实验性高原生物骨骼肌超微结构响应的运动学意义研究
2010-12-21聂秀娟
贾 磊,聂秀娟,方 梅
(1.黄山学院 体育系,安徽 黄山 245041;2.甘肃农业大学 动物医学院,甘肃 兰州 730070)
实验性高原生物骨骼肌超微结构响应的运动学意义研究
贾 磊1,聂秀娟1,方 梅2
(1.黄山学院 体育系,安徽 黄山 245041;2.甘肃农业大学 动物医学院,甘肃 兰州 730070)
通过对高原生物和小鼠运动实验,证实在高原缺氧环境或运动条件下,骨骼肌超微结构的生理响应均沿着线粒体数目增多,体积增大,肌糖原积累、红细胞数增加、载氧量增强的轨迹演变。这不仅说明研究高原生理学对探讨运动生理学有重要意义,而且也为高原训练提供了理论依据。
高原生物;骨骼肌;超微结构;线粒体;肌糖原;红细胞
为了挑战奥运,提高人体体能,扩大人类生存空间,现在越来越多的科学家把目光投向了高原生物研究。[1,2]因为高原独特的环境对机体的影响与运动时的生理响应在很多方面是相似的。如我国青藏高原平均海拔3500m以上,空气稀薄,大气压低,大气氧含量不足海平面的60%。这与运动时,尤其是大强度运动时带来的供氧不足,甚至自由基伤害造成的疲劳和运动能力下降几乎有异曲同工的效应。[3-6]因此,研究高原动物对环境的生理响应与研究运动抗疲劳和提高运动能力有着重要的价值和巨大的潜能,对进一步研究高原训练也有着重要的理论和现实意义。
本实验采用在我国青藏高原长期适应演化形成的特有动物牦牛(Bos grunniens)作为高原生物的代表,并应用犬科动物犬(Canis familiaris)在高原生活适应性和小鼠运动实验作对比,从骨骼肌超微结构的层次研究了与运动有关的内容。现就骨骼肌超微结构的响应报道如下。
1 材料与方法
牦牛选自青海3000m以上自然放牧的健壮成年牛(3头)。狗选自青海达日县(海拔4000m左右)牧民饲养的2岁犬3只。小鼠采用一月龄清洁级昆明种小白鼠20只 (购自甘肃省医学科学研究院实验动物中心),雌雄各半,体重18-20g,随机分为对照组和实验组,实验组每天进行一次游泳锻炼,方法按参考文献[11]进行,第22天做竭力运动后处死。处死和屠宰后的动物取大腿骨骼肌5×5mm3组织块置于2%戊二醛固定液中,在4℃固定1-2h,然后用缓冲液反复洗涤2-3h(中间换液3次),再加入1%锇酸固定2h,经缓冲液冲洗3次,每次15min,各级酒精充分脱水,环氧树脂(Epon-812)包埋,超薄切片机切片,铀铅染色。透射电镜观察、照相。图片采用图像分析系统测量、分析。
2 实验结果
2.1 牦牛骨骼肌对高原低氧环境的响应
牦牛骨骼肌对高原低氧环境的结构响应主要表现有:肌细胞排列紧密;线粒体数目增多,甚至有大型线粒体出现(如图1);肌糖原在肌细胞中积累,而以肌节四周甚至在Z线附近比较集中(如图2);毛细血管增加,红细胞数增多,直径较大(如图3)。

图1 牦牛骨骼肌。示肌细胞排列紧密,线粒体数目多,体积大,肌糖原积累明显

图2 牦牛骨骼肌。示肌糖原积累较多且主要分布在肌节四周及Z线附近

图3 牦牛骨骼肌。示毛细血管中红细胞以及凋亡小体
2.2 犬骨骼肌对高原低氧环境的响应
高原缺氧环境对犬骨骼肌超微结构的影响也是极为深刻的,如细胞排列疏松、胞间隙增大(图4);线粒体增多、体积变大,甚至出现巨型线粒体或空泡化 (图4,5);肌糖原明显增多,也表现出在肌节四周或Z带附近分布较多 (图6);次级溶酸体活跃(图5),说明对线粒体更新有积极作用;局部有肌丝排列疏松、Z带扭曲和肌溶灶出现(图7-9)。

