拟南芥新型脂转移蛋白AtDHyPRP1的亚细胞定位及其对灰霉菌的抗性
2012-02-26张晨李岚徐子勤
张晨,李岚,徐子勤
西北大学生命科学学院 陕西省生物技术重点实验室 西部资源与现代生物技术省部共建教育部重点实验室,陕西 西安710069
脂转移蛋白 (Lipid transfer protein,LTP) 广泛存在于动物、植物和微生物中,它们对脂类分子的亲和作用没有选择性,所以也被称为非特异性脂转移蛋白 (Non-specific lipid transfer protein,nsLTP)。LTP蛋白最重要的特点是含有8个位置高度保守的半胱氨酸,可以形成4个二硫键。它们的三级结构十分稳定,中间具有一个疏水性空穴,可以转运多种类型的脂质分子[1]。越来越多的证据显示 LTP参与了植物的抗病反应,它们能够抑制病原体的生长,并产生系统性获得抗性 (Systemic acquired resistance,SAR)[2-3],已被归类到病程相关蛋白(Pathogenesis-related,PR) PR-14家族中[4]。在离体条件下,这类蛋白质对病原体也表现出很强的抗性[5-7]。植物幼嫩叶片的表面蜡质中包含大量的LTP蛋白,而在叶片表面发生的主要生物学事件是角质的沉积,所以很多研究认为LTP蛋白的功能是将分泌到细胞外的亲脂性物质如蜡质和角质单体转运到特定的位置[8]。
拟南芥AT4G22470基因编码的蛋白质具有2个8CM结构域,2个富含脯氨酸结构域,由于对该基因的功能尚未见报道,本研究将其命名为AtDHyPRP1 (DOUBLE HYBRID PROLINE-RICH PROTEIN 1)。AtDHyPRP1属于蛋白酶抑制剂/种子储藏蛋白/脂转移蛋白家族,氨基末端存在一段信号肽序列,在功能上可能与脂类分子的转运有关。GENEVESTIGATOR数据库中的基因芯片分析结果显示AtDHyPRP1基因在拟南芥原生质体和成熟叶片中表达水平很高,它可以被丁香假单胞菌诱导。本研究利用转基因拟南芥和烟草研究了AtDHyPRP1蛋白的亚细胞定位和它对灰霉菌的抗性。
1 材料与方法
1.1 材料
1.1.1 植物材料和细菌菌株
使用的起始材料为拟南芥 Arabidopsis thaliana Ws生态型 (Wassilewskija) 种子和秦烟Nicotiana tabacum 95种子。基因克隆实验的受体细胞为大肠杆菌Escherichia coli DH5α,拟南芥和烟草遗传转化实验采用农杆菌LBA4404。
1.1.2 主要试剂
限制性内切酶、T4 DNA连接酶和PCR Mix购自Fermentas公司和西安润德公司,PFU DNA聚合酶、克隆载体pMD18-T和植物双元表达载体pRI101-AN购自TaKaRa公司,植物双元表达载体pCAMBIA1302为本实验室保存,琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒购自OMEGA公司,地高辛杂交检测试剂盒为Roche公司产品。
1.2 方法
1.2.1 AtDHyPRP1过表达载体的构建
根据AtDHyPRP1基因的ORF设计一对引物,上下游引物分别为 P1、P2 (表 1)。由于AtDHyPRP1基因没有内含子,所以本研究以拟南芥 Ws野生型植株基因组 DNA为模板进行PCR反应。将扩增产物连接到pMD18-T载体中,转化大肠杆菌DH5α,经菌液PCR鉴定后送上海生工生物技术公司进行测序。用质粒小量提取试剂盒分离 pMD18-T-AtDHyPRP1,经 VspⅠ和SacⅠ双酶切后用 DNA回收试剂盒 (OMEGA)回收大小为1 128 bp的AtDHyPRP1目的片段。同时用SacⅠ和VspⅠ同尾酶NdeⅠ对植物双元表达载体pRI101-AN进行双酶切,回收载体大片段。接着用T4 DNA连接酶进行连接,构建产生pRI101-AN-AtDHyPRP1过表达载体,经菌液PCR及酶切鉴定后送上海生工生物技术公司进行测序。质粒DNA提取、酶切、连接、转化以及大肠杆菌感受态细胞的制备等参照《分子克隆实验指南》中的常规方法进行[9]。
