高山红景天中糖基转移酶家族cDNA全长基因的克隆
2010-10-19于寒松张继星李彦舫马兰青胡耀辉
于寒松,张继星,李彦舫,马兰青*,胡耀辉,*
(1.吉林农业大学食品科学与工程学院,吉林 长春 130118;2.内蒙古民族大学生命科学学院,内蒙古 通辽 028043;3.吉林大学植物科学学院,吉林 长春 130062;4.农业部都市农业(北方)重点开放实验室,北京农学院,北京 102206)
高山红景天中糖基转移酶家族cDNA全长基因的克隆
于寒松1,张继星2,李彦舫3,马兰青4,*,胡耀辉1,*
(1.吉林农业大学食品科学与工程学院,吉林 长春 130118;2.内蒙古民族大学生命科学学院,内蒙古 通辽 028043;3.吉林大学植物科学学院,吉林 长春 130062;4.农业部都市农业(北方)重点开放实验室,北京农学院,北京 102206)
为了获得高山红景天中红景天苷生物合成关键酶——UDP-糖基转移酶家族基因的全长cDNA序列,采用在线简并引物设计软件结合3′-RACE技术获得两个糖基转移酶基因的3′端部分序列,进一步采用不同的5′-RACE试剂盒扩增两个基因的5′端序列,结果显示利用Takara公司的试剂盒扩增可以得到cDNA全长序列(GenBank登录号为EF508689和EU567325),而采用Invitrogen公司的试剂盒得到的序列不包括全部开放读码框。实验证明,利用Takara公司的SMARTerTMRACE cDNA Amplification Kit克隆获得基因全长序列的比率更高。
糖基转移酶;基因克隆;cDNA末端快速克隆法(RACE);高山红景天
Abstract:In order to obtain the critical enzyme for the synthesis ofRhodiola sachalinensisglycoside, two putative UDP-glycosyltransferase (UGT) cDNAs (GenBank accession number, EF508689和EU567325) were isolated fromR. sachalinensi.The primers were designed for 3'-rapid amplification of cDNA ends (RACE) based on the consensus hybrid oligonucleotide primers (CODEHOP) strategy by the Block Maker program. A full-length cDNA sequence (Genbank accession numbers:EF508689 and EU567325) was obtained through the amplification using a SMARTerTMRACE cDNA Amplification Kit (TaKaRa,Dalian, China), whereas the sequence obtained using a 5'-RACE system (version 2.0, InvitrogenTMLife Technologies) contained no full-open reading frame.The cloning efficient of two 5'-RACE systems was investigated. Results exhibited that TaKaRa SMARTerTMRACE cDNA Amplification Kit was more efficient in complete cDNA sequence cloning, compared with the InvitrogenTM5'-RACE system.