图4 犬骨骼肌。示肌细胞排列疏松、间隙增大,线粒体增多

图5 犬骨骼肌。示线粒体增大并有空泡化,并伴有次级溶酶体

图6 犬骨骼肌。示肌糖原积累,在肌节四周及Z线附近分布较多

图7 犬骨骼肌。示肌丝松散

图8 犬骨骼机。示Z带扭曲

图9 犬骨骼肌。示肌溶灶
2.3 小鼠骨骼肌对高原低氧环境的响应
小鼠游泳实验,因为加大了对氧供的要求,骨骼肌超微结构同样也出现了线粒体增多、体积加大,甚至出现空泡(图10);肌糖原明显增加,也多集中在肌节四周和Z线附近(图11);局部地区Z带略有弯曲(图12)和肌丝松散现象(图13。

图10 实验小鼠骨骼肌。示线粒体增大并有空泡化 25000

图11 实验小鼠骨骼肌。示肌糖原积累,主要分布在肌节四周及Z线附近50000×
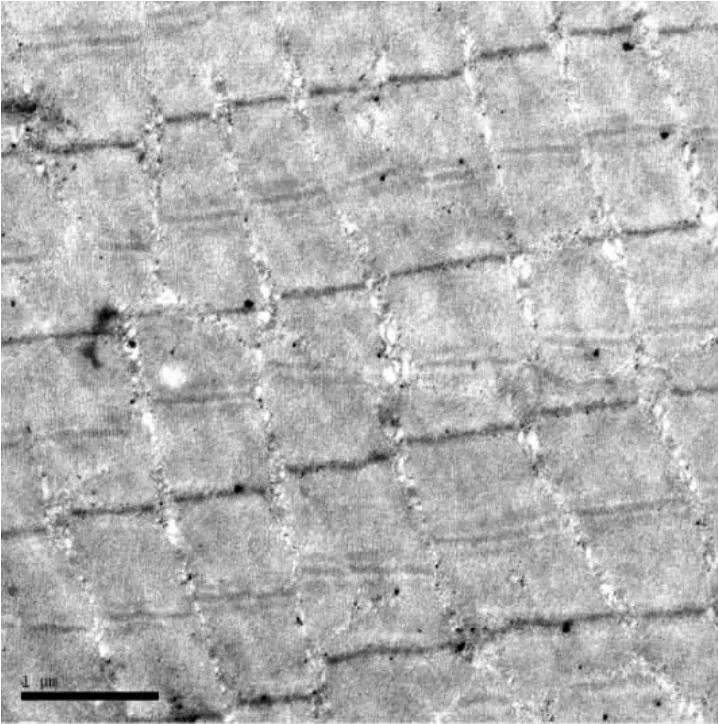
图12 实验小鼠骨骼肌。示Z线略有弯曲,个别地方Z线消失25000×

