甘薯块根碳水化合物合成与积累动态特性研究
2011-11-17吕长文王季春唐道彬王三根李艳花
吕长文 王季春 唐道彬 王三根 赵 勇 李艳花
(西南大学农学与生物科技学院1,重庆 400716)
(重庆市甘薯工程技术研究中心2,重庆 400716)
甘薯块根碳水化合物合成与积累动态特性研究
吕长文1,2王季春1,2唐道彬1,2王三根1赵 勇1,2李艳花1
(西南大学农学与生物科技学院1,重庆 400716)
(重庆市甘薯工程技术研究中心2,重庆 400716)
以不同干物质类型的甘薯品种绵粉 1号、南薯 88和商丘 52-7为材料,研究了甘薯块根形成与膨大期间碳水化合物积累与淀粉合成相关酶活性的动态变化及其相互关系。结果表明,作为品种的固有特征,干物质含量或淀粉含量的差异主要决定于品种的遗传特性。对于中高干率品种而言,淀粉在生育中后期积累较快。磷酸蔗糖合成酶(SPS)活性与品种间淀粉积累量一致,中等干物质品种 (南薯 88)中后期的蔗糖合成酶 (SS)活性较高,淀粉积累量也最多,SPS和 SS对淀粉合成与积累具有促进作用,ADPG焦磷酸化酶有随气温降低而活性下降的趋势。此外不同品种均表现出甘薯块根干物质含量与可溶性糖含量、淀粉产量与可溶性糖含量呈极显著的负相关关系。
甘薯 碳水化合物 合成与积累 动态变化
甘薯 [Ipom oea batatas(L.)Lam.]在我国常年种植面积约 6 000 000 hm2,在世界粮食生产中总产量列第 7位,它既是一种宜粮、宜饲、宜加工的多用途粮食作物[1],同时还具有多样化的食品保健功能[2-3]。近年来,随着世界能源的紧缺,新型替代可再生能源的开发和利用又将甘薯的作用继续延伸,在发展新型清洁能源方面发挥了极为重要的作用。然而甘薯遗传资源丰富,品种类型多样,传统育种方式难以实现定向选育,能否作为生物质能源作物则完全取决于品种的干率及淀粉产量的高低,而甘薯淀粉和其他碳水化合物的积累又与控制淀粉合成途径的酶极为相关。在对相关酶的研究上,有关小麦、水稻、玉米等主要粮食作物的研究报道较多[4-6],而对不同干物质含量类型甘薯的相关研究较少,对指导甘薯的定向育种的理论研究极为缺乏。本试验选择了 3种干物质含量差异较大 (淀粉含量)特异型品种,对其淀粉合成相关酶活性变化与碳水化合物形成积累的相关特性进行了研究,对阐明不同类型品种淀粉酶特性与淀粉合成与积累的关系,进而利用这种相互关系实现甘薯专用型品种的定向选育,具有一定的理论与应用价值。
1 材料和方法
1.1 试验材料
试验材料于 2006~2008年种植于重庆西南大学作物科研基地,包括高干率型品种绵粉 1号 (MF)、兼用型品种南薯 88(NS)和低干率型品种商丘 52-7(S Q),均由西南大学薯类作物研究所提供。密度为亩植 3 000株,从 50天开始取样,每 15天一次共计 5次取样。
1.2 块根干物质含量测定
取大小基本一致薯块去皮后,切成粒状称量50 g,于 105℃下杀青 15 min后,于 80℃下烘至恒重。
1.3 块根碳水化合物含量测定
将烘至恒重的薯粒磨细,100目过筛后,取样0.1 g进行淀粉、可溶性糖测定。淀粉含量采用高氯酸 -I2-KI测定[7],可溶性糖采用蒽酮法测定[8]。
