盐胁迫下不同盐生植物渗透调节的生理响应1)
2012-07-02倪建伟张华新
武 香 倪建伟 张华新 刘 涛
(中国林业科学研究院,北京,100091)
我国的盐碱地总面积在0.33亿hm2以上,主要分布在滨海、东北、西北、黄河上中游和黄淮海平原地区在内的17个省份,其中具有农业利用潜力的盐碱荒地和盐碱障碍耕地面积近0.13亿hm2,利用潜力巨大[1]。改良和利用盐碱地有助于缓解我国的人口和环境压力,而筛选和推广耐盐碱植物是改良利用盐碱地的有效措施之一,因此,培育和筛选能适应高盐环境的盐生植物,利用生物措施改良盐碱土,对于改善盐碱地生态系统和提高土地资源利用率具有重要意义。
盐碱胁下植物受到渗透胁迫和离子毒害,高等植物通过调节无机离子比例、数量和积累细胞无毒的有机物(脯氨酸、可溶性糖等)来进行渗透调节以提高其耐盐性[2]。目前,植物的耐盐性研究主要集中在利用生长指标和几个生理指标进行耐盐能力综合评价和耐盐机理研究等方面[3-8],对于盐胁迫下耐盐机理不同的盐生植物以及同一属的不同种植物渗透调节响应的研究较少。本试验以3种柳树、3种柽柳和2种白刺为试材,研究其在不同浓度盐胁迫下渗透调节物质含量变化的差别,并分析这种差别与其耐盐机理的关系,为进一步了解渗透调节物质在植物耐盐过程中的作用和研究拒盐植物、泌盐植物、稀盐植物的耐盐性差别提供了理论依据。
1 材料与方法
试验在北京市昌平区半壁店村的大棚温室内进行。供试材料为竹柳(Salix(trade name))、沙柳(Salix psammophila)、乌柳(Salix cheilophila)、中华柽柳(Tamarix chinesis Lour).、甘蒙柽柳(Tamarix austromongolica)、刚毛柽柳(Tamarix hispida)、唐古特白刺(Nitraria tangutorum)、西伯利亚白刺(Nitraria Sibirica Pall.)的1年生苗木。2011年4月初,将试验材料定植在30 cm×24 cm的底部带托盘的塑料盆中。定植所用基质为V(腐殖质土)∶V(草炭)∶V(珍珠岩)=1∶3∶1混合均匀且质量相等,用多菌灵进行消毒。定植后进行常规管理。6月底,选择长势相对一致的苗木进行不同浓度的NaCl胁迫处理。试验采用完全随机区组设计,以不加NaCl为对照,共设4 个盐浓度处理,分别为 100、200、300、400 mmol/L,3次重复,每个小区8株苗木。采取分次浇灌的施盐方式。盐胁迫期间,定期定量浇水,漏于花盆托盘中的盐溶液或水倒回盆中,防止盐分流失。盐胁迫处理历时30 d。