Key words:glycosyltransferase;gene cloning;rapid amplification of cDNA ends (RACE);Rhodiola sachalinensis
植物次生代谢物质在植物生命活动过程中起着重要作用,如细胞解毒、信号转导、防御机制等[1-3]。对人类而言,很多植物次生代谢物具有很多生理功能,也是保健品和医药品的重要来源[4]。现阶段天然产物己越来越多地被应用于保健食品、天然香料、天然色素和药物等产品的生产上[5-6]。但是,植物虽然能合成上万种次生代谢产物,但是它们的含量都很低。因此,利用现代基因工程技术获得有价值的次生代谢产物已成为科学家研究的热点。
长白山高山红景天具有明显的抗辐射、抗疲劳、抗病毒、延缓机体衰老等生理功能[7-10],主要是由于它含有红景天苷、酪萨维、红景天素和草质素苷等多种糖苷,这些物质都是糖基转移酶的催化产物[11-12],尤其是红景天的主要功能性物质——红景天苷,就是在酪醇糖基转移酶的催化下合成的。因此,以长白山高山红景天为原料,克隆糖基转移酶基因对体外合成红景天的功能性物质或上调红景天合成这些次生代谢物质的量具有重要意义。本实验以长白山高山红景天为原料,通过基因工程方法分离糖基转移酶家族的基因,同时比较两种5′-RACE试剂盒克隆未知基因5′端序列的效率。
1 材料与方法
1.1 材料与试剂
高山红景天野生植株采自吉林长白山。
SMARTerTMRACE cDNA Amplification Kit 日本Takara公司;5'RACE system for rapid amplification of cDNA ends试剂盒 美国Invitrogen公司;引物由上海生工生物工程技术服务有限公司合成;测序由上海生工生物工程技术服务有限公司和上海华大鼎安公司完成。
1.2 仪器与设备
Biometra 2000 PCR扩增仪 德国Whatman Biometra公司;MIKRO 22R高速冷冻离心机 Hettich公司;-80℃超低温冰箱 Sanyo公司;电泳仪 北京六一仪器公司;TGL 16G高速台式离心机 上海医用分析仪器厂。
1.3 方法
1.3.1 高山红景天总RNA提取
采用改进后的CTAB-LiCl法提取总RNA,具体步骤参照文献[13]。
1.3.2 引物设计
1.3.2.1 3′-RACE简并引物设计
上游引物设计:根据在GenBank上已发布的植物糖基转移酶氨基酸序列及相关文献[14-15],选取其中5个以简单酚类为催化底物的酶序列,其中包括藏红花 (登录号AY262037)、甜菊(登录号AY345982)、拟南芥(登录号AC002333)、烟草属(登录号AB052558)、甘草属(登录号AB098614)。将这些序列以FASTA格式提交到http://bioinformatics.weizmann.ac.il/blocks/blockmkr/www/make_blocks.html得到5个酶的氨基酸保守区序列,将最保守的序列提交到在线引物设计软件CODEHOP (http://bioinformatics.weizmann.ac.il/blocks/codehop.html) 中进行引物设计,得到的引物命名为引物B,序列为5′-GTT GGAGTTTTTGTTACTCAytgyggntgga-3′。
下游引物设计:根据反转录的引物Q1:5′-GAG GACTCGAGCTCAAGCTTTTTTTTTTTTT-3′设计下游引物序列为Q2:5′-GAGGACTCGAGCTCAAGC-3′。
1.3.2.2 5′-RACE特异性引物设计