1.4 块根淀粉合成相关酶测定
取大小一致鲜薯块根,去皮后取 0.1 g样品,加入提取液 5 mL(含 100 mmol/L Tricine-NaOH,pH 7.5;8 mmol/L MgCl2;2 mmol/L EDTA;12.5%Glycerol;1%PVP-40,50 mmol/L 2-Mercap-toeth2 ano1),冰浴碾磨。碾磨后,离心 10 min(2℃,15 000 r/min),其上清液即为可溶性粗酶液[9-10]。该粗酶液用于蔗糖合成酶 (SS)、蔗糖磷酸合成酶 (SPS)和ADPG焦磷酸化酶 (AGPP)。
1.4.1 ADPG焦磷酸化酶 (AGPP)活性的测定
取 20μL酶粗提液加入 110μL反应液 (反应液最终浓度是 100 mmol/L Hepes-NaOH(pH 7.4)、1.2 mmol/L ADPG,3 mmol/L PPi;5mmol/L MgCl2,4 mmol/L DTT)。30℃反应 20 min后,沸水中终止反应 30 s。10 000×g离心 10min后取上清液 100μL,加 5.2μL比色液 (5.76 mmol/L NADP,0.08 unit PG M,0.07 unit G6PDH)。30℃反应 10 min后,测定340 nm OD值。
1.4.2 蔗糖磷酸合成酶 (SPS)活性测定
取 50μL粗酶液加 50μL HEPES-NaOH缓冲液、20μL 50 mmol/L MgCl2、20μL 100 mmol/L UD2 PG、20μL 100 mmol/L 6-磷酸果糖 ,反应 30 min后 ,加 200μL 1 mol/L NaOH终止反应,加 1.5 mL浓盐酸和 0.5 mL 1%间苯二酚,测定生成蔗糖的含量。
1.4.3 蔗糖合成酶 (SS)活性测定
取 50μL粗酶液加 50μL HEPES-NaOH缓冲液、20μL 50 mmol/L MgCl2、20μL 100 mmol/L UD2 PG、20μL 100 mmol/L 果糖 ,反应 30 min后 ,加 200 μL 1 mol/L NaOH终止反应,加 1.5 mL浓盐酸和 0.5 mL 1%间苯二酚,测定生成蔗糖的含量。
1.5 数据统计与分析
试验数据运用 Excel 2003和 SPSS13.0进行统计分析与作图。
2 结果与讨论
2.1 不同时期干物质形成差异
块根是甘薯干物质贮藏的主要器官,通过对不同类型甘薯块根干物质含量在不同时期的变化可以看出 (图 1),低干率品种商丘 52-7无论是在栽后50天块根形成的初期,还是块根形成并膨大 2个月后,鲜薯的干物质含量始终维持在 12%左右,而对于南薯 88和绵粉 1号,直至栽后 110天,干物质含量仍在进一步增加,但以块根膨大的前期增长较快。由于甘薯品种的干物质主要由其遗传特性决定,但在生育的不同时期,干物质含量仍有一定变化,商丘52-7前后期差异仅 1%左右,而高干率品种绵粉 1号的前后期干率差异达 10.65%,可见干物质含量越高的品种,前后期的干物质差异更为大。

