乌桕秋叶转色前后生理特性及超微结构的变化1)
2012-06-13倪竞德黄利斌
张 敏 钱 猛 倪竞德 黄利斌
(江苏省林业科学研究院,南京,211153) (南京农业大学) (江苏省林业科学研究院)
乌桕(Sapium sebiferum(L.)Roxb.)为大戟科(Euphorbiaceae)乌桕属植物,是集能源、药用、材用、观赏为一体的多用途树种。作为长江中下游地区重要的景观树种之一,乌桕季相变化丰富,秋叶由绿变黄、变红或变紫,极具观赏价值。目前,有关乌桕开发利用[1-3]、抗逆机理[4-5]以及繁育[6-8]的研究较多,而未见其叶片呈色机理的报道。深入了解乌桕叶色变化的生理生化基础,寻求其叶色变化的原因及影响其呈色的相关因素,对提高乌桕的观赏价值和进行新品种选育具有重要的理论意义和参考价值。
本研究以乌桕秋叶为试材,对比研究了黄、红两个单株的叶片在秋季变色前后色素、氮元素、淀粉、糖等生理指标的差异及其相关性,还对叶绿体超微结构变化进行了分析,以探讨乌桕秋叶的呈色规律,为选育秋季叶色纯正的优良观赏株系提供理论依据。
1 材料与方法
在连续3 a叶色调查的基础上,选取树龄13 a、叶色性状稳定、生长健壮、长势一致、秋季转色后叶色分别为黄色和红色的乌桕单株进行本次试验。取样地点位于江苏省林业科学研究院大门口、宁丹路旁,取样时间分别为2010年9月20日(变色前)和11月12日(变色后)10:00。黄色单株和红色单株分别取样,以变色前绿色叶片为对照,取样部位及朝向基本相同。
生理生化指标测定:叶绿素质量分数测定采用丙酮乙醇混合液法[9],花色素苷质量分数测定采用盐酸乙醇浸提法[10],全氮质量分数测定采用凯氏定氮法[11],葡萄糖、果糖、蔗糖质量分数采用Roche公司(Darmstadt,Germany)Sucrose/D-Glucose/D-Fructose试剂盒测定,淀粉质量分数采用Roche公司(Darmstadt,Germany)Starch试剂盒测定。各项检测重复3次,结果用SPSS13.0系统软件进行数据统计分析。
叶绿体超微结构观察:取鲜叶洗净、擦干,快速切取1 ~2 mm2,放入 2.5% 戊二醛溶液固定,0.1 mol·L-1磷酸缓冲液冲洗,1%锇酸固定。再用0.1 mol·L-1磷酸缓冲液冲洗。30%、50%、70%、80%、90%、100%丙酮系列脱水,然后包埋剂逐级渗透,包埋,聚合后用瑞典LKB-V型超薄切片机进行超薄切片,铀铅染色,在日立H-600型透射电子显微镜下观察并拍照[12]。
2 结果与分析
2.1 乌桕秋叶变色前后色素质量分数变化
黄、红2种类型的乌桕秋叶中总叶绿素、类胡萝卜素、叶绿素a/b及花色素苷质量分数见表1。变色前后,2种类型秋叶中叶绿素质量分数及比值的差异较大。绿叶中总叶绿素质量分数分别为1.61、1.69 mg·g-1,均显著高于其变色后黄叶(0.28 mg·g-1)和红叶(0.33 mg·g-1)的质量分数(P<0.01);但变色后黄叶和红叶中总叶绿素质量分数无显著差异(P<0.05)。叶绿素主要有叶绿素a和叶绿素b两种,前者呈蓝绿色,后者呈黄绿色[13]。如表1所示,变色前叶绿素a和b之比略高于3;而变色后叶绿素a和b之比分别降低为1.95和1.38,说明叶绿素b的相对质量分数有所升高。
