嫩江沙地4种针叶树光合特性及生态适应性特征
2010-08-08王力刚许成启
王力刚 赵 岭 许成启 李 峰 高 野
(黑龙江省森林与环境科学研究院,齐齐哈尔,161005)
嫩江沙地地处黑龙江省西部半干旱区腹地,该区原生植被以耐旱的灌木类为主体,在河流沿岸有野生的杨柳类乔木分布,针叶树种无天然分布。目前该区较早引入的“三松”(长白落叶松、红皮云杉、樟子松)经近半个世纪的适应,基本上已经“安家落户”,目前在生产上已大量应用,其间又相继引入了一些其他针叶树品种,但表现不如“三松”,其中长白松相对适应性强一些。近年来,随着气候变化造成的极端气候年份增多,对这些外来树种形成挑战,对于嫩江沙地而言,气候变化的显著影响使旱情加重,由此对引入树种的适应性构成影响,因此重新对这些树种进行评价很有必要。众所周知,树种的光合特性及水分利用效率是其基本的生理生态特性,这些特性往往是其节水抗旱性能的生理表现,其差异在一定程度上反映树种之间节水、抗旱性能的不同。为此,本研究对嫩江沙地4种针叶树(长白落叶松、红皮云杉、樟子松、长白松)的主要光合参数及水分利用效率进行比较分析,探讨其耗水、抗旱性能的差异,进而为该区抗旱节水树种筛选及适应性评价提供重要依据。
1 材料与方法
嫩江沙地概况:嫩江沙地地处黑龙江省西南部半干旱区的嫩江下游,地理坐标为东经122°10′~125°10′,北纬45°30′~48°30′,沿嫩江干流两侧宽165 km、长400 km的范围内,形成东北—西南走向的带状沙区,包括杜蒙、泰来、龙江、齐市郊区、富裕、甘南、肇源、大庆及讷河部分或全部,面积274.7万hm2,其中沙漠化土地37.8万hm2。该区属于中温带大陆性季风气候,春季风大,蒸发量大;夏季高温多雨,秋季早霜低温,冬季漫长寒冷。≥10℃活动积温2 550~2 920℃,降水量376~465 mm。沙区地表水比较缺乏,而地下水较丰富。地带性土壤为黑钙土,同时新生的风沙土较多,另有草甸土、盐碱土和沼泽土零星分布。植被类型以蒙古植物区系为主,属于草甸草原植被。在沙区上有零星或丛状分布的天然木本植物——榆树、桑树、山杏、山竹子、鼠李及半灌木兴安胡枝子;沿江分布着由小黄柳、蒙古柳等构成的灌丛。人工木本植物以各类人工培育的杨树为主体,其次为人工引进的一些外来品种,如柳树、锦鸡儿、丁香、胡枝子、沙棘及一些针叶树(樟子松、落叶松、云杉等)。
材料选取与处理:选取红皮云杉(Picea koraiensis Nakai)、樟子松(Pinus sylvestris L.var.mongolica Litv.)、长白松(Pinus sylvestris var.sylvestriformis)、长白落叶松(Larix olgensis Henry)的3年生容器苗(在容器袋内培育3 a)为试验材料。将选定的试验材料于2007年4月中旬,在黑龙江省森林与环境科学研究院的绿源科技试验示范基地内布设盆栽试验,每个品种选取8株健康苗木置于试验瓦盆内,盆内径55 cm,高50 cm。所有试验苗木置于室外同一环境条件下进行浇水、松土、拔草等管理,待到8月份苗木已经正常生长时开始测定工作。本次试验测定时间选在8月中旬,此时各树种已进入生长旺季。