图 1 不同品种块根干物质含量动态变化
2.2 不同类型甘薯碳水化合物形成差异比较
从图 2可知,甘薯淀粉的形成和积累也需要一定时期。在块根形成后,薯块中淀粉增量并非随着块根的膨大而同比例增加,其含量在薯块膨大中后期也才相对稳定,且淀粉在块根中的积累在前期非常迅速,在块根形成初期 (栽后 50天),即使低干率品种的薯干淀粉含量也达到了 35%以上。从各品种不同时期淀粉含量看,高干率品种的淀粉含量仍明显高于低干率品种。
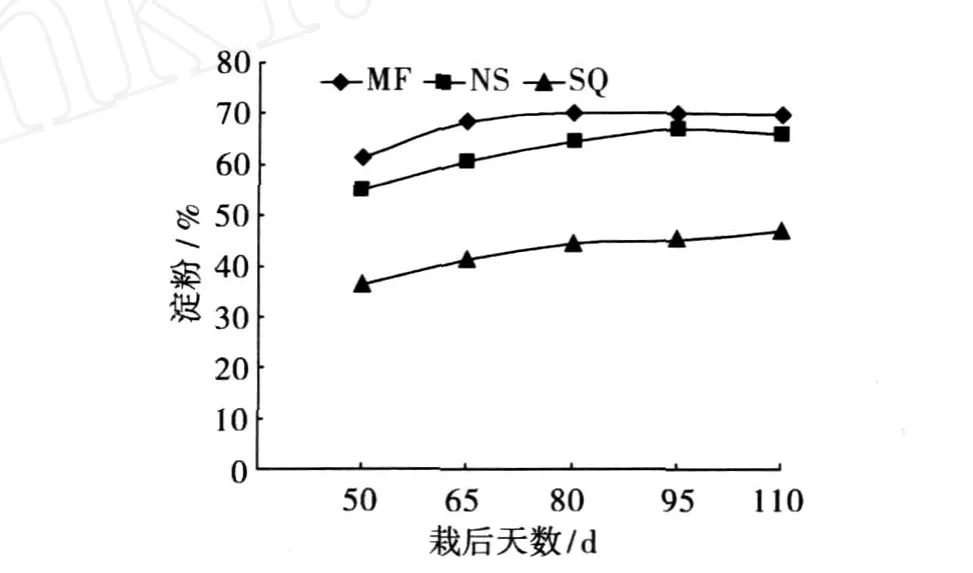
图 2 不同品种薯干淀粉含量动态变化
一般来说,甘薯淀粉含量与产量总是一对难以调和的矛盾性状,高淀粉低产量,低淀粉高产量。但从图 3所示结果看,栽后 80天之前,各品种的淀粉产量差异并不显著,但之后南薯 88与绵粉 1号与低干率品种商丘 52-7差距拉大,结合 3者的淀粉含量(图 2),可知低干率品种商丘 52-7前期块根膨大快,淀粉合成积累快,时间短,而后期积累少产量增幅较小,中干率品种南薯 88则在栽后 80天起块根迅速膨大,淀粉积累持续增长,产量大幅提升,绵粉 1号在整个时期则保持了较为稳定的淀粉合成与积累速度,从而最终各品种在淀粉产量的差异上呈现出低干率低产、中干率高产、高干率中产的结果。从不同干物质类型品种的淀粉积累与合成动态变化看,中干率品种的淀粉合成与积累时期较长,而高干率品种次之,低干率品种淀粉积累期则相对较短。从前人研究结果可知,干物质含量 (或淀粉)与产量呈显著负相关关系,这也是难以育成具有突破性的高干率且高产品种的根本原因。因此,在选育不同类专用型品种方面,高产应为首选指标,即使鲜薯淀粉含量很高,产量不行也难以为生产所用,而对于干物质较低的品种,如果在产量方面有较大突破,其总的干物质含量仍会比较可观。
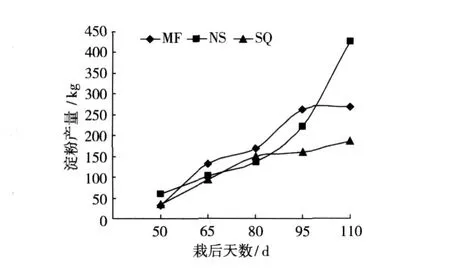
图 3 不同品种块根淀粉积累动态变化
通过对品种可溶性糖含量研究结果表明,该性状与其干物质或淀粉含量相反,干物质或淀粉含量越高的品种,可溶性糖含量就越低 (图 4)。在同一时期,块根可溶性糖含量均表现为商丘 52-7高于南薯 88,后者又高于绵粉 1号。即可溶性糖与干物质或淀粉含量呈负相关关系,前者可能是淀粉合成的供体物质。根据淀粉是在叶绿体或淀粉体中通过一系列酶反应合成这一前提,Muller等[11]提出淀粉生物合成的两条途径,无论是在叶绿体中还是库细胞中,葡萄糖、果糖、蔗糖均为淀粉合成的主要原料或前体物质,包劲松[12]所提到的淀粉合成途径模型也证实了这点,蔗糖作为可溶性糖的一种,在淀粉类作物中的淀粉形成与积累具有决定性的作用。沈淞海等[13]在研究甘薯生长发育过程中可溶性糖含量与淀粉积累的关系时发现,非还原糖在甘薯块根中占总糖含量的 51%~88%,而蔗糖占非还原糖的 90%以上,淀粉积累率低时,块根总糖水平急剧升高,同时认为蔗糖含量与块根淀粉积累率呈负相关。
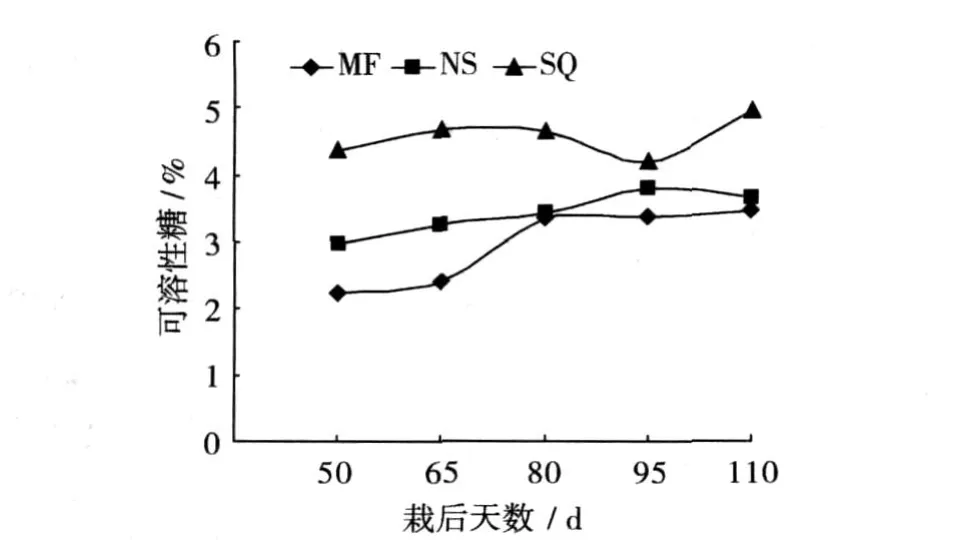
图 4 不同品种块根薯干可溶性糖含量动态变化
2.3 淀粉合成酶类活性比较