类胡萝卜素是一类天然有色物质,是叶绿素的伴生物,在秋叶显色中具有重要作用[14]。从表1的结果可以看出,黄色秋叶类胡萝卜素质量分数显著升高,比变色前绿叶提高了17.50%;而红叶单株变色前后,叶片中类胡萝卜素质量分数无显著差异。
秋季变色前后,黄叶单株叶片花色素苷质量分数变化不大;而红叶中花色素苷质量分数极显著升高,是绿叶的13.9倍,是黄叶的3.60倍。方差分析结果表明,红叶中花色素苷质量分数与绿叶和黄叶相比,均达到极显著差异水平(P<0.01),说明花色素苷质量分数的高低是决定乌桕红色秋叶着色的主导因素之一。
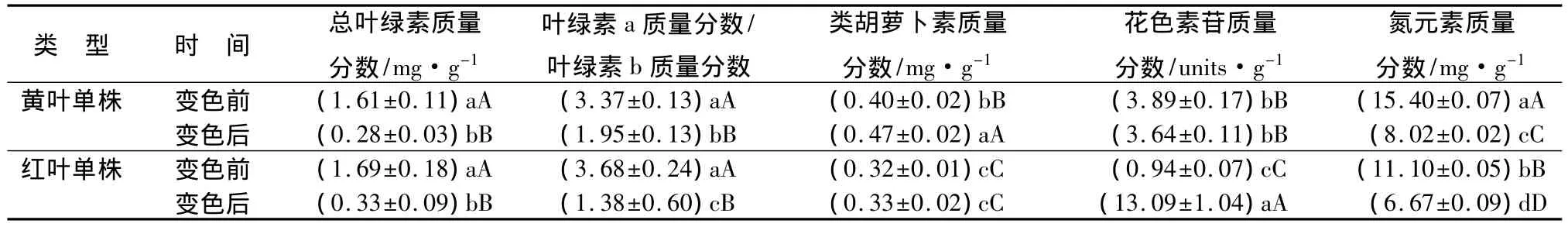
表1 乌桕秋叶变色前后色素及氮元素质量分数变化
2.2 不同单株叶片氮元素质量分数的比较
有研究表明,叶色差异与植物的氮素营养状况密切相关[15]。从表1中可以看出,乌桕秋叶在变色后氮元素质量分数明显降低。黄叶和红叶中氮元素质量分数分别比变色前绿叶降低27.92%和15.75%。其中,红叶中的氮元素质量分数最低,方差分析结果表明,不同颜色叶片中氮元素质量分数差异极显著(P<0.01)。
2.3 乌桕秋叶变色前后糖和淀粉质量分数的变化
糖是植物体内碳水化合物运输的主要形式,作为花色素苷的前体物质,在其合成过程中起重要作用[10]。从表2可以看出,乌桕叶片在秋季转色后,葡萄糖质量分数极显著降低(P<0.01)。黄色秋叶中果糖和蔗糖质量分数分别比变色前绿叶降低了51.49%和 76.01%,差异极显著(P<0.01)。红色单株的叶片在秋季转色后,果糖质量分数显著升高,是变色前绿叶的1.25倍(P<0.05);蔗糖质量分数也有所提高,但方差分析结果表明差异不显著。
淀粉是碳的主要储存物质,其质量分数在很大程度上影响着树木的生长及对环境的响应[16]。本研究中,黄色秋叶和红色秋叶中淀粉质量分数均比变色前显著降低(P<0.01)(表2),说明在叶片衰老过程中,光合产物储备减少。

表2 乌桕秋叶变色前后非结构性碳水化合物质量分数的变化 mg·g-1
植物叶片中非结构性碳水化合物(TNC)是碳吸收(光合同化)与碳消耗(生长与呼吸消耗)关系的一种量度,反映了可供植物生长利用的物质水平[17]。本研究结果表明,乌桕秋叶变色后非结构性碳水化合物质量分数降低,不同颜色由高到低排序为:绿叶、红叶、黄叶,方差分析结果表明,差异达显著水平(P<0.05)。
2.4 色素与相关生理指标的相关性分析
相关性分析表明(表3),总叶绿素质量分数与氮、葡萄糖、淀粉质量分数呈显著或极显著正相关;类胡萝卜素质量分数与果糖质量分数呈极显著负相关;花色素苷质量分数与氮元素、葡萄糖、淀粉质量分数呈负相关,与果糖质量分数呈正相关,但均未达到显著水平;花色素苷和叶绿素的比值与葡萄糖质量分数呈显著负相关关系。