图13 实验小鼠骨骼肌。示肌丝松散20000×
3 讨 论
不少研究表明,环境或运动疲劳与骨骼肌超微结构、功能的变化直接相关。这些变化主要包括敏感细胞器线粒体的积极响应,能源物质的积累,[1,7-11]骨骼肌纤维结构的改变和自由基、ATP等对细胞凋亡的启动。
3.1 线粒体的响应
牦牛生活在海拔3000m以上的青藏高原,同海平面相比,3000m处空气含氧量减少1/3,5000m处约减少1/2。西藏东部海拔1000m以下地方空气含氧量为250-280g/m3,3000m左右降为200g/m3,4000-5000m处仅为160-180g/m3。[12]为了解决供氧供能,骨骼肌的结构响应首先表现在线粒体上。
线粒体是生物氧化、产生能量的主要场所。三羧酸循环、呼吸链上的电子传递以及磷酸化都在线粒体上进行。[13]糖、脂肪和蛋白质等有机物先在细胞质中进行部分氧化,然后进入线粒体,通过三羧酸循环彻底氧化,脱下来的氢和电子通过呼吸链依次传递给氧,最后电子和H+结合氧形成水。在这个过程中释放的能量以高能磷酸键(ATP)储存在有机体内供生物做功,包括肌肉收缩。[14,15]另外,线粒体在遗传上具有独立性,可以自我复制,也可通过出芽、自缢进行繁殖。所以在氧缺乏的环境里,生物可以通过线粒体的快速反应以保证能量的需要。反映在超微结构上就出现了数量增多、体积增大。
3.2 肌糖原的积累
机体的能源物质有糖类、脂肪和蛋白质,其中糖类可被各种组织直接利用,称为直接燃料。而脂肪和蛋白质则需要分别转变成脂肪酸、甘油和氨基酸才能被氧化利用。糖在体内转变成糖原而储存。肌肉是人体耗能最多的组织,肌肉收缩做功主要靠肌糖原氧化供给。[16,17]故在缺氧需能的矛盾中,糖原在肌肉中积累,也是为了快速提供能源物质。
关于糖原在肌节周围和Z线附近较多,其机理与肌丝运动有关。因为,骨骼肌纤维收缩做功是肌原纤维内肌丝的运动所致,即细肌丝在粗肌丝之间向M线方向滑动,结果使肌节变短,肌原纤维收缩,反之舒张。在此过程中均要由ATP提供能量,而能源物质则以糖原最佳,其分布则以肌节周围和Z线附近提供能量速度最快。这也反映了机体对缺氧环境科学、快速的响应。
3.3 供氧系统的改善
有报道称,[18]动物生活在高原环境中毛细血管密度增加。牦牛不仅毛细血管多,而且红细胞数量多,且直径大(图2),据杰尼柯夫(Денисов.В.Ф)报道,红细胞平均直径,牦牛为4.83μ,同龄的吉尔吉斯牛(普通牛)仅为4.33μ。[12]说明同一单位血液中的氧容量牦牛远高于普通牛。另外,从外观看,牦牛肌肉紧密、色泽鲜红。紧密说明肌纤维发达,色泽鲜红是因肌红蛋白含量高、含氧量大所致,它和血红蛋白构成了机体组织内氧的供应系统,这对骨骼肌在低氧环境下正常工作具有重要作用。
3.4 细胞凋亡的启动
细胞凋亡是指机体在生理条件下受到刺激后,经过多种途径的信号传递导致细胞产生一系列形态和生化方面的改变而引起细胞自杀性死亡的过程。这是一种机体消除损伤、衰老、突变的细胞,有利于机体维持正常生理平衡,完全不同于死亡的一种凋亡途径。[19-21]细胞凋亡是受到高度调节的一种生理性细胞死亡过程,是受基因调控的一个主动连续的程序化反应。其形态学上的改变主要包括细胞核染色质固缩、边集,细胞膜破裂形成凋亡小体,[22]线粒体基质由于高渗面膨胀破裂、电子传递受阻、ATP产量骤减,无法维持供能,细胞走向凋亡。
基于本文研究的高原生物和小鼠肌细胞核的变化、线粒体出现肿胀、增大等的变化及肌丝排列出现紊乱等改变,可以认为,高原环境或剧烈运动因缺血、缺氧等因素启动了骨骼肌细胞的凋亡过程。而这一过程在一定程度上可能限制细胞坏死,并有利于骨骼肌的修复。同时,骨骼肌的细胞凋亡与缺血、缺氧等应激因素及运动的时间与强度有很大的相关性。[23-25]袁箭峰采用流式细胞术对不同强度耐力训练及力竭运动大鼠心肌细胞凋亡情况进行了活细胞观察,[26]结果显示中等强度训练组凋亡比率与对照组相比增加不明显,大强度训练组及力竭组与对照组相比显著增加,说明运动所造成的缺氧性应激因素可诱导心肌细胞凋亡,且凋亡发生比率随运动强度的增大而增加。丁延峰等在进行缺血、缺氧引起心肌细胞凋亡的研究中证实了心肌细胞的凋亡是不同于细胞坏死的一种死亡方式,并且提出了缺血、缺氧之前的预处理有助于减轻心肌细胞的凋亡。[27]可见,细胞凋亡的启动也是骨骼肌对高原环境或过度训练的一种保护性响应。只有当环境条件或运动量超过了一定阈值时,这时细胞就会出现坏死。但骨骼肌细胞对低氧环境的感受机制是十分复杂的,不同形式的缺氧刺激可能触发不同的机制,所以不能简单认为缺氧、运动诱导骨骼肌细胞凋亡的反应是相同的。有关骨骼肌细胞凋亡的机理还有待进一步的深入研究。
