利用分子标记对无核抗病葡萄杂交后代的辅助选择
2010-09-22张剑侠王跃进张艳艳
张剑侠,王 勇,王跃进,张艳艳
(农业部西北园艺植物种质资源利用重点开放实验室,陕西省农业分子生物学实验室,西北农林科技大学园艺学院,陕西 杨凌 712100)
生产上栽培的无核葡萄多为欧亚种(Vitis vinifera L.)品种,品质优良,但抗病性差[1],因此培育抗病无核品种是葡萄育种的重要课题。在无核葡萄常规育种中,是以有核品种作母本、无核品种作父本进行杂交,通常获得无核后代的比率很低[2]。Ramming等发明的无核葡萄胚挽救育种技术,以无核葡萄作母本进行杂交,结合离体胚培养,可使无核杂种比率达到82%[3]。采用该技术,美国已选育出“莫丽莎”等十几个无核优良株系[4]。近20年来,国内在无核葡萄胚挽救育种上也取得了一定的研究进展[5-7],特别是在利用中国野生葡萄的抗病性,进行抗病无核葡萄胚挽救育种方面,获得了较多的胚挽救苗[7],鉴定和筛选抗病无核杂种成为育种程序的下一个重要环节。
传统的育种方法是根据杂种的性状表现和育种者的经验,间接对基因型进行选择,存在花费大、周期长、效率低的问题。分子标记的发展为解决这一问题开辟了新的途径,可以实现对基因型的直接选择[8]。标记辅助选择(Marker-assisted selection,MAS)即是根据与目标性状连锁的分子标记存在与否,对杂种幼苗进行取舍,从而提高育种效率。
王跃进等从无核白葡萄中获得了与无核基因相连锁的RAPD标记UBC269-484,并依据该标记序列,设计合成了检测无核基因的DNA探针GSLP1,能够在无核葡萄中扩增出SCAR标记GS-LP1-569,该SCAR标记与无核主基因的连锁距离为1.2 cM[9-11]。张剑侠等以种间杂交组合白河-35-1(抗病)×佳利酿(感病)亲本及其F1代为试材,获得了中国野生葡萄抗黑痘病基因的RAPD标记OPS03-1354[12],并利用OPS03-1354对该组合的F2代207株及另一组合广西-1(抗病)×京可晶(感病)F1代339株进行了分子检测,结果与田间抗病表现型的符合率分别为83.57%和98.52%[13]。张艳艳等以该组合的亲本及其F1、F2代为试材,获得了6个与中国野生葡萄抗霜霉病基因相连锁的RAPD标记,其中标记S416-1224对F2代109株的检测结果与田间抗病表现型的符合率为78.90%[14]。本研究是在获得无核葡萄胚挽救苗的基础上,利用葡萄无核基因探针GSLP1、抗黑痘病基因RAPD标记OPS03-1354、抗霜霉病基因RAPD标记 S416-1224,对抗病无核葡萄杂交后代进行辅助选择,为进一步选育出无核葡萄新品系奠定基础。
1 材料与方法
1.1 材料
1.1.1 葡萄材料
为西北农林科技大学葡萄种质资源圃中保存的种间杂交、品种自交共6个组合的亲本及其胚挽救后代127株(见表1、2)。
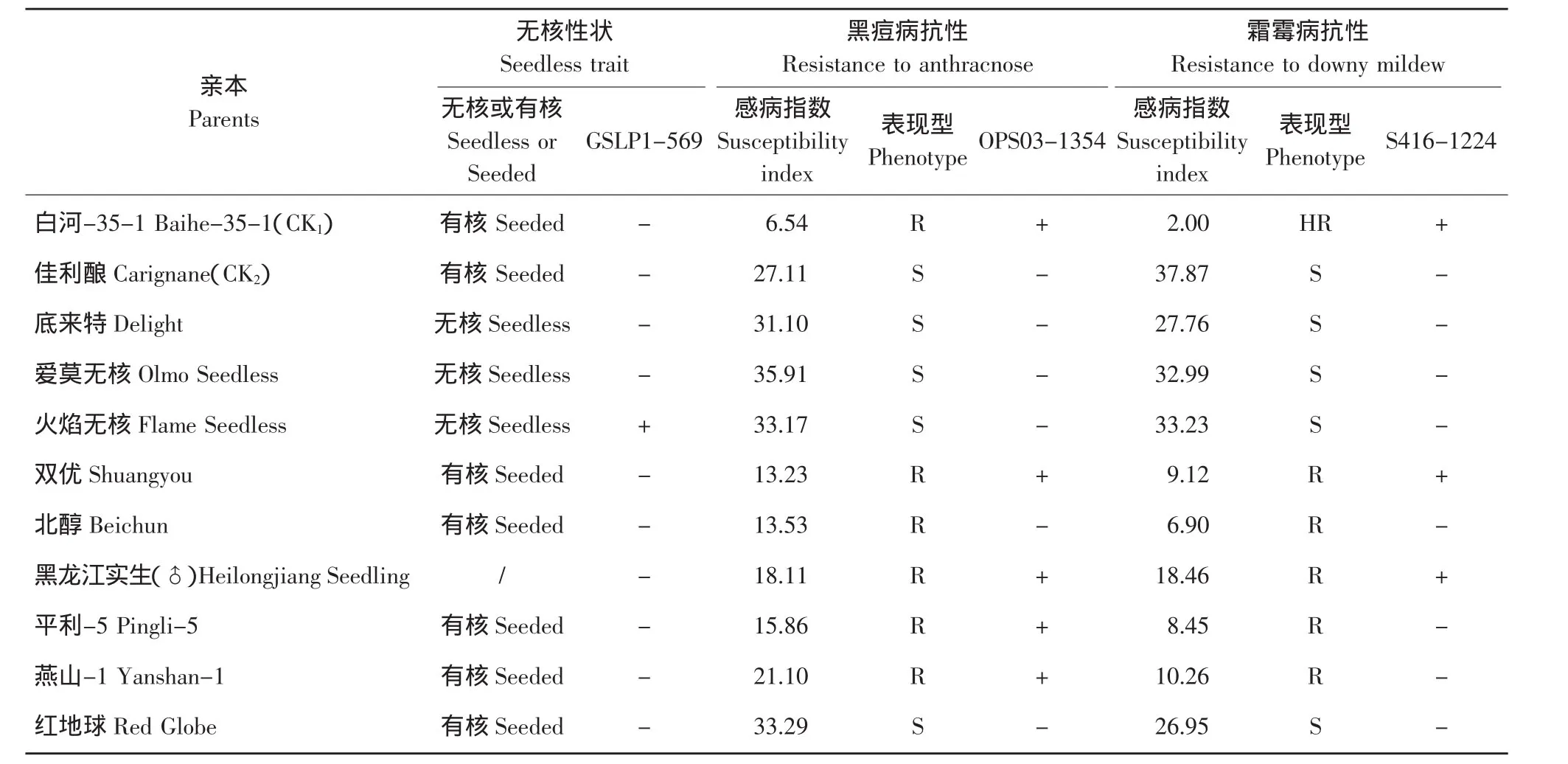
表1 亲本抗黑痘病和霜霉病的表现及其分子标记检测Table1 Resistance to anthracnose and downy mildew of parents and the test results for them by molecular markers
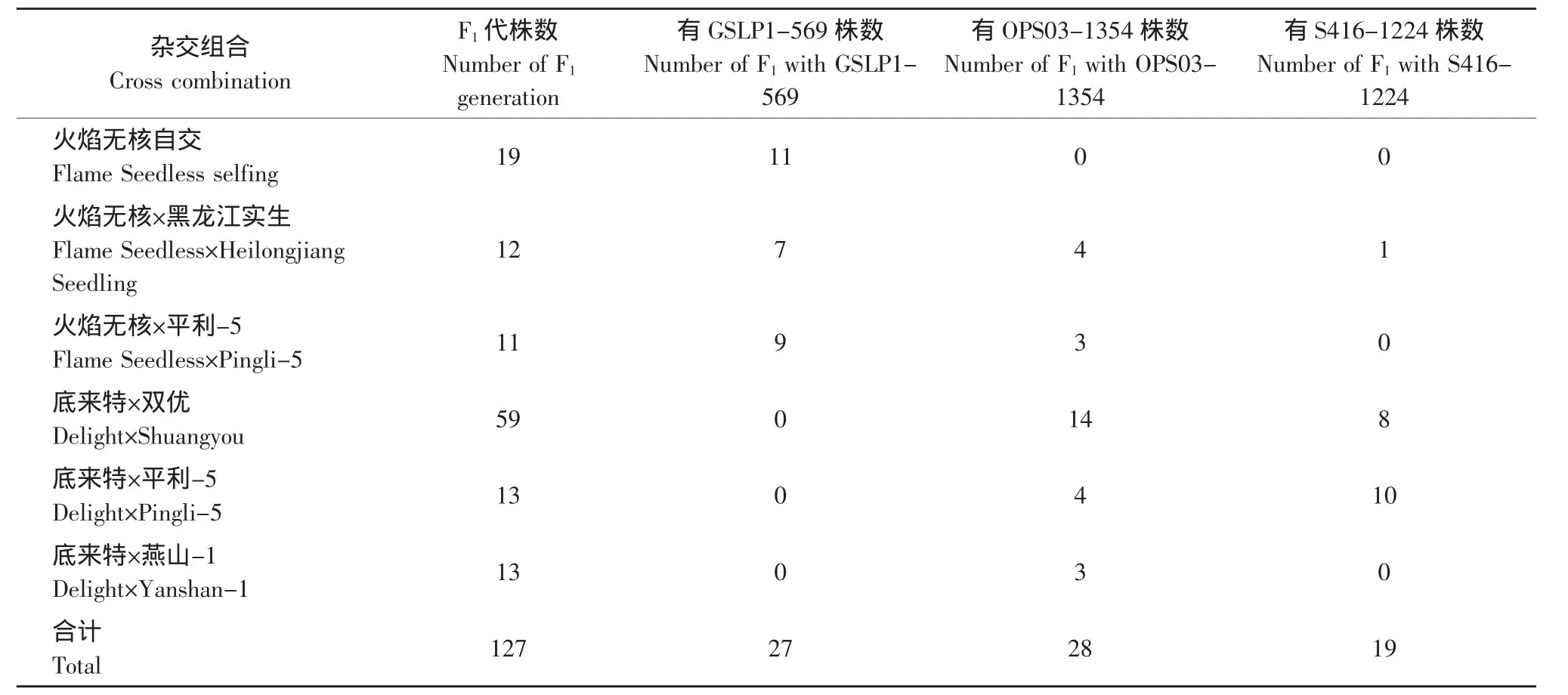