表3 色素与全氮及非结构性碳水化合物质量分数的相关系数
2.5 乌桕秋叶变色前后叶绿体超微结构分析
利用透射电镜分别对变色前绿叶、变色后黄叶和红叶的叶肉细胞进行观察。结果发现黄叶单株的秋叶变色前,单位细胞内叶绿体数较多,叶绿体沿细胞壁边缘分布,形状呈纺锤形或椭圆形,叶绿体膜完整,界限清晰。叶绿体中基质物质浓厚,淀粉粒丰富,有少量嗜锇颗粒(图1-A)。黄色秋叶叶绿体数目变少,形状不规则,体积增大,被膜边缘模糊,有的破裂;基质片层疏松、不清晰,叶绿体中未见淀粉粒,嗜锇颗粒增多(图1-B)。
红叶单株变色前叶肉细胞的叶绿体呈椭圆形,数量多,结构完整,基质片层及基粒片层清晰可见。类囊体系统发育完善,类囊体中不仅有大量的基质片层,而且基粒较多,基粒垛叠较厚,基粒片层与基质片层界限明显,可以观察到淀粉粒,但数量较少(图1-C)。变为红色后,大部分细胞器已经解体,基质中出现囊泡,个别细胞中能够观察到正在解体退化的基粒片层残片。嗜锇颗粒大量增加,聚集在一起,基质中无淀粉粒存在(图1-D)。

图1 乌桕秋叶叶肉细胞超微结构
3 结论与讨论
色素质量分数、内含物种类以及细胞结构等因素均能影响植物叶片的呈色[18]。色素是彩叶植物叶片呈色的物质基础,其中叶绿素、类胡萝卜素和花色素苷是存在于叶片中的主要色素,其质量分数与比例是决定叶片呈色的主要因素[19]。乌桕秋叶在转色过程中,不同种类的色素处于动态变化中,变色前叶绿素质量分数高,类胡萝卜素和花色素苷质量分数较低,叶片呈绿色;随着秋季气温降低,叶绿体开始解体,总叶绿素质量分数和叶绿素a质量分数显著降低。黄叶类型乌桕秋叶中类胡萝卜素质量分数大幅升高,叶片呈黄色;红叶类型乌桕秋叶在叶绿素质量分数降低的同时,花色素苷合成加快,质量分数急剧升高,叶片呈红色。不同颜色乌桕秋叶中的色素质量分数存在显著差异,红色单株叶片中花色素苷的质量分数远高于黄色单株,成为乌桕秋叶呈现红色的主要原因。
已有研究表明,叶色与全氮质量分数的变化是同步的,叶色实际上是叶片氮化合物状况的外在表现[20]。在本试验中,氮元素质量分数与总叶绿素质量分数呈显著正相关,与花色素苷质量分数呈不显著负相关。此结果与 Schaberg et al.[21]对糖槭(Acer saccharumMarsh.)秋叶的研究相一致。
对红色起主导作用的花色素苷是由花青素和糖组成的糖苷,花青素又是在糖代谢的基础上形成的[22]。还有报道指出,糖可作为信号物质诱导花色素苷合成基因的表达和调节酶的活性[23],因此,糖质量分数同乌桕秋叶显色密切相关。本研究发现,红色秋叶中果糖和蔗糖质量分数高于变色前绿叶和黄色秋叶,表明果糖和蔗糖对乌桕叶片中花色素苷的累积有明显的促进作用。葡萄糖质量分数与总叶绿素质量分数呈极显著正相关,而与花色素苷质量分数呈负相关,分析原因可能是黄色和红色秋叶在衰老过程中叶绿素质量分数降低,光合作用减弱,导致光合产物葡萄糖的积累受到影响。