3.5 高原生物适应性和小鼠运动实验骨骼肌响应
的雷同性
通过犬的高原适应性实验和小鼠的运动实验证实,机体对高原缺氧和运动缺氧有着同样的生理响应,反应在骨骼肌超微结构的变化上与牦牛雷同。如线粒体数量增多、体积增大,同时肌糖原积累,且也在肌节四周和Z线处分布较多。但是环境变化的剧烈,如犬在4000m左右生活和小鼠过量的运动,使骨骼肌出现了损伤,[3]在结构上有Z带扭曲、肌丝松散、甚至出现肌溶灶。
综上所述,以上实验不仅阐明了高原生物骨骼肌超微结构的特点,而且证实了这种生理学响应的运动学意义,同时也为高原训练从机理上提供了依据:
1.证明高原训练确实可以增加人体对缺氧的抵御能力和适应性;
2.高原训练生理响应最快的是线粒体和肌糖原;
3.机体的适应能力与环境变化的大小和运动剧烈程度相关。
这也说明了高原训练海拔高度的选择、训练的强度、时间等安排必须科学合理。所以,只有选用适当的海拔环境、通过科学的训练方式,供给相应的营养(糖原等),运动员的运动能力才会得到相应的提高。
[1]贾磊.挑战奥运竞技体能的分子机理研究与应用[C].首届中国体育博士高层论坛文集,2006:63-65.
[2]马生林.青藏高原生物多样性保护研究[J].青海民族学院学报(社会科学版),2004,30(4):76-78.
[3]贾磊,段玉虎.过度训练与自由基伤害[J].兰州大学学报,2001,29:230-233.
[4]JIL L,WU E.THOMAS DP.Effect of exercise training on antioxidant and metabolic functions in senescent rat skeletal muscle[J].Gerontology,1991,37(6):317-325.
[5]HAMMEREN J.POWERS S,LAWLEV J,et al.Exercise training-inducdalter 2 actions in skeletal muscle oxidative and antioxidant enzyine activity in senescent rats[J].Int J Sports Med,1993,13(5):412-416.
[6]KOSHELEV VB.TARASOVA OS,STOROZHEVYKH TP, et al.Changes in the systemic hemodynamics and the vascular bed of the skeletal muscles in rats adapted to hypoxia[J].Hyp Mcd J,1995,(2):16-19.
[7]刘燕萍.运动性疲劳及其机制分析[J].西安体育学院学报, 2001,18(1):46-47.
[8]周国璋.运动性疲劳的预防与恢复[J].中国临床康复,2002,6 (9):1331-1332.
[9]田野,杨锡让.运动性骨骼肌的结构机能变化的机制研究-I力竭性运动对骨骼肌纤维超微结构的影响[J].中国运动医学杂志,1992,11(4):202-204.
[10]王长青,刘丽萍,郑师陵,等.运动性疲劳时Ca2+、线粒体膜电位的改变与细胞凋亡[J].体育科学,2000,20(3):59-63.
[11]贾磊,郭层城.杜仲对骨骼肌超微结构的改善与提高运动能力的研究[J].体育科学,2003,23(4):108-110.
[12]张容昶.中国的牦牛[M].兰州:甘肃科学技术出版社,1989: 29-74.
[13]左伋.医学分子细胞生物学[M].上海:复旦大学出版社, 2005:15-52.
[14]EdwardsRHT.In:PoterR.WheianJ.eds.Human Muscle Fatigue:Physiological Mechanisms [M].London: Pitman Medcal,1981:297-301.
[15]Hermansen L.In:PoterR.Wheian J.eds.Human Muscle Fatigue:Physiological Mechanisms [M].London: Pitman Medcal,1981:75-82.
[16]王步标,华明,邓树勋.人体生理学[M].北京:高等教育出版社,1996:244-250.
[17]沈同,王镜岩.生物化学[M].北京:高等教育出版社,1998: 38-92.
[18]Hudicka O.Growth of capillaries in skeletal and cardiac muscle[J].Res.1982,50:451-461.