采用磺基水杨酸比色法测定[9]脯氨酸,采用蒽酮比色法测定[9]可溶性糖,采用考马斯亮蓝法测定[9]可溶性蛋白,矿质元素参照王宝山[10]和於丙军[11]的方法,Na+和K+用原子吸收分光光度计法测定,Cl-用自动滴定仪测定。试验数据采用SPSS分析软件进行方差分析和多重比较。
2 结果与分析
2.1 盐胁迫对8个树种有机渗透调节物质的影响
30 d盐胁迫后,各树种叶片中的脯氨酸质量分数变化有所不同,除乌柳在400 mmol/L时的低于对照外,其他与对照相比均有不同程度的增加,盐胁迫对中华柽柳、刚毛柽柳、唐古特白刺、西伯利亚白刺的脯氨酸质量分数影响显著(P<0.05)(表1)。其中,3种柳树在低盐浓度胁迫下呈增加趋势,沙柳盐胁迫前后无明显差别,竹柳、沙柳、乌柳分别在400、100、200 mmol/L时达到最大值,比对照增加了13.65%、11.69%、12.95%;中华柽柳随盐浓度的上升呈先增后降的趋势,刚毛柽柳与盐浓度呈明显的正相关,甘蒙柽柳除在300 mmol/L时略有下降外基本呈增加趋势;唐古特白刺的脯氨酸质量分数随盐浓度上升而增加,西伯利亚白刺的变化与甘蒙柽柳相似。
盐胁迫30 d对竹柳和唐古特白刺叶片中可溶性糖质量分数的影响显著(P<0.05)(表2)。其中,3种柳树盐胁迫后相比对照均有不同程度的增加,沙柳和乌柳仅在高盐浓度下与对照有较大差异;3种柽柳除中华柽柳和刚毛柽柳在盐浓度为400 mmol/L时低于对照外,其他处理均高于对照,且随盐浓度增加的变化趋势相同,均先增后降;唐古特白刺在400 mmol/L时可溶性糖质量分数低于对照,西伯利亚白刺随盐质量分数的变化呈先增后降的变化趋势,与柽柳相似。甘蒙柽柳、竹柳和唐古特白刺的增幅最大,分别为 60.56%、54.32%和 41.08%。
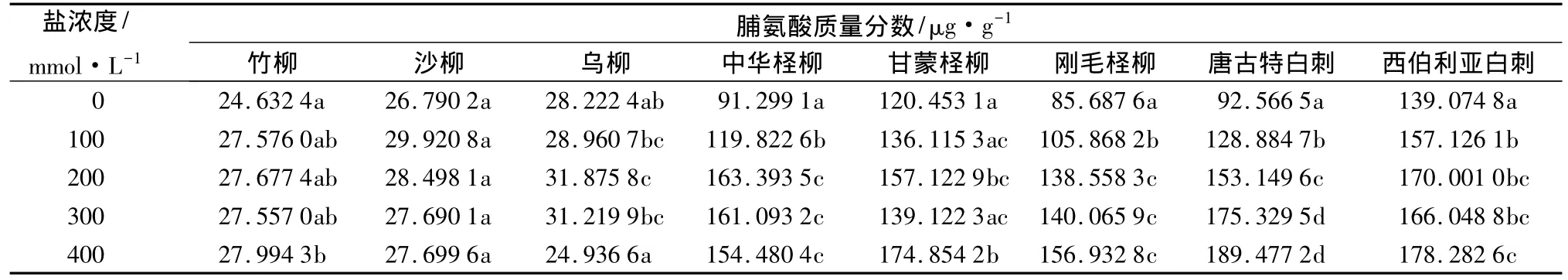
表1 不同浓度盐胁迫对叶片中脯氨酸质量分数的影响

表2 不同浓度盐胁迫对叶片中可溶性糖质量分数的影响