根据获得的两个糖基转移酶基因的3′端序列利用Primer Primer 5.0软件分别设计5'-RACE实验巢式引物,引物序列见表1。

表1 克隆UGTC和UGTR5'端引物序列Table 1 Primer sequences for cloning 5'-end ofUGTCandUGTR
其他5′-RACE所需相关引物CDS Prime、SMART II、UPM引物为SMARTerTMRACE cDNA Amplification Kit自带引物;AAP、UAP、AUAP引物为5'-RACE system for rapid amplification of cDNA ends试剂盒自带引物。
1.3.3 PCR扩增条件
以反转录合成的cDNA第一链为模板进行3'-RACE实验,PCR条件为:95℃预变性5min;94℃ 30s,58℃45s,72℃ 1min,30个循环;72℃延伸10min。5'-RACE的PCR条件按照相应的试剂盒说明书操作。
目的基因的回收、连接、转化、鉴定均按常规方法[16]操作,序列测定由上海生工生物工程技术服务有限公司完成。
1.3.4 生物信息学分析
应用NCBI(www.ncbi.nlm.nih.gov)网站上的BLAST程序完成氨基酸序列比对和蛋白质序列同源性分析。
2 结果与分析
2.1 3′-RACE实验结果
以提取的红景天总RNA为模板,利用引物Q1进行反转录,利用引物Q2和B进行PCR,反应结束后进行琼脂糖凝胶电泳,结果显示3′-RACE实验获得两个约500bp的基因片段,见图1。所得片段与pMD18-Tsimple载体连接,转化大肠杆菌后送测序公司测序。
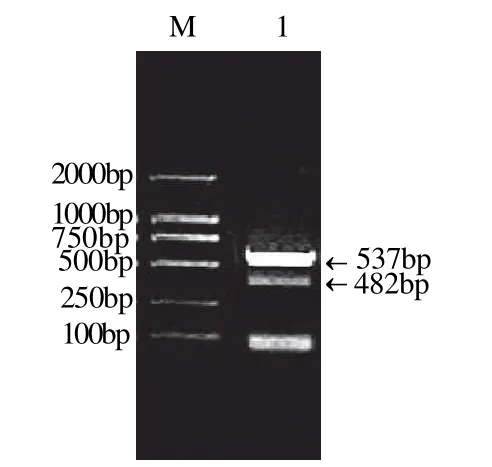
图1 3'RACE实验结果Fig.1 Results of 3'-RACE
2.2 5′-RACE实验结果
根据3′-RACE实验所得到的序列设计5′-RACE实验所需的特异性引物(表1),参照试剂盒说明书进行5′-RACE实验,反应结束后进行琼脂糖凝胶电泳。结果显示:采用Invitrogen公司试剂盒扩增得到的基因片段长度都为800bp左右,见图2、3;而采用Takara公司试剂盒扩增得到的基因片段长度在1100bp左右,见图4、5。将所得到的基因序列与T载体连接,转化大肠杆菌后测序。
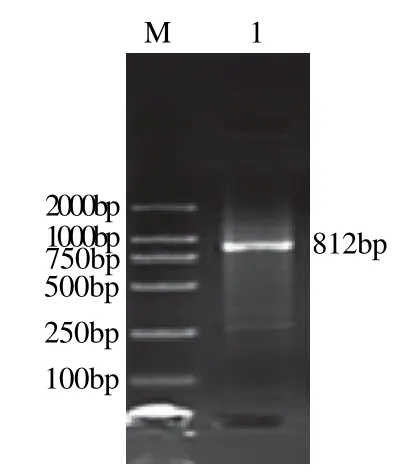
图2 Invitrogen试剂盒UGTC5'-RACE结果Fig.2 Analytical results of 5'-RACE ofUGTCgene by Invitrogen kit
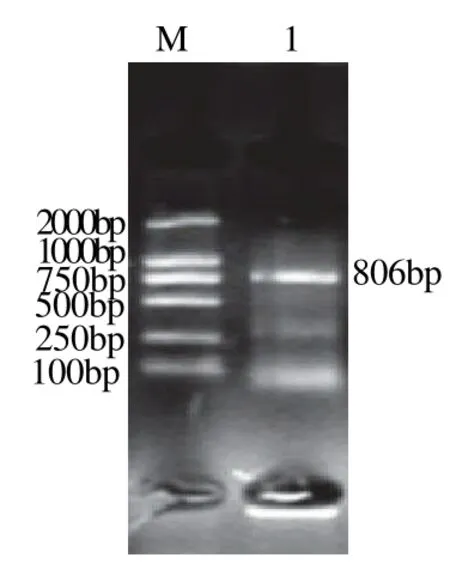
图3 Invitrogen试剂盒UGTR5'-RACE结果Fig.3 Analytical results of 5'-RACE ofUGTRgene by Invitrogen kit

图4 Takara试剂盒UGTC5'-RACE结果Fig.4 Analytical results of 5'-RACE ofUGTCgene by Takara kit
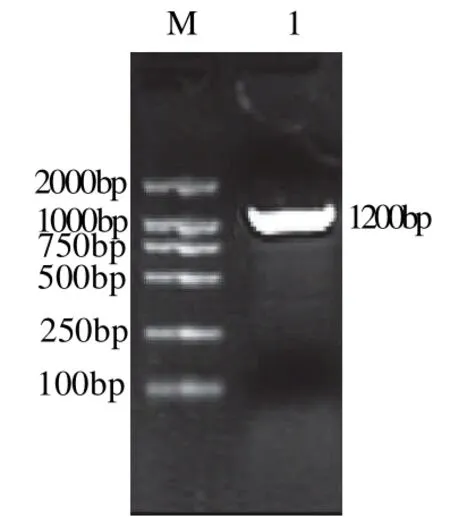
图5 Takara试剂盒UGTR5'-RACE结果Fig.5 Analytical results of 5'-RACE ofUGTRgene by Takara kit
测序结果显示,两个试剂盒扩增的两个酶5′端基因片段都分别都有800个左右碱基序列完全相同,但Takara公司试剂盒扩增得到的基因序列比Invitrogen公司试剂盒扩增产物长200个碱基左右,经过生物信息学分析,Takara公司试剂盒扩增得到的序列包括完整的开放读码框序列,并且分别含有41个和53个碱基的上游非编码序列。说明利用Invitrogen公司试剂盒扩增没有得到基因的全部编码序列,而利用Takara公司试剂盒扩增可以得到包括编码区和非编码区的全部酶基因序列。全序列的序列信息已登陆到GenBank,登录号分别为EF508689和EU567325。