表2 分子标记辅助选择无核葡萄胚挽救杂种Table2 Molecular marker-assisted selection for hybrids from embryo-rescue of grape seedless
1.1.2 分子标记的引物序列
葡萄无核基因SCAR标记GSLP1-569的探针(GSLP1)序列[10-11]:5′CCAGTTCGCCCGTAAATG 3′,中国野生葡萄抗黑痘病基因RAPD标记OPS03-1354 的引物(OPS03)序列[12-13]:5′CAGAGGTCCC 3′;中国野生葡萄抗霜霉病基因RAPD标记S416-1224 的引物(S416)序列[14]:5′GTAACCAGCC 3′。
1.1.3 主要仪器和试剂
PCR仪为PTC-100型,电泳仪为DYY-Ⅲ-8B型,凝胶成像系统为GENE GENIUS型,紫外分光光度计为UV-2802型。Taq DNA聚合酶(华美公司产品);dNTPs(宝鑫生物公司产品);引物(由上海生工公司合成);DNAMarkerDL2000(东胜生物公司产品)。
1.2 方法
1.2.1 葡萄抗黑痘病、霜霉病的鉴定
参照王跃进等对葡萄黑痘病的鉴定方法[15],2008和2009年在田间自然条件下调查杂交亲本叶片的感病百分数(每株随机调查100枚叶片),按0~7级分级,然后换算成感病指数(Susceptibility index,SI)。SI取2年的平均值,按照0~5级分级法,即SI=0,不感病;SI=0.1~0.5,高抗;SI=5.1~25.0,抗病;SI=25.1~50.0,感病;SI=50.1~100,高感。每年的6月份在黑痘病的发病盛期调查黑痘病,8月份在霜霉病的发病盛期调查霜霉病,并以抗两种病害的中国野生华东葡萄株系白河-35-1作为抗病对照(CK1),以感两种病害的欧洲葡萄品种佳利酿作为感病对照(CK2)。
1.2.2 葡萄基因组DNA的提取
参照王跃进等改良的CTAB法[9]。
1.2.3 PCR的反应体系
反应体系为:10×PCR buffer 2.5 μL,MgCl2(15 mmol·L-1)1.5 μL, dNTPs(2.5 mmol·L-1) 2.0 μL,TaqDNA 聚合酶(5 U·μL-1)0.2 μL,引物(20 μmol·L-1)1 μL,模板 DNA 60 ng,以 ddH2O 补足,总体积为25 μL,加盖25 μL矿物油。反应程序为:94℃预变性5 min;94℃1 min,36℃1 min,72℃2 min,共45个循环;然后72℃延伸10 min,终止于4℃。PCR反应重复2次。葡萄无核基因标记检测以无核品种无核白作为对照(CK3)。
1.2.4 PCR产物的电泳分离
采用 1.4%琼脂糖凝胶(含 0.5 μg·mL-1EB)电泳,电泳缓冲液为1×TAE,以DL2000作为分子质量标准,5 V·cm-1电压电泳约1 h,用凝胶成像系统观察、照相。
2 结果与分析
2.1 杂交亲本对黑痘病、霜霉病的抗性表现
由表1可知,在9个杂交亲本中,中国野生葡萄华东葡萄株系白河-35-1(抗病对照CK1)、山葡萄株系双优、山葡萄株系黑龙江实生、秦岭葡萄株系平利-5、燕山葡萄株系燕山-1和欧山杂种北醇均表现为抗黑痘病和霜霉病,而欧洲葡萄品种佳利酿(感病对照CK2)、底来特、爱莫无核、火焰无核和地球红均不抗两种病害。
2.2 分子标记对杂交亲本的检测结果
2.2.1 无核基因标记GSLP1-569对杂交亲本的检测