从图 5可知,在块根形成与膨大初期,各品种的蔗糖磷酸合成酶均处于较高活性,尤以高淀粉品种绵粉 1号为最,随后逐渐降低,至栽后 80天又逐渐升高,最后 3品种该酶活性基本处于同一水平。从整个薯块膨大期来看,以南薯 88的蔗糖磷酸合成酶活性相对较高,绵粉 1号次之,商丘 52-7为最低,可见该酶整体活性与各品种的淀粉合成与积累表现一致。
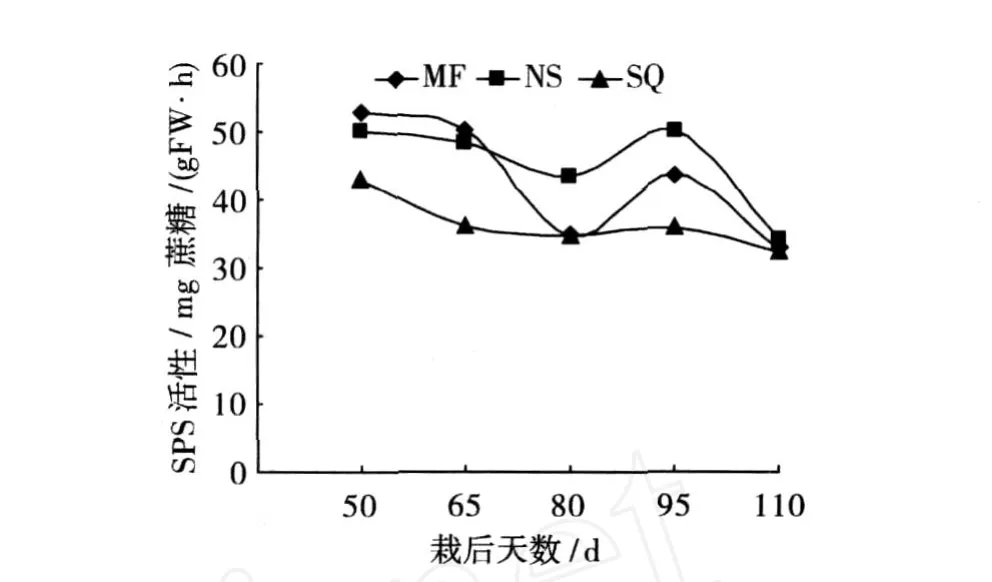
图 5 不同品种块根蔗糖磷酸合成酶活性差异
而块根蔗糖合成酶在薯块形成与膨大期间呈现出两个拐点的波浪式变化 (图 6)。首先在栽后 65天,3品种均出现了一个波峰 (较大值),而后呈不同程度的下降,特别是高干率品种蔗糖合成酶迅速下降,在栽后 80天出现活性又反弹上升,而其他两个品种则在栽后 95天又才由最低值逐渐上升。从块根的整个膨大期来看,该酶活性均维持在 20~60酶活力单位。同时,ADPG焦磷酸化酶是各类淀粉质类作物淀粉形成的关键性调节酶。由图 7可知,该酶在甘薯形成与膨大初期最高,后期相对较低,在整个生育期间变幅较大。在栽后70~80天时,3品种该酶活性均维持在 1 200~1 400酶活单位。随后该酶活性呈继续下降趋势。这也可能是由于后期气温下降所致,对酶活性产生了较大影响。

植物淀粉的生物合成主要涉及腺苷二磷酸葡萄糖焦磷酸化酶、淀粉合成酶、可溶性淀粉合成酶、淀粉分支酶和淀粉去分支酶,它们在淀粉的合成与积累过程中发挥着重要作用,是影响淀粉生物合成的关键酶[14-19]。Li等[20]研究发现,蔗糖合成酶在甘薯块根的形成与膨大中表达最活跃,而蔗糖合成酶途径在涉及淀粉积累的蔗糖分解过程中占主导。Yato2 mi等[21]通过对不同产量水平的两个甘薯品种淀粉合成相关酶活性比较发现,高产品种的蔗糖合成酶、AGPP以及淀粉合成酶的活性比低产品种相应酶的活性高,进而认为这些酶活性的高低是甘薯品种高产的原因之一。从本试验 AGPP活性看,整个生育期呈下降趋势,这与生育期环境温度变化较为一致,气温越高酶活性越强。在稳定温度条件下,该酶活性是否会有随着生育发育进程而发生类似变化趋势,还有待进一步研究。
2.4 相关性分析
通过进一步相关分析发现 (表 1),对所有品种而言,甘薯块根干物质含量、淀粉含量与可溶性糖含量之间均呈极显著的负相关关系,它们与可溶性糖含量的相关系数分别 -0.649和 -0.704;此外块根干物质与淀粉含量之间、SPS活性与可溶性糖含量之间则呈极显著正相关,其相关系数分别为 0.924和0.699。这也进一步证实了可溶性糖可能是甘薯淀粉合成的前体或中间物,同时 SPS活性越强,可溶性糖含量越低。
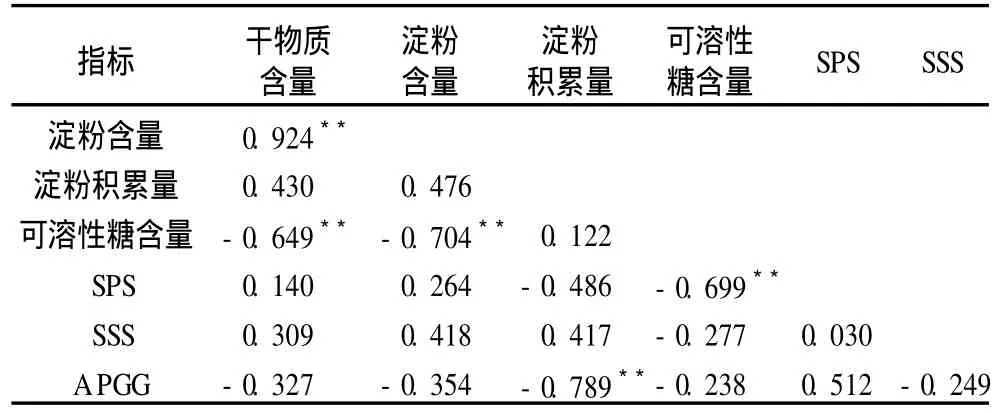
表 1 碳水化合物及相关酶活性相关性分析
3 结 论
3.1 品种干物质含量越高,生育前期与后期干物质含量差异越大。尽管南薯 88干物质含量一般,但其产量显著超过其他两个品种,其最终干物质积累量(或淀粉产量)仍高于绵粉 1号和商丘 52-7,这为品种淀粉用专用型品种选育提出了理论依据。即在品种选择中,宜选择中高干率 (物质)品系,这类品系淀粉合成与积累时间长,并非干物质 (淀粉含量)越高越好,才有可能选出具有突破性产量的优良品种。