采用美国拉哥公司(LI-COR)生产的Li-6400便携式光合作用测定系统,对选定的试验材料的叶片光合速率(Pn)、蒸腾速率(Tr)及气孔导度(Cond)等指标进行测定。在每一树种的8株树中选择标准株(就是生长发育正常良好的作为待测标准株),在标准株上选取中上部生长健康的新生叶进行活体测定。每个标准株上选3束标准功能叶,每束叶每次测定重复3次,3束的平均值作为其相应指标的测定值。在测定时为使所测数据具有直接可比性,每次测定都将叶室放满针叶。
Li-6400光合作用测定系统具有可调和的稳定光照,为了分析各树种不同光照梯度下光合参数的变化规律,本试验设置了 0、50、200、500、1 000、1 800 μmol·m-2·s-16 个光照梯度。
数据处理通过EXCEL完成,计算了各树种叶片在测定光照强度范围内单位面积平均光合速率、蒸腾速率、气孔导度、水分利用效率等。
2 结果与分析
2.1 各树种光合速率的差异
一般情况下,在植物生育期的生长旺季,在一定光强范围内其光合速率随光强的增大而增大[1],本试验同样证明了这一点。随着光强由 0 增至 1 800 μmol·m-2·s-1,各试验树种光合速率呈逐渐增大态势,但各树种之间呈现明显差异(图1),平均光合速率位次为红皮云杉>长白松>樟子松>长白落叶松,其值分别为 5.80、5.08、4.16、1.64 μmol·m-2·s-1,前3个树种差异较小,与最小的长白落叶松差异较大。在光强为 1 000 μmol·m-2·s-1时,红皮云杉光合速率达到峰值,为10.5 μmol·m-2·s-1,然后开始缓慢下降,至 1 800 μmol·m-2·s-1光强时降至 10.3 μmol·m-2·s-1;樟子松、长白松则一直持续增大,到 1 800 μmol·m-2·s-1光强时光合速率已经超过红皮云杉,都达到 11.5 μmol·m-2·s-1,且樟子松的增幅最大。长白落叶松光合速率增幅一直比较平缓,随光强增大与前3位的差距逐渐加大。
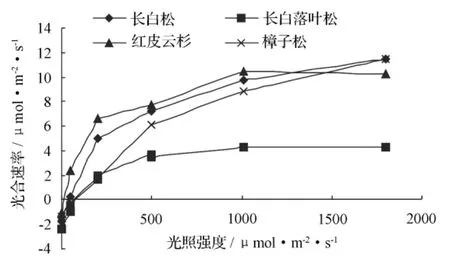
图1 各树种光合速率随光强的变化
2.2 各树种蒸腾速率的差异
蒸腾速率作为植物水分生理的一个重要指标,是植株对水分生理需求的一个量度,也是其节水、抗旱性能的反映,较高的蒸腾速率反映了较高的生理耗水需求[2-3]。
图 2 表明,随着光强在 0 ~ 1 800 μmol·m-2·s-1范围内逐渐递增,各树种蒸腾速率开始增加较快,到一定光强后增加缓慢,并逐渐达到一个峰值,然后缓慢减小,但各树种之间同样显示了不同程度的差异。