各树种盐胁迫后叶片中的可溶性蛋白质量分数的变化有所不同,盐胁迫对乌柳的影响不显著(P>0.05)(表3)。其中,竹柳和沙柳的变化趋势相同,除在300 mmol/L时略有下降外,基本呈增加趋势,高浓度盐胁迫对其影响较为明显,乌柳随盐浓度呈先增后降的趋势;盐胁迫下中华柽柳和刚毛柽柳的可溶性蛋白质量分数均低于对照,甘蒙柽柳在低盐浓度下明显增加,高盐浓度与对照含量相差不大;唐古特白刺的可溶性蛋白质量分数在200 mmol/L时达到最大,比对照增加了63.66%,西伯利亚白刺随盐浓度呈现明显的先增后降的趋势,在300 mmol/L时的含量最大,增幅为174.33%,在8个树种中增幅最大。总体来说,除中华柽柳和刚毛柽柳以外的6个树种在低浓度盐胁迫下叶片中积累可溶性蛋白。
2.2 盐胁迫对8个树种无机渗透调节物质含量的影响
盐胁迫后的Na+质量分数总体上是增加的,盐胁迫对乌柳、中华柽柳、甘蒙柽柳、唐古特白刺的叶,竹柳的茎以及乌柳、甘蒙柽柳的根中的Na+影响显著(P<0.05)(表4)。低盐浓度下,除刚毛柽柳和西伯利亚白刺的茎外,其他各树种的根茎叶中的Na+质量分数增加。3种柽柳的叶、中华柽柳和唐古特白刺的茎、中华柽柳的根均随盐浓度的增加呈先增后降的趋势。竹柳的叶以及竹柳、沙柳、乌柳、刚毛柽柳、唐古特白刺的根均与盐胁迫浓度呈正相关。3种柽柳和2种白刺的茎中Na+质量分数的变化种间差别较大。
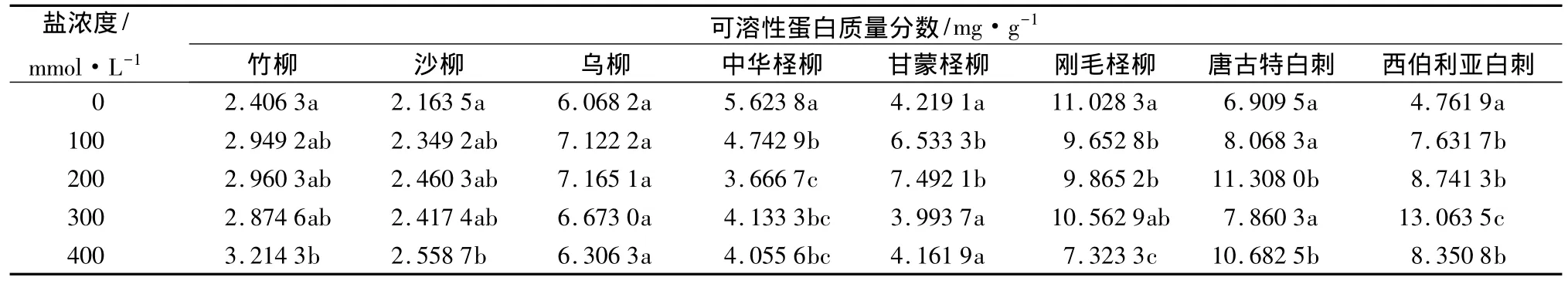
表3 不同浓度盐胁迫对叶片中可溶性蛋白质量分数的影响
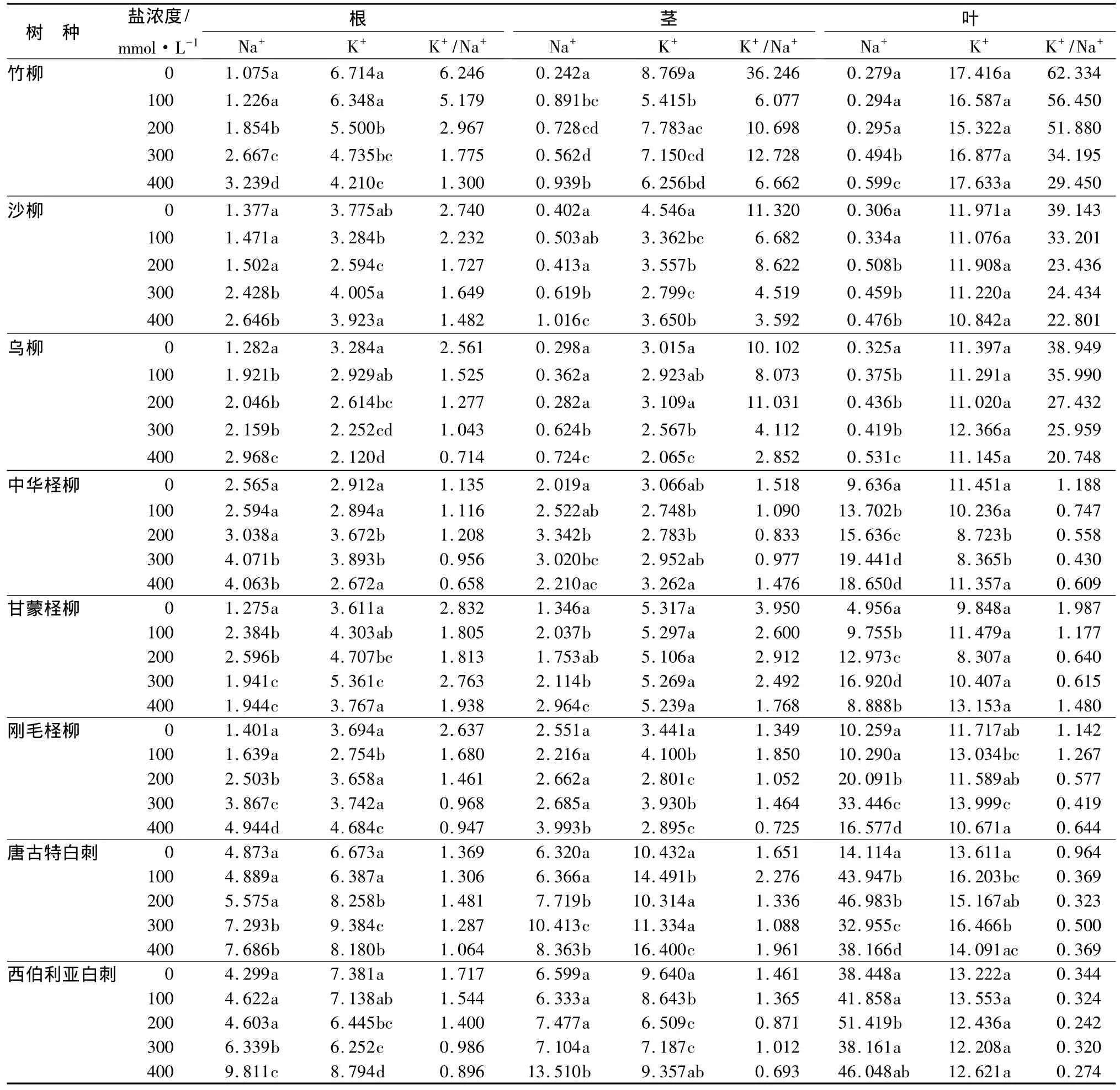
表4 不同浓度盐胁迫对8个树种Na+、K+质量分数及K+/Na+比值的影响
8个树种盐胁迫下各部位K+质量分数变化比较复杂(表4)。竹柳、沙柳、乌柳、甘蒙柽柳、西伯利亚白刺的叶和甘蒙柽柳的茎中的K+对盐胁迫的响应不显著(P>0.05),刚毛柽柳和沙柳的茎各盐浓度处理的K+质量分数均与对照差异显著(P<0.05)。沙柳、中华柽柳的叶,竹柳、沙柳、甘蒙柽柳、西伯利亚白刺的茎和竹柳、乌柳的根各浓度盐胁迫后的K+均低于对照,树种间的下降幅度差异较大。甘蒙柽柳的根和唐古特白刺叶中的K+质量分数则高于盐胁迫前。
盐胁迫后8个树种各部位的钾钠比除乌柳、刚毛柽柳、唐古特白刺的茎和中华柽柳、唐古特白刺的根在个别浓度盐胁迫下高于对照外,其他都小于盐胁迫前(表4)。竹柳、乌柳的叶和竹柳、沙柳、乌柳、刚毛柽柳、西伯利亚白刺的根的钾钠比均随盐浓度的升高而降低。3种柳树盐胁迫前后各部位的钾钠比叶>茎>根,柽柳和白刺根茎叶中的钾钠比叶中最小。
