维的生成[15].它最突出的功能是控制肌细胞分化过程中的基因转录,主要在骨骼肌、心肌和平滑肌的发育过程中发挥作用.MEF2与其它肌肉组织特异性表达的转录因子具有协同作用,共同调节肌细胞的分化.在骨骼肌中,MEF2就需要与MyoD家族成员协同作用来激活其它基因的转录[16].MEF2A在骨骼肌中的表达量比心肌组织中高[17].MEF2基因的表达对心肌细胞的正常分化和功能维持也是必要的,MEF2A和MEF2C基因的过度表达导致扩张性心肌症[18].MEF2A基因具有丰富的多态性,其中部分SNPs位点与胴体品质和胸肌发育有关[19].MEF2C基因缺乏时骨骼肌发育不良,说明MEF2C基因对肌纤维维护与成熟发挥重要作用[20].本研究发现,MEF2基因家族的4个成员在牛背最长肌组织的发育过程中表达情况各不相同,且不同的MEF2基因的基础表达水平存在比较大的差异.
MyoD和MEF2都是调控胚胎期肌肉细胞发育的重要基因,本研究发现这2个基因在出生后的牛肌肉组织中也具有比较高的表达量,而且每个基因的表达变化模式不同,表明这些基因的表达在动物出生后仍然受到精密的调控,可能在肌肉细胞的肥大或肌间脂肪的沉积上发挥重要的作用.
[1] 李永平,梁炳生.MyoD肌形成作用机制研究进展[J].国际骨科学杂志,2007,28(1):37-42.
[2] 袁婺洲,吴秀山.骨骼肌发育的分子遗传学[J].生命科学研究,2001,3(5):63-70.
[3] WU W,FOLTER S D,SHEN X,et al.Vertebrate paralogous MEF2 genes:origin,conservation,and evolution[J].Plos One,2011,6(3):17334 -17342.
[4] BUCKINGHAM M,BAJARD L,CHANG T,et al.The formation of skeletal muscle:from somite to limb[J].Anat,2003,202:59 -68.
[5] CHARLOTTE A M.Muscle development and obesity[J].Organogenesis,2008,4(3):158 -169.
[6] WINER J,JUNGC K S,SHACKEL I,et a1.Development and validation of real-time quantitative reverse transcriptase polymerase chain reaction for monitoring gene expression in cardiac myocytes in vitro[J].Analytical Biochemistry,1999,270(1):41-49.
[7] ALVES H J,ALVARES L E,GABRIEL J E,et al.Influence of the neural tube/notochord complex on MyoD expression and cellular proliferation in chicken embryos[J].Braz J Med Biol Res,2003,36:191 -197.
[8] ONUR K,JULIO J M,SHOKO Y,et al.Progenitors of skeletal muscle satellite cells express the muscle determination gene,MyoD[J].Dev Biol,2009,332:131 -141.
[9] 潘同斌,钱福鸿,韩天翼,等.大负荷运动后不同时相大鼠骨骼肌成肌调节因子MyoD、myogenin表达的变化[J].中国运动医学杂志,2011,30(10):926-930.
[10] 杨燕军,白 亮,庞卫军,等.不同猪品种肌肉组织FoxO1与MyoD基因mRNA的表达及其相关性分析[J].中国生物化学与分子生物学报,2008,24(3):257-261.
[11] 田 璐,许尚忠,岳文斌,等.MyoD基因对肉牛胴体性状影响的分析[J].遗传,2007,29(3):313-318.
[12] BIN F,SUNEEL K O,DORIAN J G,et al.Genomewide association study identifies loci for body composition and structural soundness traits in pigs[J].Plos One,2011,6(2):14726-14737.
[13] 郭新军.利用生物信息学方法对人类肌肉增强因子2(MEF2)的特性比较及进化分析[J].遗传,2011,33(9):975-981.
[14] GIRON M D,SEVILLANO N,VARGAS A M,et al.The glucose-lowering agent sodium tungstate increases the levels and translocation of GLUT4 in L6 myotubes through a mechanism associated with ERK1/2 and MEF2D[J].Diabetologia,2008,51(7):1285 -1295.
[15] KIM M S,FIELITZ J,MC ANALLY J,et al.Protein kinase D1 stimulates MEF2 activity in skeletal muscle and enhances muscle performance[J].Mol Cell Biol,2008,28(11):3600-3609.
[16] LIU N,REW H W,KIM Y,et al.An intragenic MEF2 dependent enhancer directs muscle specific expression of microRNAs 1 and 133[J].Proc Natl Acad Sci,2007,104(5):20844-20849.
[17] 高智晟,杨 龙,张冬杰,等.MEF2A基因在肌肉组织中的表达研究[J].中国农学通报,2009,25(4):21-23.
[18] XU J,GONG N L,BODI I,et al.Myocyte enhancer factors 2A and 2C induce dilated cardiomyopathy in transgenic mice[J].Biol Chem,2006,281:9152 -9162.
[19] 周 艳,刘益平,蒋小松,等.优质鸡MEF2A基因的SNPs检测及其与屠体性状的相关研究[J].畜牧兽医学报,2009,40(8):1164-1170.
[20] POTTHOFF M J,ARNOLD M A,ANALLY J,et al.Regulation of skeletal muscle sarcomere integrity and postnatal muscle function by MEF2C[J].Mol Cell Biol,2007,27(23):8143 -8151.