2.3 两个新基因的生物信息学分析
将两个新的基因序列翻译成氨基酸并应用NCBI的BLAST程序进行分析,分析结果如图6、7所示。可以看出两个基因都属于糖基转移酶超家族。UGTC翻译蛋白具有PLN02152型结构域,代表UGTC可能属于UDP-葡萄糖基转移酶类;UGTR翻译蛋白具有PLN02992型结构域,代表UGTR可能属于松柏醇葡萄糖基转移酶类。

图6 UGTC蛋白的结构功能域分析Fig.6 Analysis of structure and function domains in UGTC protein

图7 UGTR蛋白的结构功能域分析Fig.7 Analysis of structure and function domains in UGTR protein
3 讨 论
红景天苷生物合成最后一步反应机制已经清楚,植物体内的尿苷二磷酸(UDP-glucosyltransferase,UDPGT)以尿苷二磷酸葡萄糖(UDPG)和酪醇为底物催化合成红景天苷[17]。因此,提高这个关键酶表达量很有可能提高植物或植物细胞内红景天苷的含量。因此,克隆高山红景天中的糖基转移酶对在分子水平上调控红景天苷的合成具有非常重要的意义。
在线简并引物设计程序CODEHOP与Takara公司的SMARTerTMRACE cDNA Amplification Kit联合使用,对于获得未知基因的cDNA全长序列的效率较高,为新基因分离提供了一条新的快捷途径。
CODEHOP在线设计的简并引物与传统引物设计方法所得引物相比具有扩增特异性高和灵敏度高的优点,原理在于用CODEHOP设计的引物包括两部分:一部分是根据保守氨基酸序列设计的3′简并区,一部分是根据保守氨基酸和密码子偏好性原则预测的最有可能的5′特异夹板区[13,18]。
由于cDNA 5'端的序列各不相同,如何利用已知片断序列得到全长的cDNA,是一个令人困扰的问题。常见的做法是利用末端转移酶在双链cDNA的3'末端加上一连串的G或者C,再通过补齐黏末端,利用已知的两头序列进行扩增(Invitrogen试剂盒原理),但这种方法存在一些问题,如接头的连接效率较低,且这些方法需要多次用不同的酶处理有限的样品,并经过反复的纯化,对于少量样品中的低丰度信息,很大程度上会影响结果的准确性。另外由于mRNA容易部分降解,很难确保得到的cDNA是全长的cDNA。而Takara试剂盒中应用的逆转录酶在以mRNA为模板合成cDNA的时候,当到达mRNA的5'末端碰到真核mRNA特有的“帽子结构”时,即甲基化的G时会连续在合成的cDNA末端合成几个C,由于有5'“帽子结构”的mRNA才能利用这个逆转录反应得到能扩增的cDNA,因此被转录的都是没有降解的RNA,扩增得到的cDNA就都是全长cDNA。
[1] SHELTON A. Variable chemical defences in plants and their effects on herbivore behaviour[J]. Evolutionary Ecology Re-Biol, 2005, 8(3):254-263.
[2] 王莉, 史玲玲, 张艳霞, 等. 植物次生代谢物及其研究进展[J]. 武汉植物学研究, 2007, 25(5):500-508.
[3] 王春丽, 梁宗锁. 外源刺激对植物次生代谢的调节及其信号转导途径研究进展[J]. 西北植物学报, 2009, 29(5):867-873.