各树种不同部位的Cl-质量分数变化趋势及变化幅度是不同的,乌柳、中华柽柳的根,甘蒙柽柳的茎和中华柽柳、甘蒙柽柳、唐古特白刺的叶各浓度盐胁迫后的Cl-质量分数与对照差异显著(P<0.05)(表5)。盐胁迫后,竹柳、乌柳、中华柽柳、甘蒙柽柳、唐古特白刺的根和叶,甘蒙柽柳、刚毛柽柳的茎中的均高于对照。叶中增幅最大的是是刚毛柽柳和甘蒙柽柳,茎中的Cl-变化最大的是中华柽柳和刚毛柽柳,根中增加幅度最大的是乌柳和中华柽柳。白刺根茎叶中的Cl-质量分数大于柳树和柽柳。

表5 不同浓度盐胁迫对8个树种Cl-质量分数的影响
3 结论与讨论
盐胁迫下,植物通过合成和吸收积累渗透调节物质,维持高的细胞质渗透压,以保证细胞的正常生理功能抵御盐胁迫,脯氨酸、可溶性糖等是植物体内主要的渗透调节物质,这些物质的积累便于植物在高盐条件下对水分的吸收,以保证细胞的正常生理功能[12-14]。其中,脯氨酸的大量积累,能够降低细胞的水势,调节渗透平衡,避免细胞在盐胁迫下脱水死亡,但目前关于脯氨酸含量与植物抗盐性关系仍有争议,需作进一步的研究;可溶性糖可为有机物质的合成提供碳架和能量来源,起到稳定细胞膜的作用,是重要的渗透调节物质;可溶性蛋白也是一种重要的渗透调节物质,具有较强的亲水性,对提高植物的渗透势和抗逆性具有重要作用[15-18]。在本试验中,8个树种盐胁迫后叶片中积累脯氨酸,而属于拒盐植物的竹柳、沙柳和乌柳不论是脯氨酸整体质量分数还是盐胁迫后脯氨酸的最大增幅均低于属于泌盐植物的3种柽柳和稀盐植物的2种白刺,可见相比柽柳与白刺,脯氨酸在柳树渗透调节中发挥的作用较小,估计这种差别是由于柳树不同于柽柳与白刺的耐盐机理造成的,即不允许盐分进入体内或把进入体内的盐离子区域化到植物体安全部位而免受盐分伤害,但其具体相关性还需要进一步研究。可溶性糖质量分数除中华柽柳、刚毛柽柳、唐古特白刺在400 mmol/L时低于对照外,其他处理均高于对照,低盐浓度下8个树种均积累可溶性糖抵御盐胁迫。盐胁迫前后,柳树的可溶性糖均高于柽柳和白刺,增加幅度因树种不同而存在差异。3种柳树和2种白刺盐胁迫后叶片中的可溶性蛋白质量分数增加,柽柳种间差别较大,由试验可知可溶性蛋白不是甘蒙柽柳和刚毛柽柳的主要渗透调节物质。总体来说,不同植物在盐胁迫环境中用于渗透调节的主要有机渗透调节物质不同,这与植物的耐盐机理和种间差别存在一定的相关性。