1.2.2 AtDHyPRP1-GFP融合表达载体的构建
以测序正确的 pMD18-T-AtDHyPRP1作为模板,根据AtDHyPRP1基因的开放阅读框设计一对引物,上下游引物分别为P3、P4 (见表1)。采用PFU高保真DNA聚合酶进行PCR反应,用NcoⅠ和SpeⅠ对纯化的1 128 bp的扩增产物和pCAMBIA1302-GFP进行双酶切,用T4 DNA连接酶将 AtDHyPRP1编码序列插入到pCAMBIA1302的35S启动子和gfp报告基因之间,经菌液 PCR及酶切鉴定后送上海生工生物技术公司进行测序。
1.2.3 拟南芥的遗传转化与AtDHyPRP1蛋白的亚细胞定位
将pCAMBIA1302-AtDHyPRP1-GFP融合表达载体和含有 gfp报告基因的空载体pCAMBIA1302通过冻融法分别转化到农杆菌菌株LBA4404中,经菌落PCR验证后参照Bechtold和 Clough建立的方法转化拟南芥[10-11]。按单株收集转化当代的成熟种子,在含25 mg/L潮霉素的MS培养基上进行筛选。采用CTAB法从叶片组织提取基因组DNA[12],进行PCR鉴定。潮霉素抗性植株分析使用的上下游引物分别为 P5、P6 (表1)。将T1代AtDHyPRP1-GFP和GFP转基因拟南芥种子消毒后播种于附加25 mg/L潮霉素的 MS培养基中,10 d后将根切下,在OLYMPUS激光扫描共聚焦显微镜下进行观察。使用495~511 nm FITC滤光器拍摄GFP荧光,激发光波长为488 nm。
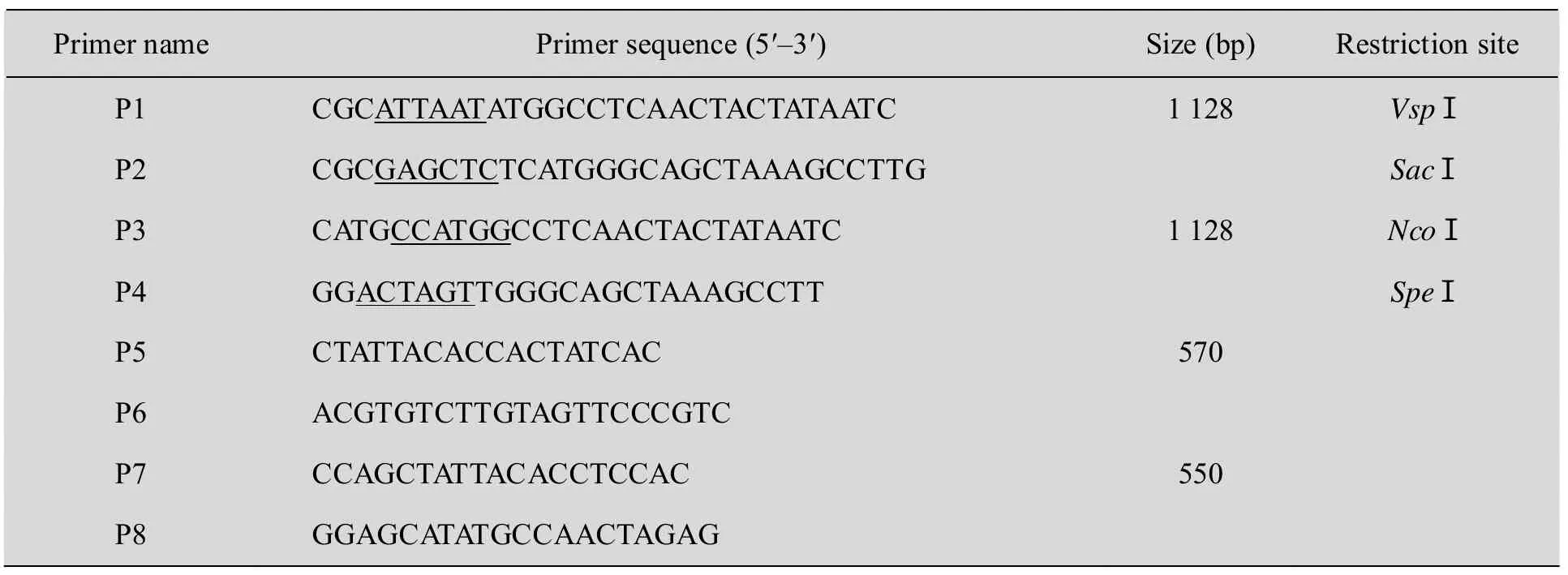
表1 PCR扩增所用引物序列Table 1 Primer sequences for PCR amplification
1.2.4 转AtDHyPRP1烟草植株的再生和分子鉴定
采用液氮冻融法将pRI101-AN-AtDHyPRP1导入农杆菌 LBA4404感受态细胞,在附加100 µg/mL链霉素、50 µg/mL卡那霉素和20 µg/mL利福平的YEB培养基上进行选择。抗性菌落经 PCR鉴定后在附加相同抗生素的YEB液体培养基中培养至 OD600≈0.5,用于烟草转化。将烟草无菌苗叶片切成块状,放入农杆菌悬浮液侵染5~10 min。在含有3%蔗糖、0.65%琼脂的MS固体培养基上暗培养3 d后,将外植体转移到附加 0.5 mg/L IAA、3 mg/L 6-BA、50 mg/L卡那霉素 (Kan)、500 mg/L头孢霉素 (Cef)、3%蔗糖和0.65%琼脂的分化培养基上进行选择。将分化出的不定芽从基部切下,转移到附加50 mg/L卡那霉素 (Kan)、500 mg/L头孢霉素 (Cef)、3%蔗糖和0.65%琼脂的MS培养基上生根。