用获得葡萄无核基因SCAR标记GSLP1-569的探针GSLP1对9个杂交亲本进行PCR检测,结果见图1。除欧洲葡萄无核品种无核白(无核对照CK3)和火焰无核中扩增出了569 bp的DNA标记外,在无核品种底来特、爱莫无核及其他有核品种和株系中均未出现这一标记。这说明无核基因探针GSLP1适用于无核白及与其亲缘关系较近的无核品种(如火焰无核)的检测。
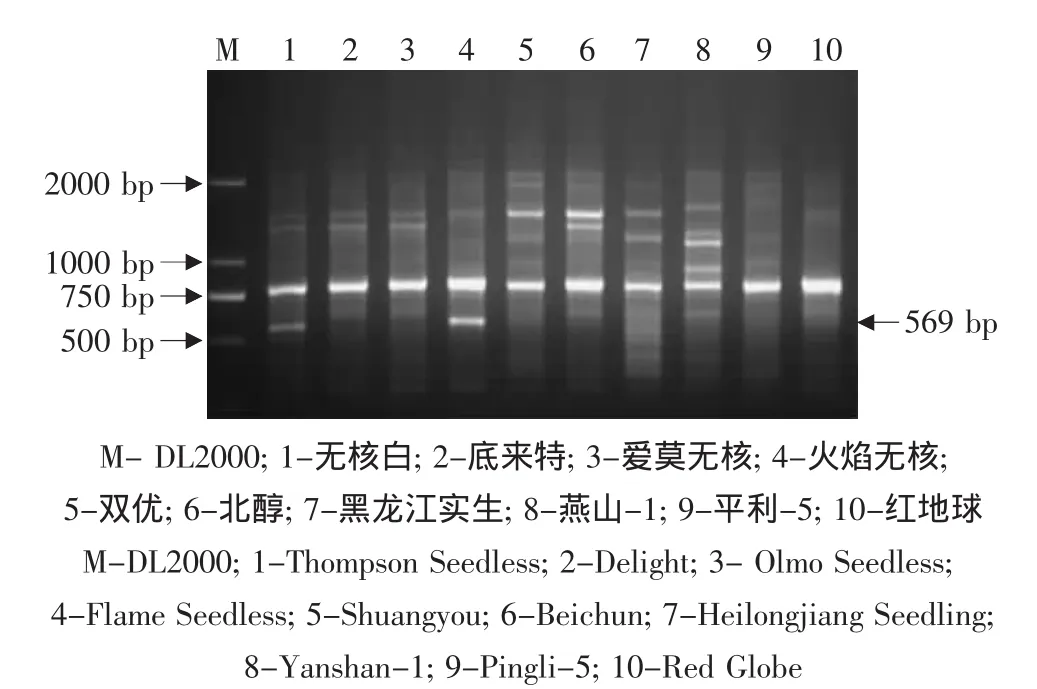
图1 无核基因探针GSLP1在亲本中的PCR扩增Fig.1 PCR amplified result of the parents using the seedless genes probe GSLP1
2.2.2 抗黑痘病基因标记RAPD标记OPS03-1354对杂交亲本的检测
用获得中国野生葡萄抗黑痘病基因RAPD标记OPS03-1354的特异引物OPS03对9个杂交亲本进行RAPD扩增,结果见图2。在中国野生华东葡萄白河-35-1(抗病对照CK1)、山葡萄双优、山葡萄黑龙江实生、燕山葡萄燕山-1、秦岭葡萄平利-5中均存在1 354 bp的RAPD标记,而欧洲葡萄佳利酿(感病对照CK2)、底来特、爱莫无核、火焰无核、地球红及欧山杂种北醇中均不存在。
2.2.3 抗霜霉病基因标记RAPD标记S416-1224对亲本的检测
用获得中国野生葡萄抗霜霉病基因的RAPD标记S416-1224的特异引物S416对9个杂交亲本的检测结果见图3。RAPD标记S416-1224存在于华东葡萄白河-35-1(CK1)、山葡萄双优、山葡萄黑龙江实生和秦岭葡萄平利-5中,而不存在于欧洲葡萄佳利酿(CK2)、底来特、爱莫无核、火焰无核、红地球及欧山杂种北醇、燕山葡萄燕山-1中。

图2 特异引物OPS03对亲本的RAPD扩增Fig.2 RAPD amplified result of the parents using the special primer OPS03

图3 特异引物S416对亲本的RAPD扩增结果Fig.3 RAPD amplified result of the parents using the special primer S416
2.3 分子标记辅助选择杂种后代
2.3.1 无核基因SCAR标记GSLP1-569对杂种后代的检测
由于无核基因SCAR标记GSLP1-569仅存在于亲本火焰无核中,因此对以火焰无核作亲本的杂交后代进行了检测,结果见表2。在火焰无核自交的19株杂种中,有 11株(FZ9、FZ16、FZ19、FZ22、FZ23、FZ29、FZ38、FZ39、FZ40、FZ42、FZ52)存在无核基因标记GSLP1-569(见图4)。在火焰无核×黑龙江实生的12株杂种中,有7株(03-2-7、03-2-14、03-2-16、03-2-18、03-2-19、03-2-20、03-2-24)存在这一标记(见图5)。在火焰无核×平利-5的11株杂种中,有9株(FP1、FP7、FP8、FP9、FP15、FP16、FP17、FP18、FP19)存在这一标记(见图6)。
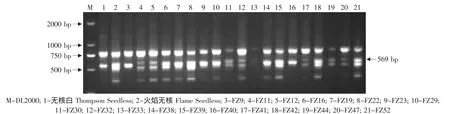
图4 探针GSLP1对火焰无核自交后代的PCR检测Fig.4 PCR amplified result of the progenies from Flame Seedless selfing using the probe GSLP1
