galc与头颈部肿瘤的研究进展
2010-11-15综述陈小毅审校
陈 芳 综述 陈小毅 审校
鞘糖脂类包括半乳糖(基)脑苷脂(galactosylcerebroside,GalCer)和半乳糖(基)鞘胺醇(galactosylsphingosine,psychosine)及其它,主要由半乳糖(基)脑苷脂酶(galactosylceramidase,GALC)降解为半乳糖、脑苷脂和鞘胺醇。Galactosylceramidase gene,简称galc,负责编码GALC。研究发现癌细胞表面的鞘糖脂类含量增加,这在抑制细胞黏附和凋亡方面影响很大[1]。鞘糖脂类含量增加的原因,除了发现头颈部鳞状细胞癌(head and neck squamous cell carcinoma,HNSCC)中galc基因转录受抑制,应用抗肿瘤药物可以促进肿瘤细胞里的神经酰胺糖基化,生成鞘糖脂类外,其它均无发现[2,3]。目前国内外关于galc与恶性肿瘤的关系的研究极少,但现有的研究结果提示galc在肿瘤研究中可能很有价值,值得深入研究。
鼻咽癌是华南地区最常见的恶性肿瘤之一[4],在人类HNSCC中发病居首位,研究发现,galc在多种头颈部肿瘤细胞系中转录受抑制、表达下调并伴甲基化,抗甲基化药物处理后表达增加,推测galc可能在头颈部恶性肿瘤的发生发展中发挥重要作用。研究发现癌细胞中GalCer过表达,可能与galc的转录受抑与甲基化有关[1],而且galc的表达异常可能与头颈部肿瘤细胞凋亡抑制、肿瘤发生密切相关,galc有望成为肿瘤靶向治疗的新靶点。本文就galc的结构、生物学功能以及galc与头颈部肿瘤的研究现状做一综述。
1 galc的结构及生物学功能
1.1 galc的结构及与转录因子的关系
galc是1种看家基因,位于14q31,大小约60 kb,包括17个外显子和16个内含子,外显子除了第1个和最后1个都相对较小,范围从39到181个核苷酸。内含子大小差异较大,从247个核苷酸到大约12 kb。与其他编码溶酶体蛋白的基因相似,其5’非翻译区富含GC,不含CAAT或者TATA序列。其启动子区显示有对转录因子sp1(50-CCCGCC-30)、yy1(50-AAATGG-30)和ap2(50-GCCTGCAGGC-30)易感的信号序列。即使有些因子作用的启动区域稍微不同,但是却大体相似,以此推测galc转录受抑制可能是抑制性转录因子结合到了合适的启动子上导致galc转录受抑制,进而导致溶酶体酶GALC表达减少,最终导致细胞膜上的GalCer的大量增加。如果能找到sp1、yy1和ap2识别galc转录启动子的相关区域,就可以解释galc的抑制。SP1虽然是最有效的转录因子,它并不会抑制galc,因为SP1的作用主要是增强与之相结合的基因的功能,而非抑制。yy1和ap2都可能抑制 galc,据报道 yy1 还能抑制 p53[5~9]。
1.2 galc的组织表达与生物学功能
galc具有很强的组织表达特异性,在鞘磷脂中大量表达,尤其是在脑白质中,在肾脏、小肠和大肠中含量也很丰富。galc可以从人脑或人尿中提取,也可以从胎盘组织提取,因为在胎盘组织中也很丰富。galc转录后出核膜,在细胞质中翻译成GALC。galc编码的GALC,又称半乳糖(基)神经酰胺酶(Galactosylceramidase,也简称GALC),是1种溶酶体的β-半乳糖苷酶,能分解GalCer、psychosine以及其他糖鞘脂类物质为半乳糖、神经酰胺和鞘胺醇,避免这些物质在体内蓄积。GALC由1分子的半乳糖以糖苷键结合于神经酰胺上形成,是两性分子,疏水端由2个长链脂肪酸组成并黏附在细胞膜内,亲水端由丝氨酸和半乳糖组成位于膜外。除了与胞外基质的交流外糖鞘脂类还有多种生物学功能包括黏附、细胞生长和分化、凋亡和表面抗原[10]。研究发现,GalCer具有凋亡抑制作用,其在细胞内的蓄积可以促使细胞黏附减少,抑制细胞凋亡,促进细胞增殖和迁移并延长细胞寿命,进而导致肿瘤形成,而其产物神经酰胺则有促进凋亡的作用。GalCer的结构图如下所示:
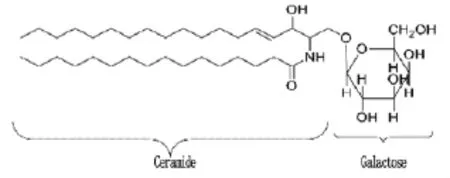
2 galc与鼻咽癌
2.1 galc在鼻咽癌组织中的表达及临床意义
galc在正常鼻咽组织和慢性炎组织中存在高表达现象,而在鼻咽癌组织中基本不表达,这提示galc可能在HNSCC的发生、发展中发挥重要作用。研究发现,在HNSCC的癌细胞表面存在着GalCer的过表达,提示galc的表达失活可能与头颈部肿瘤的发生发展相关,Gorogh等在研究中除了发现GalCer过表达外,还发现galc的转录受抑,推测GALC失活很可能是GalCer在肿瘤细胞蓄积的机制之一。
2.2 galc表达下调的可能机制
galc表达下调的可能机制主要有以下几种:①galc的启动子区:若galc启动子区能特异性结合某些转录抑制因子将导致其转录受抑,翻译水平随之下降,GALC功能降低。研究发现AP2和YY1可能都是galc的转录抑制因子[8,9]。②基因突变:galc启动子区和编码区的基因突变和缺失都会导致galc的转录下降。虽然对喉癌中galc的5’侧翼区进行序列分析,结果未检测到突变和缺失,但该序列分析不能排除编码区的突变,因为只有受抑制的galc的mRNA逆转录出的cDNA中碱基对从1749到1897中的一小部分被检测到,不包含galc的编码序列。③galc甲基化:甲基化可导致基因不稳定、突变率升高,基因表达异常。甲基化作用无论是发生在启动子区还是在编码区,都会影响DNA的转录。低度的甲基化使DNA对激活转录的蛋白质更敏感,而过度甲基化则使转录受抑[11~15]。人类肿瘤细胞中,DNA甲基化导致的基因沉默是1种常见现象[16]。④翻译后失活:迄今为止,除了HNSCC的研究中报道过galc表达失活外,其他肿瘤中尚无相关报道,所以更有可能是GALC或其它相关酶类翻译后失活。鞘脂激活蛋白的酶组包括A、B、C、D 4组,作用都很重要,其中A和C能激活并增强 GALC的功能,所以A和C受抑无论是到功能的还是源于基因突变,都会导致GALC功能的降低[15,16]。总之,不管通过那种方式引起的GALC表达减少、功能降低,最终都将导致其底物GalCer不能被有效降解而在细胞膜大量蓄积。
3 galc甲基化与头颈部肿瘤
甲基化导致的基因表达失活是头颈部肿瘤中的1个常见的分子机制,与其发生发展密切相关,许多与之相关的基因都存在异常甲基化现象。例如,p16ink4a甲基化是该基因失活的重要机制,与头颈部肿瘤的发生密切相关[17]。cdh1编码的E-钙黏素在细胞黏附系统中起重要作用,该基因的异常表达可导致肿瘤的侵袭及转移。研究发现,该基因在头颈部肿瘤中存在高甲基化现象,而在相应的正常组织中无甲基化。还有研究发现,p14、p15、p73、hMLH1、MGMT、DAP 激酶等基因的失活都与启动子甲基化有关[18~20]。郭忠民等对多种头颈部肿瘤细胞进行了检测,结果发现GALC甲基化在头颈部肿瘤中非常常见,且GALC的表达水平与其甲基化的程度呈反比,甲基化水平越高,GALC的表达相对越低。同时,他们利用药物逆转galc的甲基化作用后,发现galc的表达上调,证明galc表达受抑和失活极有可能是其发生甲基化的结果。