盐胁迫下,植物体内矿质元素的变化与分布状况对植物的渗透调节能力和耐盐能力有非常重要的影响。根系吸收过程中高浓度的Na+和Cl-存在,干扰了植物对营养元素K+、Ca2+和N的吸收,对植物产生毒害作用,决定林木耐盐能力的关键之一在于林木对Na+、K+、Cl-等离子的吸收的调节能力,即限制Na+和Cl-进入体内,选择性地吸收K+,才能提高其耐盐能力[19]。本试验中,8个树种盐胁迫后各部位Na+质量分数增加,3种柳树根中的Na+质量分数最大,茎和叶中的Na+差别与柳树品种有关,柽柳叶中最大,白刺含量最高的部位也是叶,茎略大于根,由此可见,耐盐机理不同的植物根系对Na+离子的选择性吸收及其在各部位的分配是不同的。K+质量分数变化比较复杂,总的来说,盐胁迫使3种柳树的K+减少,柽柳和白刺各部位的K+变化品种间的差异较大,从树种的三个部位来看,叶中的K+明显高于根和茎。8个树种各部位的Cl-质量分数总体来说是增加的,叶片中的Cl-高于根和茎。盐胁迫后8个树种的钾钠比除根茎的个别处理高于对照外基本都是随着盐浓度的升高而降低的,可见Na+的吸收抑制了植物对K+的吸收,3种柳树钾钠比叶>茎>根,3种柽柳不同部位的钾钠比差别较小,2种白刺叶中钾钠比最小,根和茎差别较小,这可能是由于拒盐植物、泌盐植物、稀盐植物的不同耐盐机理影响无机渗透调节物质在植物各器官、组织及细胞上的区域化分布相关。试验中白刺根茎叶中的Na+和Cl-含量明显大于柽柳与柳树,这是因为白刺属于稀盐植物,在NaCl胁迫下,主要通过从外界吸收盐离子,即利用液泡积盐进行渗透调节,而属于拒盐植物的柳树和属于泌盐植物的柽柳则是分别通过拒绝盐分进入和利用盐腺与盐囊泡降低体内的盐分水平。
渗透调节是植物在盐渍环境中抵御盐胁迫的重要生理机制,渗透调节物质复杂多样,不同盐生植物因耐盐机理、种间差异等原因,其用于渗透调节的主要渗透调节物质有所不同,同一渗透调节物质的重要性也存在差别,因此关于拒盐植物、稀盐植物和泌盐植物耐盐机理与渗透调节机制的关系以及种间差异对植物渗透调节的影响,还需要结合同一耐盐机理的其他树种和更多的渗透调节指标如甜菜碱、多元醇、有机酸等,做更深入的研究。
[1]刘永信,王玉珍.盐碱地区域化种植耐盐植物可行性研究[J].宁夏农林科技,2011,52(5):49-50.