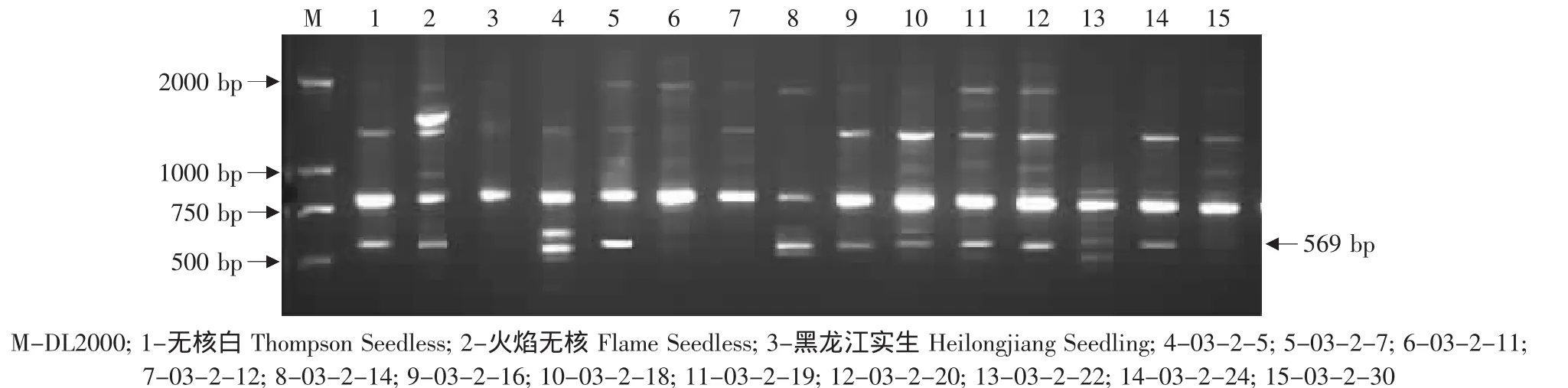
图5 探针GSLP1对火焰无核×黑龙江实生后代的PCR检测Fig.5 PCR amplified result of the hybrids from Flame Seedless×Heilongjiang Seedling using the probe GSLP1
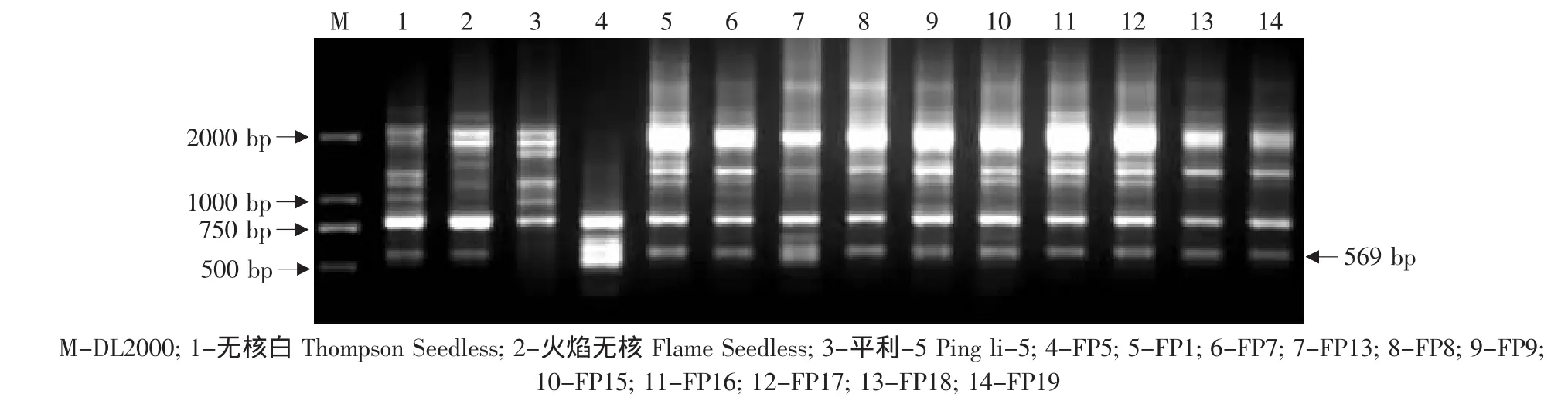
图6 探针GSLP1对火焰无核×平利-5后代的PCR检测Fig.6 PCR amplified result of the hybrids from Flame Seedless×Pingli-5 using the probe GSLP1
2.3.2 抗黑痘病基因RAPD标记OPS03-1354对杂种后代的检测
由于亲本黑龙江实生、平利-5、双优、燕山-1中存在抗黑痘病基因的RAPD标记OPS03-1354,因此对这些野生株系作亲本的杂交组合后代用特异引物OPS03进行RAPD检测,结果见表2。在火焰无核×黑龙江实生的12株杂种中有4株(03-2-14、03-2-19、03-2-24、03-2-30)中存在这一标记(见图7)。在火焰无核×平利-5的11株杂种中有3株(FP5、FP13、FP17)存在这一标记(见图 8)。在底来特×双优的59株杂种中,有14株(03-1-9、03-1-14、03-1-19、03-1-24、03-1-27、03-1-35、03-1-41、03-1-44、03-1-53、03-1-60、03-1-68、03-1-79、03-2-87、03-1-95)存在这一标记(见图9);在底来特×平利-5的13株杂种中有4株(DP4、DP8、DP14、DP17)存在这一标记(见图10);在底来特×燕山-1的13株杂种中有3株(DY3、DY18、DY22)存在这一标记(见图11)。
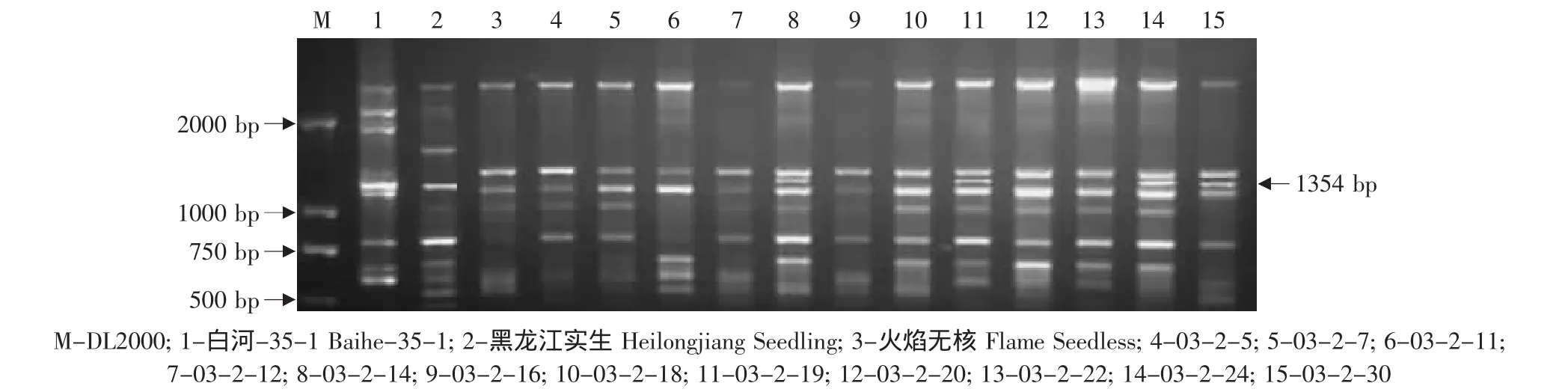
图7 特异引物OPS03对火焰无核×黑龙江实生后代的RAPD扩增Fig.7 RAPD amplified result of the hybrids from Flame Seedless×Heilongjiang Seedling using the special primer OPS03
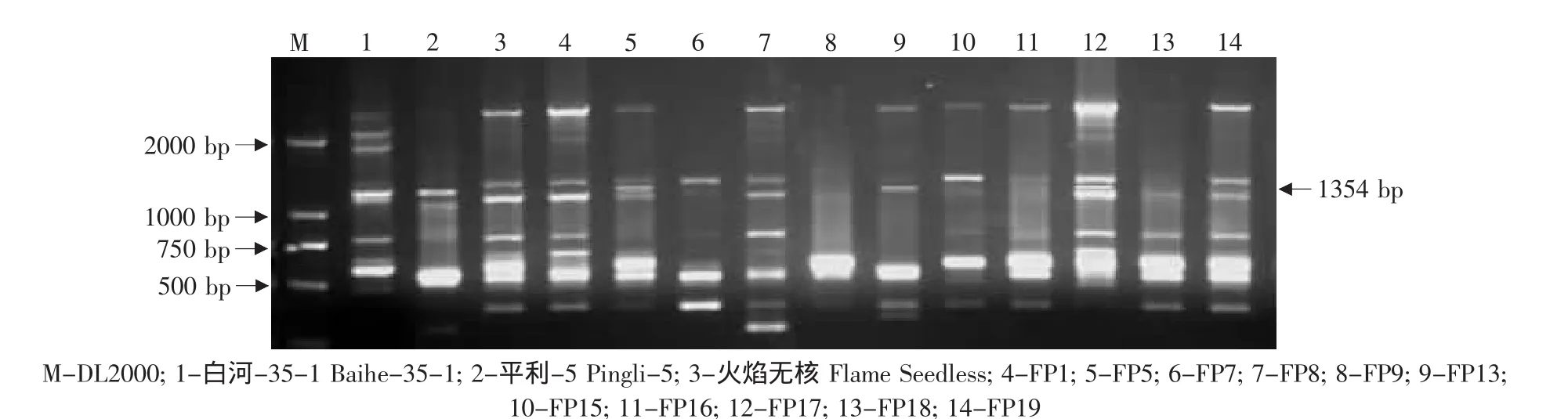
图8 特异引物OPS03对火焰无核×平利-5后代的RAPD扩增Fig.8 RAPD amplified result of the hybrids from Flame Seedless×Pingli-5 using the special primer OPS03
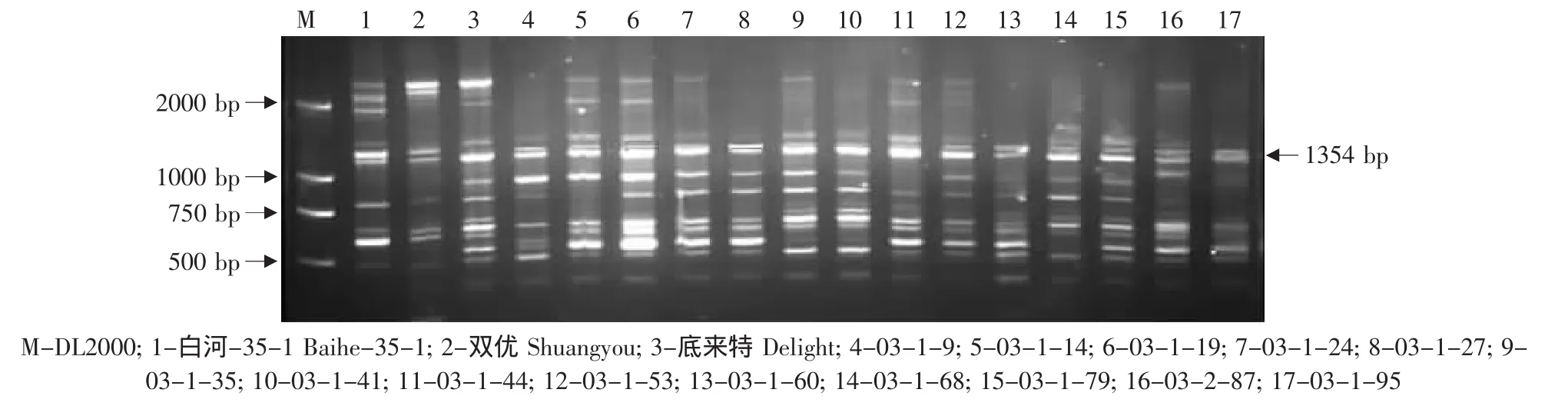
图9 特异引物OPS03对底来特×双优后代的RAPD扩增Fig.9 RAPD amplified result of the hybrids from Delight×Shuangyou using the special primer OPS03
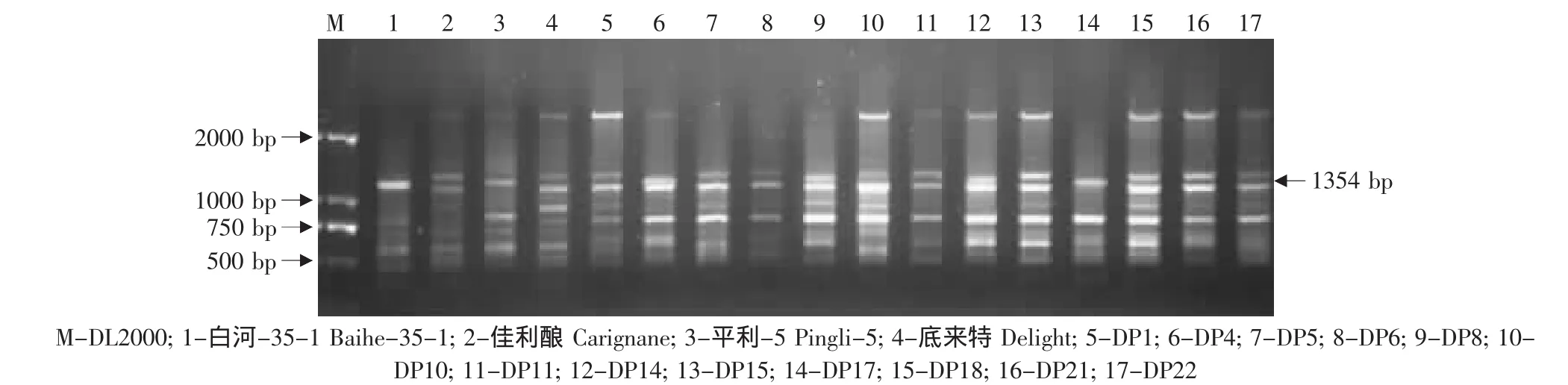