[4] 杜丽娜, 张存莉, 朱玮, 等. 植物次生代谢合成途径及生物学意义[J].西北林学院学报, 2005, 20(3):150-155.
[5] JONES P, VOGT T. Glycosyltransferases in secondary plant metabolism:tranquilizers and stimulant controllers[J]. Planta, 2001, 213(2):164-174.
[6] GITZ D C, LAN LIU, MCCLURE J W. Phenolic metabolism, growth,and UV-B tolerance in PAL inhibited red cabbage seedlings[J].Phytochemistry, 1998, 49(2):377-386.
[7] 马莉, 蔡东联, 黎怀星, 等. 红景天苷对小鼠抗疲劳作用的体内研究[J]. 武警医学, 2007, 18(11):818-820.
[8] 张洪志, 王春泉, 孙淑平, 等. 红景天等药物对提高运动员耐力的研究[J]. 航天医学与医学工程, 1996, 9(3):219-220.
[9] DEMENT'EVA L A, IAREMENKO K V. Effect of aRhodiolaextract on the tumor process in an experiment[J]. VoPr Onkol,1987, 33(7):57-60.
[10] 贾正平, 谢景文, 孙卫胜, 等. 狭叶红景天水提物的辐射保护作用[J].兰州医学院学报, 1997, 23(3):17-18.
[11] 张晓丹, 余自云, 张茹. 红景天属植物的化学成分研究进展[J]. 航空航天医药, 2006, 17(1):61-63.
[12] 李建新, 刘巨涛, 金永日, 等. 高山红景天茎叶的化学成分研究[J].中草药, 1998, 29(10):659-661.
[13] 于寒松, 张继星, 马兰青, 等. 利用CODEHOP方法克隆高山红景天葡萄糖基转移酶基因cDNA片段[J]. 吉林农业大学学报, 2008, 30(2):150-153.
[14] BOWLES D, ISAYENKOVA J, LIM E K, et al. Glycosyltransferases:managers of small molecules[J]. Curr Opin Plant Biol, 2005, 8(3):254-263.
[15] LORENC-KUKULA K, KOROBCZAK A, AKSAMIT-STACHURSKA A, et al. Glucosyltransferase:the gene arrangement and enzyme function[J]. Cell Mol Biol Lett, 2004, 9(4B):935-946.
[16] SAMBROOK J, RUSSELL D W. Molecular cloning:a laboratory manual[M]. 3rd ed. NewYork:Cold Spring Harbor Laboratory Press,2001.
[17] BOWLES D, LIM E K, POPPENBERGER B, et al. Glycosyltransferases of lipophilic small molecules[J]. Annu Rev Plant Biol, 2006, 57(6):567-597.
[18] ROSE T, SCHULTZ E, HENIKOFF J, et al. Consensus-degenerate hybrid oligonucleotide primers for amplification of distantly related sequences[J]. Nucleic Acids Res, 1998, 26(7):1628-1635.
Full-length cDNA Cloning of Glycosyltransferase Family fromRhodiola sachalinensis
YU Han-song1,ZHANG Ji-xing2,LI Yan-fang3,MA Lan-qing4,*,HU Yao-hui1,*
(1. College of Food Science and Engineering, Jilin Agricultural University, Changchun 130118, China;2. College of Life Sciences, Inner Mongolia University for the Nationalities, Tongliao 028043, China;3. College of Plant Science, Jilin University, Changchun 130062, China;4. Key Laboratory of Urban Agriculture(North), Ministry of Agriculture, Beijing University of Agriculture, Beijing 102206, China)
Q785
A
1002-6630(2010)21-0244-04
2010-06-30
国家自然科学基金项目(30900112)
于寒松(1979—),男,讲师,博士,研究方向为食品生物技术。E-mail:yuhansong@jluhp.edu.cn
*通信作者:马兰青 (1971—), 男,副教授,博士,研究方向为植物次生代谢分子调控与代谢工程。E-mail:lqma@bac.edu.cn
胡耀辉(1951—),男,教授,学士,研究方向为食品生物制造与新资源利用。E-mail:huyaohui@vip.163.com