3.2 薯干可溶性糖含量与干物质含量、淀粉含量之间呈极显著负相关,而块根干物质与淀粉含量之间、SPS活性与可溶性糖含量之间则表现为极显著正相关关系。可见可溶性糖可能是淀粉合成的前体物质或中间物,且高干率品种的可溶性糖转化合成淀粉的效率较高。
3.3 不同类型品种 SPS、SS及 APGG的酶活性变化趋势基本一致,但不同时期活性存在差异,低干率品种商丘 52-7的 SPS与 SS活性不同时期均较低,而全生育期内各品种平均 AGPP活性基本处于同一水平,由此也可见 SPS与 SS酶在甘薯块根淀粉合成与积累中具有积极作用,特别是对于高干率 (高淀粉)品种,上述酶活性更强。
[1]李强,刘庆昌,翟红,等.中国甘薯主要亲本遗传多样性的ISSR分析 [J].作物学报,2008,34(6):972-977
[2]台建祥,华希新,王家万,等.特白一号薯叶制品功能性实验及临床应用研究[J].作物学报,1998,24(2):161-167
[3]安康,房伯平,陈景益,等.甘薯保健功能的研究进展及发展前景[J].广东农业科学,2004,增刊:6-9
[4]高松洁,郭天财,王文静,等.不同穗型冬小麦品种灌浆期籽粒中与淀粉合成有关的酶活性变化[J].中国农业科学,2003,36(11):1373-1377
[5]蔡一霞,王维,张祖建,等.结实期水分亏缺对反义 Wx基因转化系水稻籽粒淀粉合成关键酶活性及淀粉累积的影响[J].作物学报,2006,32(3):330-338
[6]张智猛,戴良香,胡昌浩,等.氮素对玉米淀粉累积及相关酶活性的影响[J].作物学报,2005,31(7):956-962
[7]门福义,刘梦芸.马铃薯栽培生理 [M].北京:中国农业出版社,1995,317-320
[8]高俊凤.植物生理学实验指导 [M].北京:高等教育出版社,2006,144-148
[9]Doehlert Douglas C,Lamert Robert J.Metabolic characteris2 tics associatedwith starch,protein and oil deposition in devel2 opingmaize kernels[J].Crop Science,199l,31:151-157
[10]程方民,蒋德安,吴平,等.早籼稻籽粒灌浆过程中淀粉合成酶的变化及温度效应特征[J].作物学报,2001,27(2):201-206
[11]Muller-Rober B,Kobmann J.Approaches to influence starch quantity and starch quality in transgenic plants[J].Plant cell and environment,1994,17:601
[12]包劲松.植物淀粉生物合成研究进展 [J].生命科学,1999,11(增刊 ):104-107
[13]沈淞海,沈海铭,吴建华.甘薯生长发育过程中可溶性糖含量与淀粉积累的关系[J].浙江农业大学学报,1994,20(4):400-404