图10 特异引物OPS03对底来特×平利-5后代的RAPD扩增Fig.10 RAPD amplified result of the hybrids from Delight×Pingli-5 using the special primer OPS03
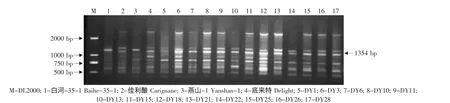
图11 特异引物OPS03对底来特×燕山-1后代的RAPD扩增Fig.11 RAPD amplified result of the hybrids from Delight×Yanshan-1 using the special primer OPS03
2.3.3 抗霜霉病基因RAPD标记S416-1224对杂种后代的检测
由于亲本黑龙江实生、平利-5、双优中存在抗霜霉病基因的RAPD标记S416-1224,因此对这些野生株系作亲本的杂交组合后代用特异引物S416进行RAPD检测,结果见表2。在火焰无核×黑龙江实生的12株杂种中,有1株(03-2-12)中存在这一标记(见图12);在底来特×平利-5的13株杂种中,有10 株(DP1、DP4、DP5、DP6、DP8、DP15、DP17、DP18、DP21、DP22)中存在这一标记(见图 13);在底来特×双优的59株杂种中,有8株(03-1-5、03-1-14、03-1-19、03-1-24、03-1-27、03-1-44、03-1-79、03-1-87)存在这一标记(见图14);在火焰无核×平利-5的11株杂种中不存在这一标记。
综上所述,对6个杂交组合127株胚挽救苗的分子检测结果表明,拥有无核基因SCAR标记GSLP1-569的杂种有27株,其中4株杂种03-2-14、03-2-19、03-2-24(火焰无核×黑龙江实生)和FP17(火焰无核×平利-5)还同时拥有抗黑痘病基因RAPD标记OPS03-1354。拥有抗黑痘病基因RAPD标记OPS03-1354的杂种28株,拥有抗霜霉病基因RAPD标记S416-1224的杂种19株,其中有10株杂种DP1、DP8、DP17(底来特×平利-5)、03-1-14、03-1-19、03-1-24、03-1-27、03-1-44、03-1-79、03-1-87(底来特×双优)同时拥有抗黑痘病基因和抗霜霉病基因RAPD标记。

图12 特异引物S416对火焰无核×黑龙江实生后代的RAPD扩增Fig.12 RAPD amplified result of the hybrids from Flame Seedless×Heilongjiang Seedling using the special primer S416
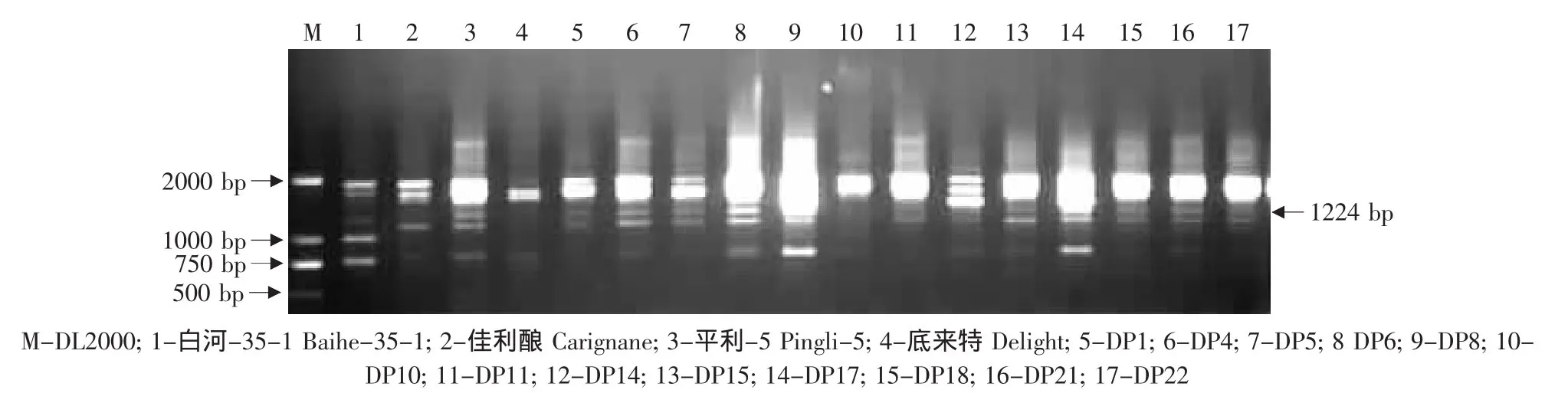
图13 特异引物S416对底来特×平利-5后代的RAPD扩增Fig.13 RAPD amplified result of the hybrids from Delight×Pingli-5 using the special primer S416
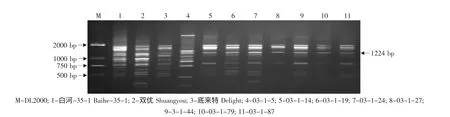
图14 特异引物S416对底来特×双优后代的RAPD扩增结果Fig.14 RAPD amplified result of the hybrids from Delight×Shuangyou using the special primer S416
3 讨论