采用CTAB法提取基因组DNA,对具有卡那霉素抗性的烟草植株进行PCR检测。PCR反应所用上下游引物分别为P7、P8 (表1)。为了检测AtDHyPRP1基因在烟草基因组中的拷贝数,用SacⅠ对20 μg基因组DNA进行单酶切,用PCR扩增产生的AtDHyPRP1基因ORF中间的一段550 bp的序列制备探针,按Roche公司的DIG High Prime DNA Labeling and Detection Starter Kit Ⅱ试剂盒使用手册进行Southern印迹分析。同时用Trizol试剂 (Invitrogen) 提取总RNA,通过RT-PCR和Northern印迹技术进行表达分析。在RT-PCR试验中,用RT-Master Mix反转录试剂盒 (TaKaRa) 合成cDNA,PCR反应所用上下游引物分别为 P7、P8 (表 1),扩增产物大小550 bp。在Northern印迹实验中,采用甲醛变性胶对提取的RNA进行分离,杂交和检测方法参照DIG High Prime DNA Labeling and Detection Starter Kit Ⅱ (Roche) 说明书进行。
1.2.5 蒜薹灰霉菌侵染分析
蒜薹灰霉菌Botrytis cinerea Pers.ex of Garlic Sprout由本实验室保存。菌株在马铃薯培养基上于28 ℃培养4 d后,用无菌水悬浮孢子,经玻璃棉过滤除去菌丝后用 1/4马铃薯培养基稀释成5×104个孢子/mL的悬浮液。将10 μL孢子悬浮液接种于野生型秦烟95和AtDHyPRP1转基因烟草叶片表面,然后置于22 ℃、16 h光照、90%湿度条件下,每隔5 d观察叶片的颜色变化情况和灰霉菌的侵染程度。侵染实验重复3次,每次接种3片叶子。
为了检测侵染部位H2O2的积累水平,将侵染5 d后的叶片用0.5% DAB (3,3-二氨基联苯胺) 在37 ℃染色过夜,经乙醇脱色后用80%甘油封片,在显微镜下观察侵染情况。同时,在接种5 d后用0.4%台酚蓝于37 ℃染色1 h,在85%乙醇中煮10 min脱去叶绿素,用80%甘油封片,在显微镜下观察细胞的存活状态。
2 结果与分析
2.1 AtDHyPRP1蛋白的结构特征
AtDHyPRP1基因 (GenBank Accession No. NM_118373) 开放阅读框全长为 1 128个碱基对,无内含子,编码的蛋白质由375个氨基酸构成 (图1)。AtDHyPRP蛋白的氨基酸序列包括4个部分,从氨基末端开始依次为信号肽 (1-23)、富含脯氨酸结构域 PRD (Proline-rich domain) (24-173)、8CM (Eight Cysteine Motif) (174-254)、PRD (255-294) 和8CM (295-375)。2个8CM只有 2个氨基酸不同,分别为 I209/V330和P228/R349。
2.2 AtDHyPRP1基因的分离和载体构建
以拟南芥Ws野生型植株基因组DNA为模板,通过PCR扩增AtDHyPRP1基因的ORF序列,1%琼脂糖凝胶电泳结果显示有两条带,一条符合预期大小 (1 128 bp),一条位于750~1 000 bp之间。出现这种结果的原因是AtDHyPRP1蛋白有2个8CM,PCR反应所用的下游引物可以与这2个8CM编码序列的3'端配对。将大片段回收后连接至pMD18-T载体,测序结果表明有2个碱基与数据库中下载的来自 Col-0的AtDHyPRP1基因序列不同,分别为 765位的T变成了A,811位的A变成了C。Ws生态型中的第1个碱基变化没有引起氨基酸改变,但第2个碱基变化使第271位的K变为Q。

图1 AtDHyPRP1蛋白 (NP_193978.2) 的氨基酸序列Fig. 1 The deduced amino acid sequence of AtDHyPRP1. The signal peptide is underlined. Two 8CM are framed, the conservative cysteine residues are shadowed, the different amino acid residues between two 8CM are shown in bold.