[14]康国章,王永华,郭天财,等.植物淀粉合成的调控酶[J].遗传,2006,28(1):110-116
[15]陈选阳,袁照年,张招娟.基因工程改良甘薯的研究与展望[J].中国农学通报,2006,22(1):21-25
[16]BallicoraM A,Iglesias A A,Preiss J.ADP-glucose pyro2 phosphorylase:a regulatory enzyme for plant starch synthesis[J].Photosynthesis research,2004,79(1):1-24
[17]刘霞,尹燕枰,贺明荣,等.播期对小麦品种藁城 8901籽粒淀粉合成相关酶活性及淀粉组分积累的影响[J].作物学报,2006,32(7):1063-1070
[18]Vandeputte G E,DelcourJ A.From sucrose to starch granule to starch physical behavior:a focus on rice starch[J].Car2 bohydrate Polymers,2004,58:245-266
[19]Keeling P L,Bacon P J,Holt D C.Elevate temperature re2 duce starch deposition in wheat endosper m by reducing the activity of soluble starch synthase[J].Planta,1993,191:342-349
[20]Xiu-qing Li,Da-peng Zhang.Gene expression activity and pathway selection for sucrose metabolism in developing storage root of s weet potato[J].Plant cell physiol,2003,44(6):630-636
[21]YatomiM,Kubota F,Saito K,et al.Evaluation of root sink a2 bility of s weet potato(Ipomoea batatas Lam.)cultivars on the basis of enzymatic activity in the starch synthesis path2 way[J].Journal of Agronomy and Crop Science,1996,177(1):17-23.
Dynamic Characteristics of Carbohydrate Synthesis and Accumulation in Tuberous Roots of Sweet Potato
Lv Changwen1,2Wang Jichun1,2Tang Daobin1,2Wang Sangen1Zhao Yong1,2Li Yanhua1
(College ofAgronomy and Biotechnology,SouthwestUniversity1,Chongqing 400716)