葡萄黑痘病(Sphacerlouna ampelinum)和霜霉病(Plasmopara viticola)是葡萄上的两种重要真菌病害,发病条件有较大的相似性,均是高湿和适宜的温度下(18~25℃)容易发生和流行,特别是能对欧洲葡萄造成严重的危害[15-18]。因此,除了无核性状,抗病性也是葡萄育种的重要目标之一,培育抗病无核葡萄品种是生产发展的迫切需要。中国野生葡萄资源丰富,其中的一些种和株系抗病性极强[15-17],利用中国野生葡萄的抗病性与欧洲葡萄无核品种杂交是培育抗病无核葡萄品种的一个有效策略。而利用与无核性状相连锁的基因标记和与抗病性状相连锁的基因标记对杂种后代进行辅助选择,将会节省人力、物力和财力,提高育种效率,缩短育种周期。Striem等应用RAPD标记对早玫瑰(Early Muscat)×火焰无核(Flame Seedless)F1代的无核亚性状进行早期MAS,可以淘汰约44%的有核杂种[19]。Scott等利用AFLP技术对火焰无核的突变体进行了早期鉴定[20]。王跃进等利用葡萄无核基因DNA探针对我国栽培的无核葡萄品种进行了检测,并初步应用于辅助育种[21]。Kim等获得了葡萄抗黑痘病基因的RAPD标记OPB15-1247,进一步发展成为SCAR标记,应用于黑痘病抗性基因型的基因位点检测[22]。Riaz等利用来自峡谷葡萄b43-17抗皮尔斯病基因的微卫星标记VVIP26、ctg1026876和VMC2a5对创建的F1代、回交一代、回交二代共4 321株幼苗进行辅助选择,筛选出抗皮尔斯病的幼苗1 683株[23]。尽管分子标记研究在葡萄上取得了一定的进展,但MAS在育种上的应用仍属于起步阶段。
本研究利用葡萄无核基因SCAR标记GSLP1-569、抗黑痘病基因RAPD标记OPS03-1354和抗霜霉病基因RAPD标记S416-1224,对6个杂交或自交获得的127株胚挽救苗进行了分子检测,从中筛选出了无核杂种、抗黑痘病的无核杂种、抗黑痘病的有核杂种、抗霜霉病的有核杂种、抗黑痘病且抗霜霉病的有核杂种。对于筛选获得的无核杂种单株,下一步将重点观察其田间对抗黑痘病和霜霉病抗性以及结果后的无核性状,从中选出无核葡萄新品系。而仅拥有抗病基因标记的杂种,对其果实性状如大粒、色泽艳丽、脆肉、可溶性固形物含量高等表现突出的单株可作为进一步杂交育种的亲本予以利用。
在本研究中,无核基因探针GSPL1在无核品种无核白(无核对照CK3)和火焰无核上能够扩增出无核基因SCAR标记GSLP1-569,而且能够在火焰无核自交后代中检测出该标记,说明无核基因探针GSLP1对与无核葡萄原始亲本无核白亲缘关系较近的无核品种或其杂种具有良好的检测作用。该无核标记未能在无核品种底来特、爱莫无核中检测出来,从分子角度推测葡萄无核基因可能是多个,属于数量性状,对于以底来特、爱莫无核等无核品种作母本杂交获得的胚挽救苗的标记辅助选择,还有待于进一步研究和获得新的无核基因标记及其探针。同样,抗黑痘病基因RAPD标记OPS03-1354和抗霜霉病基因RAPD标记S416-1224存在于中国野生葡萄中,而不存在于欧洲葡萄品种中,对欧洲葡萄无核品种×欧洲葡萄品种的杂交组合的亲本及其后代不能够进行检测,因此在具体的标记辅助选择应用中,RAPD标记OPS03-1354和S416-1224仅适合于亲本之一为中国野生葡萄的杂交后代的分子检测。同时,将这两个RAPD标记转换成稳定性、专一性更好的SCAR标记,将会进一步提高标记辅助选择的准确性。对于不存在显性或主效抗病基因的欧洲葡萄,研究和开发其他类型的抗病基因分子标记将有助于对该种内杂交后代的辅助选择,这也将是我们今后研究的重点内容之一。