[2]许详明,叶和春,李守明.植物抗盐机理的研究进展[J].应用与环境生物学报,2001,6(4):379-387.
[3]谢福春,张文婷,刘富强,等.土壤盐胁迫对海州常山生理生化特性的影响[J].江西农业大学学报,2008,30(5):839-844.
[4]刘炳响,梁海永,李子敬,等.不同盐碱条件下白榆器官中K+、Na+、Ca2+和 Mg2+分布特征[J].西北林学院学报,2008,23(5):7-11.
[5]Akihiro Ueda,Yuko Yamamoto Yamane,Tetsuko Takabe.Salt stress enhances proline utilization in the apical region of barley roots[J].Biochemical and Biophysical Research Communications,2007,355:61-66.
[6]张华新,宋丹,刘正祥.盐胁迫下11个树种生理特性及其耐盐性研究[J].林业科学研究,2008,21(2):168-175.
[7]孙海菁,王树凤,陈益泰.盐胁迫对6个树种的生长及生理指标的影响[J].林业科学研究,2009,22(3):315-324.
[8]张华新,刘正祥,刘秋芳.盐胁迫下树种幼苗生长及其耐盐性[J].生态学报,2009,29(5):2263-2271.
[9]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[10]王宝山,赵可夫.小麦叶片中Na、K提取方法的比较[J].植物生理学通讯,1995,31(1):50-52.
[11]於丙军,罗庆云,曹爱忠,等.栽培大豆和野生大豆耐盐性及离子效应的比较[J].植物资源与环境学报,2001,10(1):25-29.
[12]薄鹏飞,孙秀玲,孙同虎,等.NaCI胁迫对海滨木槿抗氧化系统和渗透调节的影响[J].西北植物学报,2008,28(1):113-118.
[13]郝峰鸽,周俊国,周秀梅.NaCI胁迫对喜树幼苗生长和叶片生理特性的影响[J].东北林业大学学报,2010,38(1):18-19.
[14]杨涓,许兴.盐胁迫下植物有机渗透调节物质积累的研究进展[J].宁夏农学院学报,2003,24(4):86-91.
[15]郭媛媛,黄大庄.闫海霞.NaCI胁迫对黄顶菊生长及生理生化的影响[J].北华大学学报,2011,12(3):341-345.
[16]Heuer B.Influence of exogenous application of proline and glycinebetaine on growth of salt stressed tomato plants[J].Plant Science,2003,165:693-699.
[17]Eva Mari A,Mecafery S,Anderson J M.Photoinhibition and DI protein degradation in peas acclimated to different growth irradiances[J].Plant Physiology,1993,103:599-626 .
[18]王劲,杜世章,刘君蓉.植物耐盐机制中的渗透调节[J].绵阳师范学院学报,2006,25(5):56-61.
[19]汪贵斌,曹福亮,游庆方,等.盐胁迫对4树种叶片中 K+和Na+的影响及其耐盐能力的评价[J].植物资源与环境学报,2001,10(1):30-34.