(Chongqing Engineering Center of Sweet Potato2,Chongqing 400716)
Three sweet potato varieties of different drymatter contentwere used to research the changes of car2 bohydrates accumulation and correlated enzymes activities,as well as the relationship among these traits.Results:As an inherent character,the difference of dry matter content or starch content between varieties mainly depend on the variety genetic characteristics.As for the varieties of middle or high dry matter,the starch accumulation is relatively fast at late stages.The activity of SPS is consistentwith the amountof starch accumulation;the activity of SS of variety Nanshu-88 is the highest among the three varieties,and its amount of starch accumulation is also the most.So SPS and SS could promote starch synthesis and accumulation.ADP-glucose pyrophosphorylase activity trends to decline with temperature lowering.In addition,there is an extremely significant negative correlation between soluble sugar con2 tent and drymatter content,and between soluble sugar content and starch content,for all the varieties.
s weet potato,carbohydrate,synthesis and accumulation,dynamic change
S531
A
1003-0174(2011)02-0023-05
重庆市自然科学基金 (CSTC2009BB5295),重庆市科技创新能力建设项目(CSTC2009CB1001)
2010-03-29
吕长文,男,1978年出生,博士,讲师,作物栽培与耕作
王三根,男,1954年出生,教授,博士生导师,植物生理生化