3 结论
a.葡萄无核基因探针GSLP1对与无核葡萄原始亲本无核白亲缘关系较近的无核品种及其杂种具有良好的检测无核基因和无核性状的作用。
b.中国野生葡萄抗黑痘病基因RAPD标记OPS03-1354和抗霜霉病基因RAPD标记S416-1224适合于中国野生葡萄及其杂交后代的抗性检测。
c.利用葡萄无核基因SCAR标记GSLP1-569及中国野生葡萄抗黑痘病基因RAPD标记OPS03-1354和抗霜霉病基因RAPD标记S416-1224,对6个组合的127株无核葡萄胚挽救苗进行无核、抗病的分子检测,筛选出拥有无核基因SCAR标记GSLP1-569的杂种有27株,其中4株同时拥有抗黑痘病基因RAPD标记OPS03-1354。拥有抗黑痘病基因RAPD标记OPS03-1354的杂种28株,拥有抗霜霉病基因RAPD标记S416-1224的杂种19株,其中有10株同时拥有抗黑痘病基因和抗霜霉病基因RAPD标记。获得的无核、抗病葡萄杂种将为进一步选育出新品系奠定基础。
[1]Dalbó M A,Ye G N,Weenden N F,et al.Marker-assisted selection for powdery mildew resistance in grapes[J].Journal of the American Society for Horticultural Science,2001,126(1):83-89.
[2]Ledbetter C A,Burgos L.Inheritance of stenospermocarpic seedle-ssness in Vitis vinifera L.[J].The Journal of Heredity,1994,85(2):157-160.
[3]Ramming D W,Emershad R L.In ovule embryo culture of seeded and seedless Vitisvinifera(Abstr)[J].HortScience,1982,17(11):487.
[4]王跃进,万怡震.美国加州的葡萄生产和科研[J].西北农林科技大学学报:自然科学版,2002,3(1):134-140.
[5]唐冬梅,蔡军社,骆强伟,等.用于无核葡萄选育的胚挽救技术研究[J].果树学报,2008,25(3):316-321.
[6]郭修武,郭印山,张海娥,等.接种时期和培养基对无核葡萄胚挽救的影响[J].园艺学报,2007,34(2):329-332.
[7]Tian L L,Wang Y J,Niu L,et al.Breeding of disease-resistant seedless grapes using Chinese wild Vitis spp.in vitro embryo rescue and plant developement[J].Scientia Horticulturae,2008,117:136-141.
[8]方宣钧,吴为人,唐纪良.作物DNA标记辅助育种[M].北京:科学出版社,2002.
[9]王跃进,Lamikanra O.葡萄无核基因的RAPD遗传标记[J].西北农业大学学报,1996,24(5):1-10.