用VspⅠ和SacⅠ对pMD18-T-AtDHyPRP1进行双酶切,得到 AtDHyPRP1编码序列。用NdeⅠ和 SacⅠ对植物双元表达载体 pRI101-AN进行双酶切,得到线性化质粒 DN A。将AtDHyPRP1编码序列与pRI101-AN质粒DNA连接,构建产生 pRI101-AN-AtDHyPRP1。将pRI101-AN-AtDHyPRP1送上海生工生物技术公司测序,结果没有变化,表明载体构建成功。
以测序正确的 pMD18-T-AtDHyPRP1为模板,采用高保真 PFU DNA聚合酶扩增AtDHyPRP1编码序列,上下游引物 5¢端分别包含NcoⅠ和SpeⅠ酶切位点。用NcoⅠ和SpeⅠ对扩增产物和植物双元表达载体pCAMBIA1302进行双酶切,用T4 DNA连接酶将AtDHyPRP1编码序列连接到pCAMBIA1302中的gfp基因之前,产生pCAMBIA1302-AtDHyPRP1-GFP融合表达载体。将pCAMBIA1302-AtDHyPRP1-GFP送上海生工生物技术公司测序,结果显示序列正确,读码框没有变化。
2.3 转基因拟南芥的分子鉴定和AtDHyPRP1蛋白的亚细胞定位
用包含pCAMBIA1302-AtDHyPRP1-GFP的农杆菌LBA4404通过真空渗透方法转化拟南芥Ws野生型植株,在附加25 mg/L潮霉素的MS培养基上对收获的种子进行筛选。未转化的种子苗在潮霉素抗性培养基上不能正常生长,逐渐黄化死去,而转基因植株能够正常生长。用激光共聚焦显微镜观察 PCR检测阳性的转基因拟南芥植株的根,结果显示AtDHyPRP1定位于细胞表面,表明AtDHyPRP1蛋白主要分布在细胞膜或细胞壁上 (图2A)。而pCAMBIA1302空载体转化的拟南芥根细胞中 GFP分布在整个细胞内(图2B)。
2.4 转AtDHyPRP1基因烟草的分子鉴定和表达分析
通过冻融法将重组质粒 pRI101-ANAtDHyPRP1转入农杆菌 LBA4404,采用菌落PCR进行鉴定。用含有pRI101-AN-AtDHyPRP1的农杆菌LBA4404对秦烟95进行遗传转化,经卡那霉素筛选和PCR检测获得6个转基因植株。图3为TT1和TT2两个转基因烟草株系的PCR检测结果。
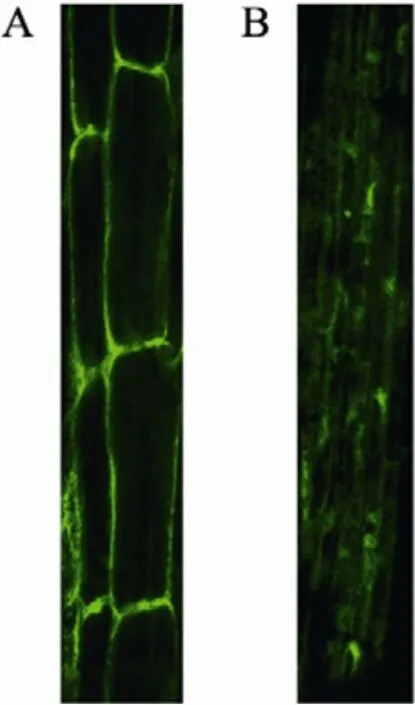
图2 转基因拟南芥根细胞中GFP荧光的观察Fig. 2 Fluorescence detection of GFP in root cells of transgenic Arabidopsis. (A) GFP fluorescence in root cells of pCAMBIA1302-AtDHyPRP1-GFP transformed Arabidopsis plant. (B) GFP fluorescence in root cells of pCAMBIA1302 transformed plant.