相对来讲,针叶树普遍比阔叶乔木抗旱能力强,较为节水,但它们之间也存在一定差异。本试验结果表明:4种针叶树蒸腾速率位次为樟子松>长白松>红皮云杉>长白落叶松。图 2 显示:长白松与樟子松在 0 ~1 800 μmol·m-2·s-1光强范围内,蒸腾速率曲线比较接近,其平均值也较为接近,分别为 2.47、2.50 mmol·m-2·s-1;红皮云杉的平均值为 2.20,与之有一定差距;落叶松最小,为 1.50 mmol·m-2·s-1,与前三者差距较大。4种针叶树蒸腾速率比较结果表明:长白落叶松相对较为节水,其次为红皮云杉,而长白松与樟子松耗水则相对较大。
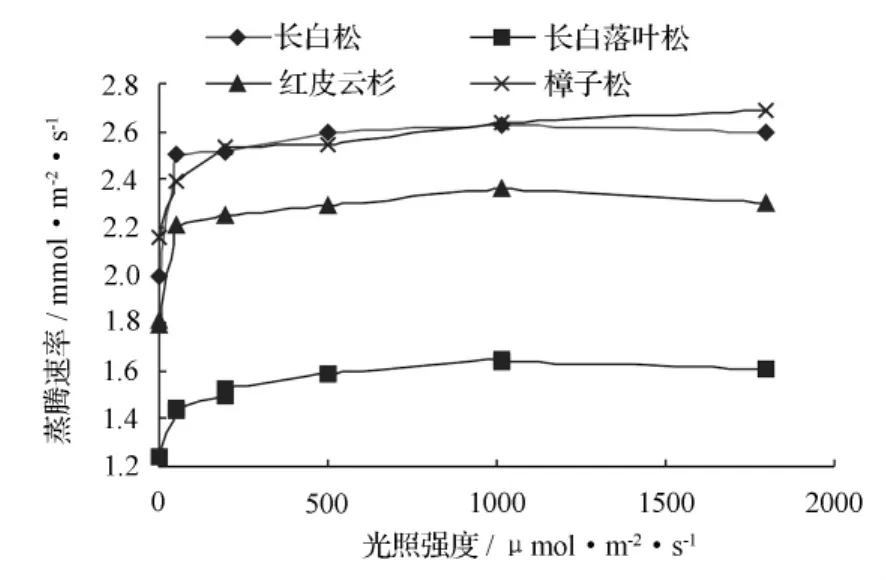
图2 各树种蒸腾速率随光强的变化
2.3 各树种气孔导度的差异
气孔导度(用气孔阻力的倒数表示)是气孔状况的指标,其大小表明CO2、水蒸气出入细胞的难易程度,进而影响到叶片的光合强度和蒸腾强度[4]。
图3表明:各树种气孔导度随光强增强而增大,到某一光强时,气孔导度达到一个峰值,之后随光强继续增强,气孔导度开始逐渐降低,在 0 ~1 800 μmol·m-2·s-1光强范围内,各树种达到峰值的光强不同。气孔导度变化曲线也随树种不同而不同:气孔导度位次为樟子松>长白松>红皮云杉>长白落叶松,平均值分别为 0.17、0.16、0.13、0.099 μmol·m-2·s-1,表明参试针叶树相对而言,长白落叶松呈节水表现,其次为红皮云杉,而樟子松与长白松则耗水较多。
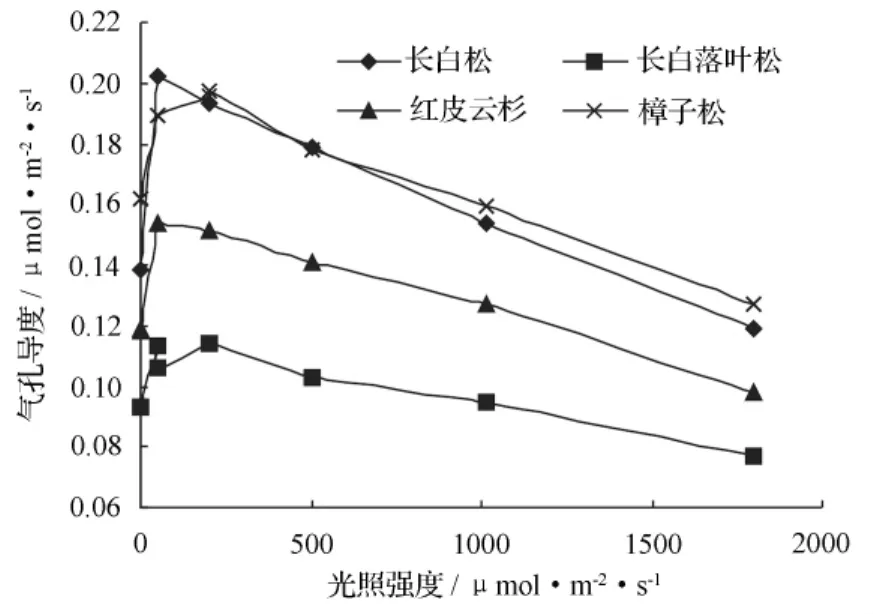
图3 各树种气孔导度随光强梯度的变化
2.4 各树种水分利用效率的差异
树木叶片水分利用效率是指单位水量通过叶片蒸腾散失时光合作用所同化的CO2的量,为光合速率与蒸腾速率的比值(Pn/Tr)[5],它能较为准确地反映出树木叶片的瞬间或短期的反应行为,是评价水分亏缺条件下树木生长适宜程度的一个综合生理生态指标,可在一定程度上反映出树木的耗水性和抗旱性[6-7]。供试各树种水分利用效率随光强梯度的变化情况见图4。
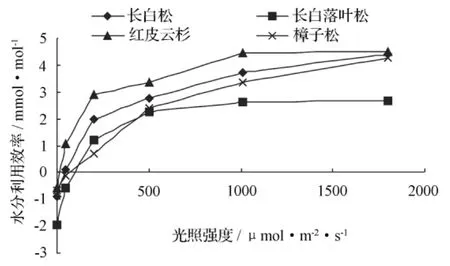
图4 各树种水分利用效率随光强梯度的变化
图4显示,各树种水分利用效率随光强的递增呈逐渐增长态势,与光强呈二次曲线(抛物线)关系,各树种水分利用效率在 0 ~1 800 μmol·m-2·s-1光强内与光强的关系较为规律,但各树种之间存在一定差异:红皮云杉水分利用效率最高,平均值为 2.61 mmol·mol-1,其次为长白松(2.03),最小为长白落叶松(1.03),樟子松为 1.655 mmol·mol-1,仅高于长白落叶松。红皮云杉在较弱光强下就表现了较高的水分利用效率,在光强为 50 μmol·m-2·s-1时,其水分利用效率就已超过1 mmol·mol-1(而此时长白落叶松和樟子松还处于负值状态),且随光强梯度递增,一直保持较高的水分利用效率,光强超过 1 000 μmol·m-2·s-1后,红皮云杉水分利用效率开始变得平缓,增长变缓,此时长白松和樟子松增长幅度较红皮云杉大,表现出在强光下樟子松和长白松水分利用效率增长加快,是其喜光特性的反映。落叶松在光强为500 μmol·m-2·s-1之前,水分利用效率增长较快,之后则变得平缓,水分利用效率增长变缓,且一直处于较低的位次。