4 galc-targeted抗肿瘤治疗
肿瘤的发生发展不仅是细胞增殖失控和分化异常的结果,而且与细胞凋亡失衡有关。诱导肿瘤细胞凋亡已成为目前肿瘤治疗主要手段之一。
4.1 GALC的底物、产物与细胞凋亡
GALC的底物鞘糖脂在癌细胞表面的蓄积会导致肿瘤形成相关的糖链抗原的抗原属性显著改变、黏附降低、接触性生长抑制减弱、运动增强、侵袭性增强更容易转移。除此之外,他们还为细菌、病毒、毒素提供了结合位点。GalCer抑制凋亡,然而它被GALC水解后的产物神经酰胺却促进凋亡,如此看来,galc受抑制不止是HNSCC癌变过程的指示剂,还是1个促进剂。
神经酰胺,GalCer代谢的1个产物,是研究最多的1种鞘脂,因潜在诱导凋亡而著称。Chi等通过调查健康人的喉黏膜、黏膜白斑病和喉癌患者的喉黏膜发现其神经酰胺的含量是逐级递减的[21]。因为神经酰胺与细胞凋亡有关,所以推测它的含量逐级递减可能会导致喉癌的发生。而神经酰胺的产量会受到外界刺激的影响,例如:TNF-,内毒素,INF-,IL-1,抗肿瘤药,高温,电离辐射等。这些因素都可通过促进神经酰胺的产生诱导细胞凋亡。Reynolds等通过动物模型发现放疗和化疗会通过增强从头合成和鞘磷脂水解两方面提升神经酰胺的含量[3]。神经酰胺作为凋亡促进剂已经应用在肿瘤治疗研究中。神经酰胺在细胞内主要由从头合成和鞘磷脂水解(分解合成)2条途径产生,其含量主要依靠代谢相关酶的活性。GALC就是鞘磷脂水解途径中的1个很重要的酶,GALC把GalCer降解成神经酰胺,可谓一举两得。所以可以设想,建立以galc为靶点的肿瘤治疗方案,从而达到诱导肿瘤细胞凋亡的目的。
4.2 galc甲基化的治疗
DNA甲基化对于肿瘤是1种简便而灵敏的检测指标,DNA甲基化异常不仅可在手术标本中检测到,还可从各种体液如唾液、外周血清等检测到,为临床应用带来极大便利,有可能成为将来检测头颈部肿瘤的常用方法。而且由于DNA甲基化所致的表达异常是可以逆转的,故通过恢复仅被抑制的基因的表达,即可恢复细胞正常生长调控功能,有可能达到治疗肿瘤的目的。大量体外实验证实,应用甲基转移酶抑制剂或去甲基化药物处理后,可使表达受抑的基因重新表达[22]。因此,以galc甲基化为靶点的治疗极有可能为肿瘤治疗带来希望。所以,深入研究galc失活的机制可能为头颈部肿瘤的诊断和治疗提供新的路径。
综上所述,在肿瘤研究中galc是1个比较新颖的基因,目前国内外关于galc与恶性肿瘤关系的研究极少,但现有的研究结果暗示galc在HNSCC的研究中是1个非常有价值的基因。galc的失活能导致鼻咽癌的发生,那它的激活是否能抑制鼻咽癌发生;galc与其他有关癌基因、抑癌基因和凋亡相关基因的关系如何;对GALC的检测能否应用到鼻咽癌的诊断;这些均需进一步探讨。
[1]Beier UH,Görögh T.Implications of galactocerebrosidase and galactosylcerebroside metabolism in cancer cells〔J〕.Int J Cancer,2005,115(1):6.
[2]Gouaze V,Yu JY,Bleicher RJ,et al.Overexpression of gluco-sylceramide synthase and P-glycoprotein in cancer cells selected for resistance to natural product chemotherapy〔J〕.Mol Cancer Ther,2004,3(5):633.