[10]王跃进,Lamikanra O.检测葡萄无核基因DNA探针的合成与应用[J].西北农林科技大学学报:自然科学版,2002,30(3):42-46.
[11]杨克强,王跃进,张今今,等.葡萄无核基因定位与作图的研究[J].遗传学报,2005,32(3):297-302.
[12]张剑侠,王跃进,周鹏,等.中国野生葡萄抗黑痘病基因的RAPD标记[J].果树学报,2001,18(2):68-71.
[13]张剑侠,王跃进,张艳艳,等.中国野生葡萄抗黑痘病基因RAPD标记的克隆、序列分析及辅助育种应用[J].果树学报,2009,26(4):306-310.
[14]张艳艳,张剑侠,王跃进.中国野生葡萄抗霜霉病基因RAPD标记的筛选[J].果树学报,2008,25(6):816-820.
[15]王跃进,贺普超.葡萄白腐病和黑痘病抗性鉴定方法[J].西北农林科技大学学报:自然科学版,1988,16(3):17-32.
[16]贺普超,王跃进,王国英,等.中国葡萄属野生种抗病性研究[J].中国农业科学,1991,24(3):50-56.
[17]Wang Y,Liu Y,He P.Resistance of Chinese Vitis species to Elsinoe ampelina(de Bary)Shear[J].Hort Science,1998,33(1):123.
[18]刘会宁,李华.欧亚种葡萄对白粉病与霜霉病的抗性研究[J].东北农业大学学报,2004,35(3):302-308.
[19]Striem M J,Ben-Hayyim G,Spiegel-Roy P.Identifying molecular genetic markers associated with seedlessness in grape[J].Journal of the American Society for Horticultural Science,1996,121(5):758-763.
[20]Scott K D,Ablett E M,Lee L S,et al.AFLP markers distinguishing an early mutant of Flame Seedless grape[J].Eupytica,2000,113:45-249.
[21]王跃进,杨英军,周鹏,等.用DNA探针检测我国栽培的无核葡萄及辅助育种初探[J].园艺学报,2002,29(2):105-108.
[22]Kim G H,Yun H K,Choi C S.Identification of AFLP and RAPD markers linked to anthracnose resistance in grapes and their conversion to SCAR markers[J].Plant Breeding,2008,127(4):418-423.
[23]Riaz S,Tenscher A C,Graziani R,et al.Using marker-assisted selection to breed pierce′s disease-resistant grapes[J].American Journal of Enology and Viticulture,2009,60(2):199-207.