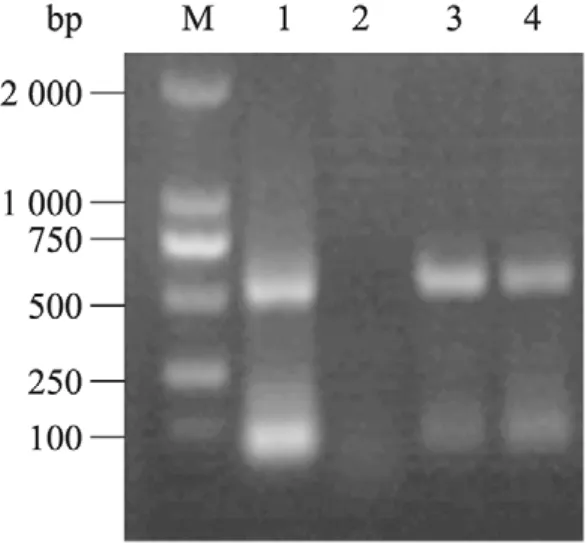
图3 转AtDHyPRP1基因烟草的PCR检测Fig. 3 PCR analysis of the transgenic tobacco plants harboring AtDHyPRP1. M: DL2000 DNA marker; 1: pRI101-AN-AtDHyPRP1; 2: wild type tobacco plant; 3–4: transgenic tobacco plants with kanamycin resistance (TT1 and TT2, respectively).
从PCR鉴定为阳性的转AtDHyPRP1基因烟草提取基因组 DNA,经限制性酶切后进行Southern印迹分析,结果显示TT1整合有一个拷贝的AtDHyPRP1基因 (图4-3),TT2整合有两个拷贝的AtDHyPRP1基因 (图4-4)。对这2个转基因植株进行RT-PCR (图5) 和Northern印迹(图6) 分析,结果表明AtDHyPRP1基因能够有效表达。
2.5 转AtDHyPRP1基因烟草对蒜薹灰霉菌的抗性分析

图4 T0代转AtDHyPRP1基因烟草的Southern印迹分析Fig. 4 Southern blotting analysis of T0 tobacco plants harboring AtDHyPRP1. 1: pRI101-AN-AtDHyPRP1 digested with Sac I and Xba I; 2: genomic DNA of wild type tobacco plant digested with Sac I; 3–4: genomic DNA of transgenic tobacco plants harboring AtDHyPRP1 digested with Sac I (TT1 and TT2, respectively).
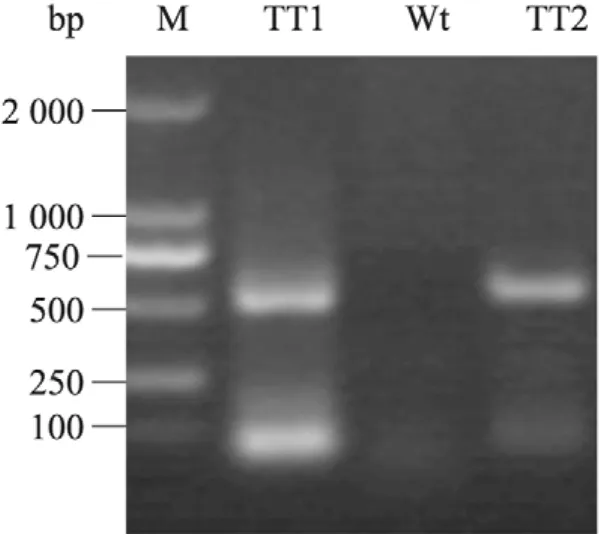
图5 转AtDHyPRP1基因烟草的RT-PCR检测Fig. 5 RT-PCR analysis of the transgenic tobacco plants harboring AtDHyPRP1. M: DL2000 DNA marker; TT1: transgenic tobacco line TT1; Wt: wild type tobacco plant; TT2: transgenic tobacco line TT2.