[19]贾磊.运动应激与细胞凋亡[J].兰州大学学报(医学版), 2007,33(4):72-76.
[20]方晓阳,盛伟.细胞凋亡的概念来源及其研究进展[J].生命的化学,2003,23(2):121-123.
[21]MEIER P, FINCH A, EVAN G. Apoptosis in development[J].Nature,2000,407(6805):796-801.
[22]JIA LEI,LI-ZHE AN.Effects of the Extract of Ammopiptanthus mongolicus cheng f. (JAI)on Induction of Apoptosis of HepG2 in vitro and Its Molecular Mechunisms[J].Biomedical and Environmental Sciences,2006, 19(2):118-123.
[23]SUSIN S A,LORENZO H K,ZMAZAMIC N,et al. Molecular characterization of mitochondrial apoptosis-inducing factor[J].Nature,1999,397(4):441-445.
[24]沈志祥.运动与细胞凋亡 [J].山西师大体育学院学报, 2004,19(3):80-82.
[25]贺洪,唐晖,汪保和.运动与细胞凋亡的研究进展[J].浙江体育科学,2000,22(6):42-43.
[26]袁剑峰.运动心脏重塑过程中细胞凋亡现象的活细胞观察[J].中国运动医学,2001,20(4):301-351.
[27]丁延峰.缺血预处理减轻在体家兔心肌细胞凋亡[J].生理学报,2000,(3):220-240.
A Study on Kinematic Significance of Skeletal Muscular Ultrastructure Respondence in Experimental Altitude Organism
Jia Lei1,Nie Xiujuan1,Fang Mei2
(1.Department of Physical Education,Huangshan University,Huangshan 245041,China;
2.College of Veterinary Medicine,Gansu Agricultural University,Lanzhou 730070,China)
With altitude organism and rats as experimental subject,the physiological respondence of their muscular ultrastructure was investigated.The results showed that the physiological respondence of their muscular ultrastructure developed along the path of mitochondria increase both in number and in size, muscle glycogen and erythrocytosis accumulation,as well as transport oxygen capability enhancement after hypoxia exposure or sports.The results not only proved the significance of researching into altitude physiology for sports physiology but also provided theoretical evidence for altitude training.
altitude organism;skeletal muscle;ultrastructure;mitochondria;muscle glycogen; erythrocytosis
G804.1
A
1672-447X(2010)04-0098-05
2010-03-15
安徽省教育厅自然科学研究项目(KJ2009B274Z);黄山学院引进人才启动基金项目(2008xskq013)
贾 磊(1971-),湖南岳阳人,黄山学院体育系副教授,博士,研究方向为运动分子生物学及运动生理学。
胡德明