碳水化合物是光合作用的主要产物,是树木生长代谢过程中重要的能量来源。叶片同化的光合产物一部分用于构建各种组织和器官,称为结构性碳水化合物,如木质素、纤维素等;另一部分用于生理代谢活动,称为非结构性碳水化合物(TNC),包括葡萄糖、蔗糖、果糖、淀粉等[24]。已有研究表明非结构性碳水化合物不仅在植株生命代谢中发挥重要作用,还是植物适应环境的信号物质[25-26]。乌桕秋季变色后,非结构性碳水化合物质量分数显著降低,相关性分析表明,其质量分数与叶绿素质量分数呈正相关,与类胡萝卜素质量分数呈负相关。可能是由于黄色和红色秋叶中光合色素质量分数降低,光合产物生成较少;同时类胡萝卜素和花色素苷合成的基础物质是糖类物质,其生成过程中也消耗了大量的非结构性碳水化合物。花色素苷的累积受激素、酶、光、pH值、温度等多种因素的影响[27],可能是本试验中其与可溶性碳水化合物的相关性不明显的原因之一。糖代谢与叶片呈色密切相关[28-29],但其作用机制还有待于更深入细致的研究。
叶绿体是叶片进行光合作用的细胞器,也是植物叶片衰退进程中变化最灵敏的器官[30]。本研究发现,乌桕秋季叶色改变前后,叶绿体数量、形状以及基粒片层的结构均发生了明显变化,红色秋叶的内部结构比黄色秋叶更趋于简单。叶绿体中的淀粉粒是细胞碳水化合物存在的主要形式之一。从观察结果来看,绿叶中淀粉粒数量较多,体积较大,说明其叶绿体保持着活跃的光合作用功能。而叶色变黄或变红后,淀粉粒数量减少、体积减小,甚至消失,其原因可能是当叶绿体解体退化时,由于类囊体片层的减少,光合效率大大降低,淀粉的合成变慢,但水解速度加快,于是淀粉粒越来越小,直至最终消失[31]。乌桕秋叶变色后,嗜锇颗粒大量增加,并聚集,这种变化与衰老叶片中退化叶绿体的结构特点相似[32]。
总之,乌桕秋季叶色的变化是在多种因素的综合作用下完成的,最直接的原因是色素的成分和比例发生改变,而相关生理指标和叶绿体结构的变化都是秋叶呈色的重要影响因素。
[1]王洪庆,赵春阳,陈若芸.乌桕叶化学成分研究[J].中国中药杂志,2007,32(12):1179-1181.
[2]邹晓霞,杨江科,刘云,等.复合酶法制备乌桕梓油生物柴油工艺优化[J].中国油脂,2010,35(6):33-37.
[3]彭旦明,彭镇华.枫杨、乌桕对钉螺毒性的研究[J].应用生态学报,1995,6(3):301-304.
[4]蔡金峰,曹福亮,汪贵斌,等.淹水胁迫对乌桕幼苗叶片活性氧代谢的影响[J].福建林学院学报,2010,30(1):51-55.
[5]曹福亮,蔡金峰,汪贵斌,等.淹水胁迫对乌桕生长及光合作用的影响[J].林业科学,2010,46(10):57-61.
[6]金雅琴,何锐,张纪林,等.乌桕1年生播种苗生长规律[J].东北林业大学学报,2010,38(9):10-12.
[7]李建科,杨静慧,刘太林,等.野生乌桕的组培和再生[J].北方园艺,2010(13):164-166.
[8]郄亚微,徐有明,李学琴,等.能源林树种乌桕茎段离体培养与快速繁殖[J].东北林业大学学报,2010,37(12):8-9,42.