图6 转AtDHyPRP1基因烟草的Northern blotting分析Fig. 6 Northern blotting analysis of the transgenic tobacco plants harboring AtDHyPRP1. Wt: wild type tobacco plant; TT1,TT2: transgenic tobacco plant TT1 and TT2, respectively.
在3次重复实验中,共接种9个TT1叶片,9个TT2叶片,9个野生型叶片。蒜薹灰霉菌侵染5 d后,转AtDHyPRP1基因烟草仍然保持绿色,而野生型叶片四周已经开始褪绿。侵染8 d后,野生型烟草叶片上的病斑逐渐扩大,超过50%的叶片表面颜色变黄;而TT1有7个叶片仍为绿色,2个叶片在叶边缘出现变黄现象;TT2有6个叶片仍为绿色,3个叶片在叶边缘出现变黄现象。说明野生型烟草相对于转 AtDHyPRP1烟草在受到灰霉菌侵染时叶片褪绿的时间要早。侵染 12 d后,野生型烟草叶片变黄面积超过90%,其中3个叶片已经腐烂;TT1只有1个叶片基本上保持绿色,8个叶片的边缘已经褪绿,其中2个叶片的变黄部分已由边缘向中间扩散,其余6片只是在叶边缘出现变黄现象,叶片变黄面积约为20%;TT2有3个叶片的变黄部位已由边缘向中间扩散,6个叶片只是在叶边缘出现变黄现象,叶片变黄面积约为30%,并且在大部分侵染位点能看到有黄色的斑点,但斑点与健康组织之间存在明显的界限,无叶片腐烂现象出现。侵染20 d后,9个TT1叶片都已褪绿,有1片叶边缘出现黄色,其余叶片进一步变黄,变黄面积达到50%;TT2叶片变黄面积达到60%;这时9个野生型烟草叶片几乎完全枯死,3片软化成为湿腐状,最终遍及整片叶子,而 18个转AtDHyPRP1基因烟草叶片都没有腐烂,叶片中部仍然保持有绿色 (图7)。以上结果说明与野生型烟草相比,转AtDHyPRP1基因烟草对灰霉菌具有明显的抗性。
具有抗性的植物在受到病原体侵染时,会发生超敏反应 (Hypersensitive response,HR)。HR指病原体入侵后的很短时间内被侵染位点产生的大量活性氧 (ROS) 能够引起细胞死亡,将病原体的生长限制在一定的范围内,防止其扩散并造成大面积的伤害。ROS包括H2O2、O2−和OH等。DAB可以和H2O2反应而显示出红褐色[13]。在蒜薹灰霉菌侵染10 d后,野生型烟草叶片经过DAB染色可以观察到有少量的染色较浅的区域,显示灰霉菌侵染引起了较低水平的 H2O2积累,没有产生超敏反应,病原体可以扩散,菌丝的生长最终引起整个叶片枯萎 (图 8A)。而转AtDHyPRP1基因烟草叶片经DAB染色后可以观察到较为集中的染色较深的红褐色区域,显示高水平H2O2积累引发了HR反应,灰霉菌不能继续侵染周围的细胞 (图 8B),说明 AtDHyPRP1基因可以明显提高烟草的抗真菌能力。台酚蓝染色结果显示野生型烟草叶片在灰霉菌侵染之后出现大量死亡细胞 (图 9A),而转AtDHyPRP1基因烟草叶片细胞的坏死现象明显减轻 (图9B)。

图7 转AtDHyPRP1基因烟草对蒜薹灰霉菌的抗性分析Fig. 7 Resistance of the transgenic tobacco plants to B. cinerea Pers. ex of Garlic Sprout. (A) 5 d after inoculation with the conidia of B. cinerea. (B) 8 d after inoculation with the conidia of B. cinerea. (C) 12 d after inoculation with the conidia of B. cinerea. (D) 20 d after inoculation with the conidia of B. cinerea. TT1 and TT2: transgenic tobacco plant TT1 and TT2, respectively; Wt: wild type tobacco plant.

图8 H2O2积累水平的DAB染色观察Fig. 8 Observation of H2O2 accumulation by DAB staining. (A) Leaf of wild type tobacco plant. (B) Leaf of transgenic tobacco plant harboring AtDHyPRP1.