[3]Reynolds CP,Maurer BJ,Kolesnick RN,et al.Ceramide synthesis and metab-olism as a target for cancer therapy〔J〕.Cancer Lett,2004,206(2):169.
[4]闵华庆,洪明晃,郭 翔,鼻咽癌〔M〕.第1版.北京:中国医药科技出版社,2003:16~30.
[5]Li L,He S,Sun JM,et al.Gene regulation by Sp1 and Sp3〔J〕.Biochem Cell Biol,2004,82(4):460.
[6]Sugawara T,Nomura E,Nakajima A,et al.Characterization of binding between SF-1 and Sp1:predominant interaction of SF-1 with the N-terminalregionof Sp1〔J〕.J Endocrinol Invest,2004,27(2):133.
[7]Mansilla S,Priebe W,Portugal J,et al.Sp1-targetedinhibitionofgenetran-scription by WP631 in transfected lymphocytes〔J〕.Biochemistry,2004,43(23):7584.
[8]Sui G,Affar el B,Shi Y,et al.Yin Yang 1 is a negative regulator of p53〔J〕.Cell,2004,117(7):859.
[9]Yakovleva T,Kolesnikova L,Vukojevic V,et al.YY1 binding to a subset of p53 DNA-target sites regulates p53-dependent transcription〔J〕.Biochem Biophys Res Commun,2004,318(2):615.
[10]Yang J,Yu Y,Sun S,et al.Ceramide and other sphingolipids in cellular responses〔J〕.Cell Biochem Biophys,2004,40(3):323.
[11]Badal S,Badal V,Calleja-Macias IE,et al.The human papillomavirus-18 genome is efficiently targeted by cellular DNA methylation〔J〕.Virology,2004,324(2):483.
[12]Huttley GA.Modeling the impact of DNA methylation on the evolution of BRCA1 in mammals〔J〕.Mol Biol Evol,2004,21(9):1760.
[13]Chan Y,Fish JE,D'Abreo C,et al.The cell-specific expression of endothelial nitric-oxide synthase:a role for DNA methylation〔J〕.J Biol Chem,2004,279(33):35087.
[14]Jones PA,Baylin SB.The epigenomics of cancer〔J〕.Cell,2007,128(4):683.
[15]Matsuda J,Vanier MT,Saito Y,et al.A mutation in the saposin A domain of the sphingolipid activator protein(prosaposin)gene results in a late-onset,chronic form of globoid cell leukodystrophy in the mouse〔J〕.Hum Mol Genet,2001,10(11):1191.
[16]Schuette CG,Pierstorff B,Huettler S,et al.Sphingolipid activator proteins:proteins with complex functions in lipid degradation and skin biogenesis〔J〕.Glycobiology,2001,11(6):81R.
[17]Shintani S,Nakahara Y,Mihara M,et al.Inactivation of the p14(ARF),p15(INK4B)and p16(INK4A)genes is a frequent event in human oral squamous cell carcinomas〔J〕.Oral Oncol,2001,37(6):498.
[18]Esteller M,Corn PG,Baylin SB,et al.A gene hypermethylation profile of human cancer〔J〕.Cancer Res,2001,61(8):3225.
[19]Araki D,Uzawa K,Watanabe T,et al.Frequent allelic losses on the short arm of chromosome 1 and decreased expression of the p73 gene at 1p36.3 in squamous cell carcinoma of the oral cavity〔J〕.Int J Oncol,2002,20(2):355.
[20]Liu K,Huang H,Mukunyadzi P,et al.Promoter hypermethylation:an important epigenetic mechanism for hMLH1 gene inactivation in head and neck squamous cell carcinoma〔J〕.Otolaryngol Head Neck Surg,2002,126(5):548.
[21]Chi FL,Yuan YS,Wang SY,et al.Study on ceramide expression and DNA content in patients with healthy mucosa,leukoplakia,and carcinoma of the larynx〔J〕.Arch Otolaryngol Head Neck Surg,2004,130(3):307.
[22]Li QL,Ito K,Sakakura C,et al.Causal relationship between the loss of RUNX3 expression and gastric cancer〔J〕.Cell,2002,109(1):113.