3 结论与讨论
供试的4个针叶树品种中,长白落叶松、红皮云杉、樟子松在嫩江沙地引入较早且已经成为该区的主要造林树种;而长白松自20世纪80年代中期引入,近年来又重新引入,面积不大,属于新引入针叶品种,从目前长势及适应性看,前期生长较樟子松快[8]。从4个品种相对来看,红皮云杉属光合速率较高、耗水较低、水分利用效率较高的品种,长白落叶松属光合、耗水、水分利用效率都较低的品种,二者都属于相对节水型树种,尤其是长白落叶松蒸腾速率明显低于其它3个树种;而长白松与樟子松相对蒸腾速率较高,属耗水量较大树种。从生产上表现,樟子松属于抗旱性较强树种[9],但其蒸腾却较大,可见节水与抗旱性虽关系密切,却有着不同的适应机制,筛选既节水又抗旱的品种任务仍很艰巨。
树种的水分生理与光合生理是比较复杂的生物物理化学过程,除与自身结构、适应机制有关外,还与外界许多环境因子,如大气温度、湿度、土壤水分有着较为密切的联系[1],而且是许多因子综合作用的结果。光照强度是影响其光合特性十分重要的因子之一,随着其梯度变化,相应的光合参数发生变化,各树种之间的变化比较则体现其某些生理特性的差别,这些差异可作为筛选节水、抗旱植物的依据。对各树种之间日进程的比较及在不同土壤水分梯度下的比较研究还须深入,从而为节水抗旱性树种的筛选提供更为充分的生理学依据。
[1] 常宗强,冯起,苏定红,等.额济纳洲胡杨的光合特性及其对光强和 CO2浓度的响应[J].干旱区地理,2006,29(4):496-502.
[2] 贺康宁,田阳,史常青,等.黄土半干旱区集水造林条件下林木生长适宜的土壤水分环境[J].林业科学,2003,39(1):10-16.
[3] 赵长明,魏小平,尉秋实,等.民勤绿洲荒漠过渡带植物白刺和梭梭光合特性[J].生态学报,2005,25(8):1909-1913.
[4] 徐俊增,彭世彰,丁加丽,等.水利学报[J],2006,37(4):486-491.
[5] 刘昌明,王会肖.土壤-作物-大气界面水分过程与节水调控[M].北京:科学出版社,1999:30-37.
[6] 王颖,魏国印,张志强,等.7种园林树种光合参数及水分利用效率的研究[J].河北农业大学学报,2006,29(6):44-48.
[7] 李国泰.8种园林树种光合作用特征与水分利用效率比较[J].林业科学研究,2002,15(3):291-296.
[8] 杨金龙,范宗泉,王立刚,等,长白松引种育苗及造林技术[J].防护林科技,2007(3):130-131.
[9] 杨克杰.樟子松遗传改良及栽培[M].赤峰:内蒙古科学技术出版社,2002:3-5.