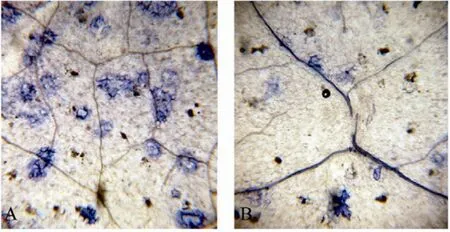
图9 侵染烟草叶片的台酚蓝染色Fig. 9 Trypan blue staining of the tobacco leaves after infection with B. cinerea. (A) Leaf of wild type tobacco plant. (B) Leaf of transgenic tobacco plant harboring AtDHyPRP1.
3 讨论
本研究从拟南芥中克隆了 AtDHyPRP1基因,生物信息学分析发现AtDHyPRP1在N端具有信号肽序列,显示该蛋白在合成后可能被运送或分泌到植物细胞的表面来执行特殊的功能。对GENEVESTIGATOR (www.genevestigator.ethz. ch/gv/index.jsp) 数据库中的芯片实验结果进行分析,显示AtDHyPRP1基因在受到丁香假单胞菌侵染诱导时表达水平最高,说明该基因与植物的抗病防御机制有关。本研究通过农杆菌转化方法将AtDHyPRP1基因导入烟草基因组,蒜薹灰霉菌侵染结果证实组成性表达该基因对病原体确实具有明显的抗性。
AtDHyPRP1蛋白的相对分子质量为38.79 Da,具有2个8CM结构域,2个PRD结构域,是一种新型的非特异性脂转移蛋白。尽管对AtDHyPRP1的功能目前还没有研究报道,但是从大规模检测拟南芥个体经不同处理后的基因表达水平的芯片实验数据中,可以找到一些有关该基因的信息。由于包含富脯氨酸序列,AtDHyPRP1也被认为是一种PRP/Hybrid PRP蛋白[14]或类Extensin蛋白 (Extensin-like)[15],它可能与细胞周期调控有关,在G1期存在表达峰值。用无毒丁香假单胞菌Pst DC3000 (avrRpt2) 处理gh3.5-1D激活标签突变体后,GH3.5在侵染部位和其他组织中都会过量表达,SA积累水平和PR-1表达水平均表现增加,在这种系统性获得抗性应答过程中AtDHyPRP1基因的表达水平会上升3.3倍[16]。用Pst hrpA、Pst COR-hrpS、Pst DC3000和大肠杆菌等病原体处理拟南芥时AtDHyPRP1基因的转录水平均表现上升[17]。 Mohr和Cahill的微阵列实验结果显示Pst无毒菌株侵染可以使AtDHyPRP1表达水平明显提高,同时发现叶片中ABA浓度的增加能够使拟南芥对Pst无毒菌株变得十分敏感,并伴随着木质素和水杨酸浓度的下降,以及抗性相关基因表达的抑制[18]。以上研究报道说明AtDHyPRP1基因编码的蛋白质主要参与拟南芥对病原体的抗性应答过程。
在与病原体的相互作用中,LTP可能参与激发子触发植物细胞过敏性反应的信号转导过程。激发子是一类能诱导寄主植物产生防卫反应的特殊化合物,与受体分子结合后可通过一定的信号传导途径激发植物产生防卫反应[19]。另外,激发子具有甾醇转移活性,它们与甾醇形成的复合物可以和植物细胞膜上的受体蛋白互相作用,引起植物细胞的超敏反应,从而激活抗病防卫反应的信号转导途径,诱导植物抗性。发生HR反应后,侵染位点会有H2O2等活性氧 (ROS) 分子的积累。由于 LTP和激发子在空间结构上有相似性,所以LTP可以竞争性结合激发子的膜受体,抑制细胞死亡并诱导系统抗性的产生[20]。本研究用蒜薹灰霉菌孢子对转AtDHyPRP1基因烟草和野生型烟草叶片进行离体侵染,从DAB染色结果可以明显地看出野生型烟草叶片表面的被侵染位点着色较浅,病原体可以扩散。而转AtDHyPRP1基因烟草叶片上的侵染位点着色很深,说明发生了HR反应并有大量的H2O2积累,从而限制了菌体的扩散。台酚兰染色结果也证实灰霉菌侵染可以引起野生型叶片细胞的大量死亡,而转AtDHyPRP1基因烟草叶片细胞的坏死数量明显减少。
蛋白质是生物功能的具体执行者,蛋白质的功能与其在细胞中的亚细胞定位密切相关。本研究在构建pCAMBIA1302-AtDHyPRP1-GFP植物表达载体的基础上,通过浸花法获得了表达AtDHyPRP1-GFP融合蛋白的拟南芥转基因植株,激光共聚焦显微观察显示AtDHyPRP1定位于细胞表面,这与生物信息学分析和灰霉菌抗性分析结果一致。
[1] Arondel V, Vergnolle C, Cantrel C, et al. Lipid transfer proteins are encoded by a small multigene family in Arabidopsis thaliana. Plant Sci, 2000, 157(1): 1−12.