[9]张宪政.作物生理研究法[M].北京:农业出版社,1990,142.
[10]陈继卫,沈朝栋,贾玉芳,等.红枫秋冬转色期叶色变化的生理特性[J].浙江大学学报:农业与生命科学版,2010,36(2):181-186.
[11]王渊,黄敬峰,王福民,等.油菜叶片和冠层水平氮素含量的高光谱反射率估算模型[J].光谱学与光谱分析,2008,28(2):273-277.
[12]袁丽钗,李雪平,彭镇华,等.菲白竹组培苗白化、绿化突变体的超微结构及15个叶绿体编码基因的表达[J].植物学报,2010,45(4):451-459.
[13]王忠.植物生理学[M].北京:中国农业出版社,2000:66.
[14]陶俊,张上隆,张良诚,等.柑橘果皮颜色的形成与类胡萝卜素组分变化的关系[J].植物生理与分子生物学学报,2003,29(2):121-126.
[15]王绍华,吉志军,刘胜环,等.水稻氮素供需差与不同叶位叶片氮转运和衰老的关系[J].中国农业科学,2003,36(11):1261-1265.
[16]Ericsson T,Rytter L,Vapaavuori E.Physiology of carbon allocation in trees[J].Biomass and Bioenergy,1996,11(2/3):115-127.
[17]潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物的生理生态学研究进展[J].植物学通报,2002,19(1):30-38.
[18]朱书香,杨建民,王中华,等.4种李属彩叶植物色素含量与叶色参数的关系[J].西北植物学报,2009,29(8):1663-1669.
[19]赵昶灵,郭华春.植物花色苷生物合成酶类的亚细胞组织研究进展[J].西北植物学报.2007,27(8):1695-1701.
[20]王绍华,曹卫星,王强盛,等.水稻叶色分布特点与氮素营养诊断[J].中国农业科学,2002,35(12):1461-1466.
[21]Schaberg P G,van den Berg A K,Murakami P F,et al.Factors influencing red expression in autumn foliage of sugar maple trees[J].Tree Physiology,2003,23(5):325-333.
[22]李云飞,李彦慧,王中华,等.土壤干旱胁迫对紫叶矮樱叶片呈色的影响[J].生态学报,2009,29(7):3678-3684.
[23]赵玉辉,郭印山,李作轩.果实花青素研究进展[J].北方园艺,2006(3):46-47.
[24]印婧婧,郭大立,何思源,等.内蒙古半干旱区树木非结构性碳、氮、磷的分配格局[J].北京大学学报,2009,45(3):519-527.
[25]Gibson S I.Plant sugar-response pathway.Part of a complex regulatory web[J].Plant Physiol,2000,124(4):1532-1539.
[26]潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物的生理生态学研究进展[J].植物学通报,2002,19(1):30-38.
[27]Tomas-Barheran F A,Robins R J.Phytoehemisty of fruits and vegetables[M].Oxford:Clarendon Press,1999:29-49.
[28]唐前瑞,陈德富,陈友云,等.红檵木叶色变化的生理生化研究[J].林业科学,2006,42(2):111-115.
[29]胡静静,沈向,李雪飞,等.黄连木秋季叶色变化与可溶性糖和矿质元素的关系[J].林业科学,2010,46(2):80-86.
[30]魏晓东,陈国祥,徐艳丽,等.银杏叶片衰老过程中光合生理特性及叶绿体超微结构的变化[J].植物研究,2008,28(4):433-437.
[31]范燕萍,李慧玲,李浩健.几种花叶线艺兰叶片色斑色素组成和叶绿体超微结构研究[J].华南农业大学学报,2006,27(2):8-12.
[32]雷华,陈国祥,高志萍,等.两优培九剑叶叶绿体衰退进程中的细胞生物学特性初探[J].中国水稻科学,2008,22(3):279-284.