[2] Suzuki H, Xia YJ, Cameron R, et al. Signals for local and systemic responses of plants to pathogen attack. J Exp Bot, 2004, 55(395): 169−179.
[3] Maldonado AM, Doerner P, Dixon RA, et al. A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature, 2002, 419(6905): 399−403.
[4] Van Loon LC, Van Strien EA. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol, 1999, 55(2): 85−97.
[5] Nishimura S, Tatano S, Gomi K, et al. Chloroplast-localized nonspecific lipid transfer protein with anti-fungal activity from rough lemon. Physiol Mol Plant P, 2008, 72(4/6): 134−140.
[6] Molina A, García-Olmedo F. Enhanced tolerance to bacterial pathogens caused by the transgenic expression of barley lipid transfer protein LTP2. Plant J, 1997, 12(3): 669−675.
[7] Ge XC, Chen JC, Li N, et al. Resistance function of rice lipid transfer protein LTP110. J Biochem Mol Biol, 2003, 36(6): 603−607.
[8] Zottich U, Da Cunha M, Carvalho AO, et al. Purification, biochemical characterization and antifungal activity of a new lipid transfer protein (LTP) from Coffea canephora seeds with α-amylase inhibitor properties. Biochim Biophys Acta, 2011, 1810(4): 375−383.
[9] Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual. 3rd ed. New York: Cold Spring Harbor Laboratory Press, 2001.
[10] Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci, 1993, 316: 1194−1199.
[11] Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J, 1998, 16(6): 735−743.
[12] Stewart CN Jr, Via LE. A rapid CTAB DNA isolation technique useful for RAPD fingerprinting and other PCR applications. Biotechniques, 1993, 14(5): 748−749.
[13] Wang CF, Huang LL, Buchenauer H, et al. Histochemical studies on the accumulation of reactive oxygen species (O2-and H2O2) in the incompatible and compatible interaction of wheat——Puccinia striiformis f. sp. tritici. Physiol Mol Plant Pathol, 2007, 71(4/6): 230−239.
[14] Schultz CJ, Rumsewicz MP, Johnson KL, et al. Using genomic resources to guide research directions. The Arabinogalactan protein gene family as a test case. Plant Physiol, 2002, 129(4): 1448−1463.
[15] Menges M, Hennig L, Gruissem W, et al. Cell cycle-regulated gene expression in Arabidopsis. J Biol Chem, 2002, 277(44): 41987−42002.
[16] Zhang ZQ, Li Q, Li ZM, et al. Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-Pseudomonas syringae interaction. Plant Physiol, 2007, 145(2): 450−464.
[17] Thilmony R, Underwood W, He SY. Genome-wide transcriptional analysis of the Arabidopsis thaliana interaction with the plant pathogen Pseudomonas syringae pv. tomato DC3000 and the human pathogen Escherichia coli O157:H7. Plant J, 2006, 46(1): 34−53.
[18] Mohr PG, Cahill DM. Suppression by ABA of salicylic acid and lignin accumulation and the expression of multiple genes, in Arabidopsis infected with Pseudomonas syringae pv. tomato. Funct Integr Genomics, 2007, 7(3): 181−191.
[19] Liu MF, Zhu XC, Xiao HS. Research progress of elicitor and it’s signal transduction in plants. Chin Agric Sci Bull, 2008, 24(3): 215−220.刘梅芳, 朱晓程, 肖华山. 植物激发子及其信号传导之研究进展, 中国农学通报, 2008, 24(3): 215−220.
[20] Blein JP, Coutos-Thévenot P, Marion D, et al. From elicitins to lipid-transfer proteins: a new insight in cell signalling involved in plant defence mechanisms. Trends Plant Sci, 2002, 7(7): 293−296.